
Identification of Novel Molecular Targets of Four Microcystin Variants by High-Throughput Virtual Screening

 , ,
, ,  and
and 
Abstract
1. Introduction
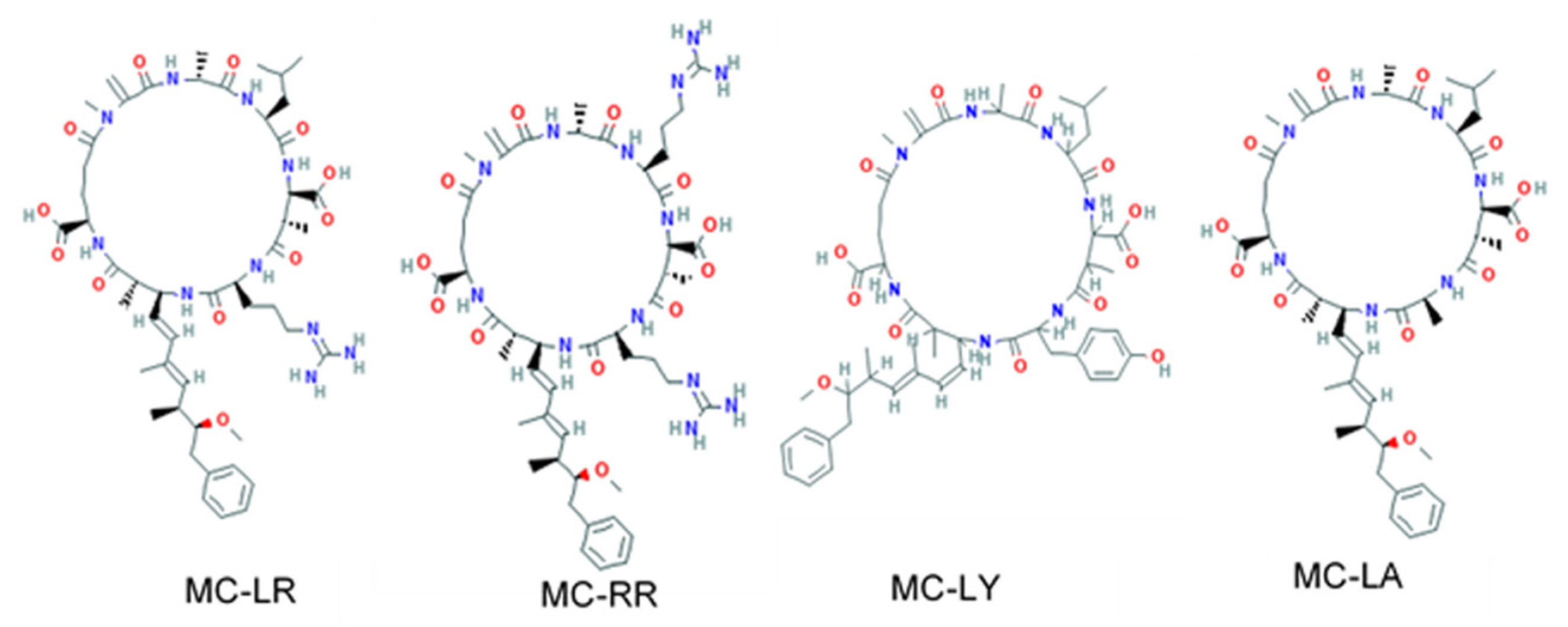
2. Materials and Methods
Protein–Ligand Coupling Calculations with AutoDock Vina
3. Results and Discussion
3.1. Main Complexes Formed with Microcystins with High Affinity Scores
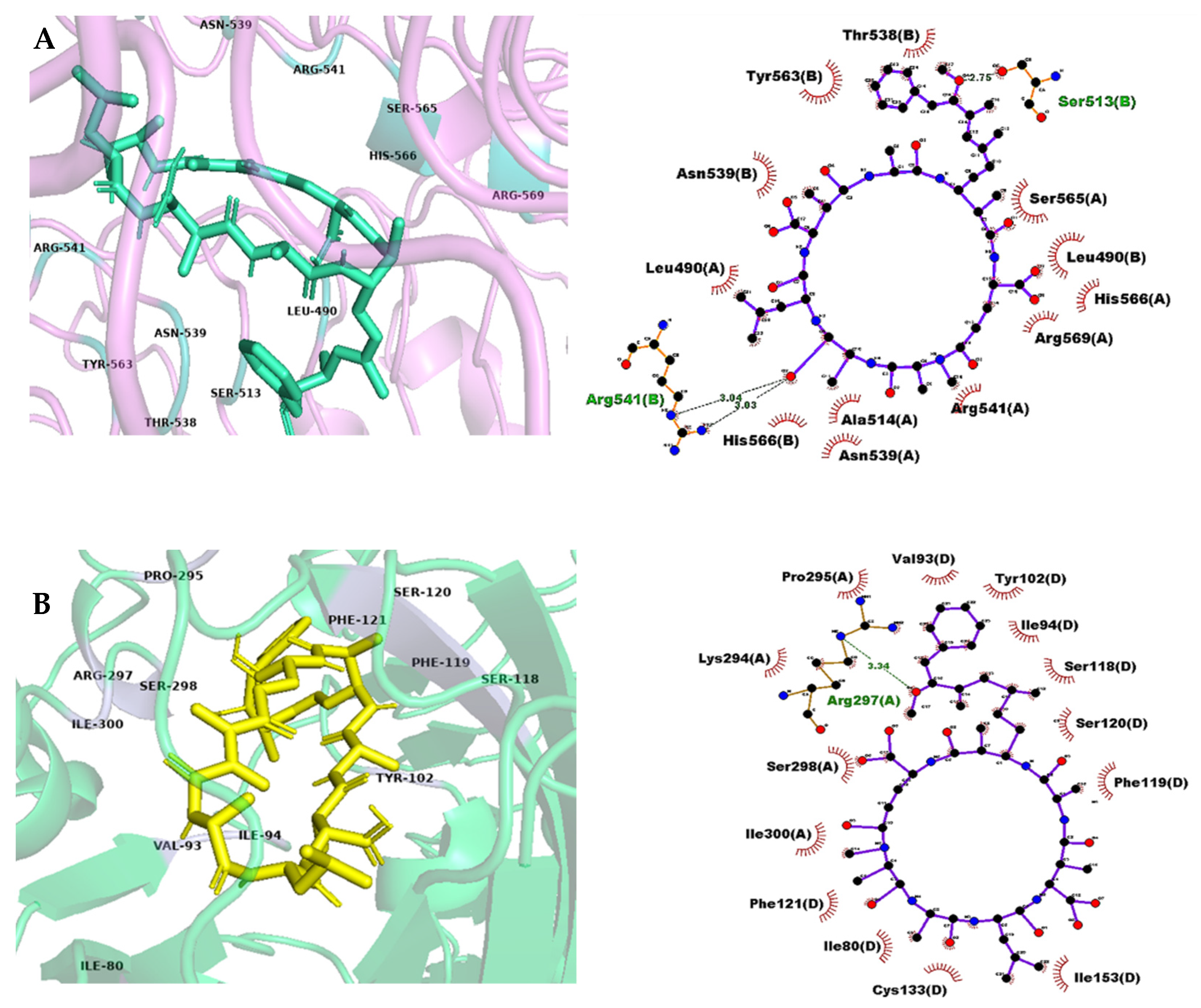
3.1.1. Metabolic Effects and Insulin Regulation
3.1.2. Interactions Related to Inflammatory Proteins and Neuroprotective Alteration
3.1.3. Nuclear Receptors and Genotoxicity Potential
3.1.4. Cellular Proliferation and Cancer
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.-L.; Utsumi, M. An overview of the accumulation of microcystins in aquatic ecosystems. J. Environ. Manag. 2018, 213, 520–529. [Google Scholar] [CrossRef]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Kaufononga, S.A.F.; Cary, S.C.; Hamilton, D.P. High levels of structural diversity observed in microcystins from Microcystis CAWBG11 and characterization of six new microcystin congeners. Mar. Drugs 2014, 12, 5372–5395. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Vasconcelos, V. Molecular mechanisms of microcystin toxicity in animal cells. Int. J. Mol. Sci. 2010, 11, 268–287. [Google Scholar] [CrossRef] [PubMed]
- Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A.; Benayache, N.Y.; Nguyen-Quang, T. Structural Diversity, Characterization and Toxicology of Microcystins. Toxins 2019, 11, 714. [Google Scholar] [CrossRef]
- Brózman, O.; Kubickova, B.; Babica, P.; Laboha, P. Microcystin-LR Does Not Alter Cell Survival and Intracellular Signaling in Human Bronchial Epithelial Cells. Toxins 2020, 12, 165. [Google Scholar] [CrossRef] [PubMed]
- Spoof, L.; Catherine, A. Appendix 3: Tables of Microcystins and Nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 526–537. ISBN 978-1-119-06876-1. [Google Scholar]
- Testai, E.; Buratti, F.M.; Funari, E.; Manganelli, M.; Vichi, S.; Arnich, N.; Biré, R.; Fessard, V.; Sialehaamoa, A. Review and analysis of occurrence, exposure and toxicity of cyanobacteria toxins in food. EFSA Support. Publ. 2016, 13, 998E. [Google Scholar] [CrossRef]
- Zegura, B.; Zajc, I.; Lah, T.T.; Filipic, M. Patterns of microcystin-LR induced alteration of the expression of genes involved in response to DNA damage and apoptosis. Toxicon 2008, 51, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Zegura, B.; Straser, A.; Filipič, M. Genotoxicity and potential carcinogenicity of cyanobacterial toxins—A review. Mutat. Res. 2011, 727, 16–41. [Google Scholar] [CrossRef]
- Zhong, S.; Liu, Y.; Wang, F.; Wu, Z.; Zhao, S. Microcystin-LR induced oxidative stress, inflammation, and apoptosis in alveolar type II epithelial cells of ICR mice in vitro. Toxicon 2020, 174, 19–25. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, K.; Zhou, Y.; Li, X.; Mao, X.; Wei, X.; Xu, Y.; Yang, W.; Chen, G.; Liu, C. Microcystin-LR impairs glucose metabolism in pancreatic β cells in vivo and in vitro. Toxicol. Lett. 2020, 326, 106–113. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Shen, S.; Wang, C.; Xiang, Z.; Han, X.; Li, D. Toxic effects of microcystin-LR on the development of prostate in mice. Toxicology 2017, 380, 50–61. [Google Scholar] [CrossRef]
- Bittner, M.; Štern, A.; Smutná, M.; Hilscherová, K.; Žegura, B. Cytotoxic and Genotoxic Effects of Cyanobacterial and Algal Extracts-Microcystin and Retinoic Acid Content. Toxins 2021, 13, 107. [Google Scholar] [CrossRef]
- Weng, D.; Lu, Y.; Wei, Y.; Liu, Y.; Shen, P. The role of ROS in microcystin-LR-induced hepatocyte apoptosis and liver injury in mice. Toxicology 2007, 232, 15–23. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Díez-Quijada, L.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Occurrence and toxicity of microcystin congeners other than MC-LR and MC-RR: A review. Food Chem. Toxicol. 2019, 125, 106–132. [Google Scholar] [CrossRef]
- Meyer, S.; Kehr, J.-C.; Mainz, A.; Dehm, D.; Petras, D.; Süssmuth, R.D.; Dittmann, E. Biochemical Dissection of the Natural Diversification of Microcystin Provides Lessons for Synthetic Biology of NRPS. Cell Chem. Biol. 2016, 23, 462–471. [Google Scholar] [CrossRef]
- Ohta, Y.; Kitanaka, A.; Mihara, K.; Imataki, O.; Ohnishi, H.; Tanaka, T.; Taminato, T.; Kubota, Y. Expression of CD38 with intracellular enzymatic activity: A possible explanation for the insulin release induced by intracellular cADPR. Mol. Cell. Biochem. 2011, 352, 293–299. [Google Scholar] [CrossRef]
- Lee, H.C. Structure and enzymatic functions of human CD38. Mol. Med. Camb. Mass 2006, 12, 317–323. [Google Scholar] [CrossRef]
- Liu, Q.; Kriksunov, I.A.; Graeff, R.; Munshi, C.; Lee, H.C.; Hao, Q.Q. Crystal Structure of Human CD38 Extracellular Domain. Structure 2005, 13, 1331–1339. [Google Scholar] [CrossRef]
- Morandi, F.; Airoldi, I.; Marimpietri, D.; Bracci, C.; Faini, A.C.; Gramignoli, R. CD38, a Receptor with Multifunctional Activities: From Modulatory Functions on Regulatory Cell Subsets and Extracellular Vesicles, to a Target for Therapeutic Strategies. Cells 2019, 8, 1527. [Google Scholar] [CrossRef] [PubMed]
- Ting, K.Y.; Leung, C.F.P.; Graeff, R.M.; Lee, H.C.; Hao, Q.; Kotaka, M. Porcine CD38 exhibits prominent secondary NAD(+) cyclase activity. Protein Sci. 2016, 25, 650–661. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Graeff, R.; Lee, H.C. Roles of cADPR and NAADP in pancreatic cells. Acta Biochim. Biophys. Sin. 2012, 44, 719–729. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Moreau, C.; Liu, Q.; Graeff, R.; Wagner, G.K.; Thomas, M.P.; Swarbrick, J.M.; Shuto, S.; Lee, H.C.; Hao, Q.; Potter, B.V.L. CD38 Structure-Based Inhibitor Design Using the N1-Cyclic Inosine 5′-Diphosphate Ribose Template. PLoS ONE 2013, 8, e66247. [Google Scholar] [CrossRef]
- Zhang, Q.; Qin, W.; Yang, L.; An, J.; Zhang, X.; Hong, H.; Xu, L.; Wang, Y. Microcystis bloom containing microcystin-LR induces type 2 diabetes mellitus. Toxicol. Lett. 2018, 294, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, S.R.; Butler, L.M.; Hoy, A.J. The diversity and breadth of cancer cell fatty acid metabolism. Cancer Metab. 2021, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Zwang, Y.; Yarden, Y. p38 MAP kinase mediates stress-induced internalization of EGFR: Implications for cancer chemotherapy. EMBO J. 2006, 25, 4195–4206. [Google Scholar] [CrossRef]
- Zastepa, A.; Pick, F.R.; Blais, J.M. Fate and Persistence of Particulate and Dissolved Microcystin-LA from Microcystis Blooms. Hum. Ecol. Risk Assess. Int. J. 2014, 20, 1670–1686. [Google Scholar] [CrossRef]
- Moriya, C.; Jinnin, M.; Yamane, K.; Maruo, K.; Muchemwa, F.C.; Igata, T.; Makino, T.; Fukushima, S.; Ihn, H. Expression of matrix metalloproteinase-13 is controlled by IL-13 via PI3K/Akt3 and PKC-δ in normal human dermal fibroblasts. J. Investig. Dermatol. 2011, 131, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.L.; Maroulakou, I.G.; Eldridge, J.; Liby, T.L.; Sridharan, V.; Tsichlis, P.N.; Muise-Helmericks, R.C. VEGF stimulation of mitochondrial biogenesis: Requirement of AKT3 kinase. FASEB J. 2008, 22, 3264–3275. [Google Scholar] [CrossRef]
- Pereira, S.R.; Vasconcelos, V.M.; Antunes, A. Computational study of the covalent bonding of microcystins to cysteine residues—A reaction involved in the inhibition of the PPP family of protein phosphatases. FEBS J. 2013, 280, 674–680. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xie, P. Mechanisms of Microcystin-induced Cytotoxicity and Apoptosis. Mini-Rev. Med. Chem. 2016, 16, 1018–1031. [Google Scholar] [CrossRef] [PubMed]
- Maynes, J.T.; Luu, H.A.; Cherney, M.M.; Andersen, R.J.; Williams, D.; Holmes, C.F.B.; James, M.N.G. Crystal Structures of Protein Phosphatase-1 Bound to Motuporin and Dihydromicrocystin-LA: Elucidation of the Mechanism of Enzyme Inhibition by Cyanobacterial Toxins. J. Mol. Biol. 2006, 356, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Craig, M.; Luu, H.A.; McCready, T.L.; Holmes, C.F.B.; Williams, D.; Andersen, R.J. Molecular mechanisms underlying the interaction of motuporin and microcystins with type-1 and type-2A protein phosphatases. Biochem. Cell Biol. 1996, 74, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Hastie, C.; Borthwick, E.; Morrison, L.; Codd, G.; Cohen, P. Inhibition of several protein phosphatases by a non-covalently interacting microcystin and a novel cyanobacterial peptide, nostocyclin. Biochim. Biophys. Acta BBA-Gen. Subj. 2005, 1726, 187–193. [Google Scholar] [CrossRef]
- Chen, Y.-M.; Lee, T.-H.; Lee, J.; Huang, H.-B.; Huang, R.; Chou, H.-N. Comparison of protein phosphatase inhibition activities and mouse toxicities of microcystins. Toxicon 2006, 47, 742–746. [Google Scholar] [CrossRef]
- Zhou, M.; Tu, W.; Xu, J. Mechanisms of microcystin-LR-induced cytoskeletal disruption in animal cells. Toxicon 2015, 101, 92–100. [Google Scholar] [CrossRef]
- Pope, C.N.; Brimijoin, S. Cholinesterases and the fine line between poison and remedy. Biochem. Pharmacol. 2018, 153, 205–216. [Google Scholar] [CrossRef]
- Li, Q.; Yang, H.; Chen, Y.; Sun, H. Recent progress in the identification of selective butyrylcholinesterase inhibitors for Alzheimer’s disease. Eur. J. Med. Chem. 2017, 132, 294–309. [Google Scholar] [CrossRef]
- Yen, Y.-C.; Kammeyer, A.M.; Jensen, K.C.; Tirlangi, J.; Ghosh, A.K.; Mesecar, A.D. Development of an Efficient Enzyme Production and Structure-Based Discovery Platform for BACE1 Inhibitors. Biochemistry 2019, 58, 4424–4435. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, Y.; Zhang, C.; Xiang, Z.; Ding, J.; Han, X. Learning and memory deficits and alzheimer’s disease-like changes in mice after chronic exposure to microcystin-LR. J. Hazard. Mater. 2019, 373, 504–518. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-B.; Zhang, X.; Ju, J.; Li, Y.; Yin, L.; Pu, Y. Alterations in neurobehaviors and inflammation in hippocampus of rats induced by oral administration of microcystin-LR. Environ. Sci. Pollut. Res. Int. 2014, 21, 12419–12425. [Google Scholar] [CrossRef] [PubMed]
- Lampón, N.; Hermida-Cadahia, E.F.; Riveiro, A.; Tutor, J.C. Association between butyrylcholinesterase activity and low-grade systemic inflammation. Ann. Hepatol. 2012, 11, 356–363. [Google Scholar] [CrossRef]
- Lockridge, O. Review of human butyrylcholinesterase structure, function, genetic variants, history of use in the clinic, and potential therapeutic uses. Pharmacol. Ther. 2015, 148, 34–46. [Google Scholar] [CrossRef] [PubMed]
- McGettrick, A.F.; O’Neill, L.A.J. Localisation and trafficking of Toll-like receptors: An important mode of regulation. Curr. Opin. Immunol. 2010, 22, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Adegoke, E.O.; Wang, C.; Machebe, N.S.; Wang, X.; Wang, H.; Adeniran, S.O.; Zhang, H.; Zheng, P.; Zhang, G. Microcystin-leucine arginine (MC-LR) induced inflammatory response in bovine sertoli cell via TLR4/NF-kB signaling pathway. Environ. Toxicol. Pharmacol. 2018, 63, 115–126. [Google Scholar] [CrossRef]
- Lin, W.; Guo, H.; Wang, L.; Zhang, D.; Wu, X.; Li, L.; Qiu, Y.; Yang, L.; Li, D.; Tang, R. Waterborne microcystin-LR exposure induced chronic inflammatory response via MyD88-dependent toll-like receptor signaling pathway in male zebrafish. Sci. Total Environ. 2020, 702, 134969. [Google Scholar] [CrossRef] [PubMed]
- Nunes, K.P.; de Oliveira, A.A.; Mowry, F.E.; Biancardi, V.C. Targeting toll-like receptor 4 signalling pathways: Can therapeutics pay the toll for hypertension? Br. J. Pharmacol. 2019, 176, 1864–1879. [Google Scholar] [CrossRef] [PubMed]
- Tanji, H.; Ohto, U.; Shibata, T.; Miyake, K.; Shimizu, T. Structural reorganization of the Toll-like receptor 8 dimer induced by agonistic ligands. Science 2013, 339, 1426–1429. [Google Scholar] [CrossRef]
- Wang, X.; Huang, P.; Liu, Y.; Du, H.; Wang, X.; Wang, M.; Wang, Y.; Hei, T.K.; Wu, L.; Xu, A. Role of nitric oxide in the genotoxic response to chronic microcystin-LR exposure in human-hamster hybrid cells. J. Environ. Sci. China 2015, 29, 210–218. [Google Scholar] [CrossRef]
- Polakovičová, M.; Jampílek, J. Advances in Structural Biology of ACE and Development of Domain Selective ACE-inhibitors. Med. Chem. 2019, 15, 574–587. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Massey, I.Y.; Feng, H.; Yang, F. A Review of Cardiovascular Toxicity of Microcystins. Toxins 2019, 11, 507. [Google Scholar] [CrossRef]
- Shi, J.; Zhou, J.; Zhang, M. Microcystins Induces Vascular Inflammation in Human Umbilical Vein Endothelial Cells via Activation of NF-κB. Mediat. Inflamm. 2015, 2015, 942159. [Google Scholar] [CrossRef] [PubMed]
- Alverca, E.; Andrade, M.; Dias, E.; Sam Bento, F.; Batoréu, M.C.C.; Jordan, P.; Silva, M.J.; Pereira, P. Morphological and ultrastructural effects of microcystin-LR from Microcystis aeruginosa extract on a kidney cell line. Toxicon 2009, 54, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Power, D.M.; Elias, N.P.; Richardson, S.J.; Mendes, J.; Soares, C.M.; Santos, C.R. Evolution of the thyroid hormone-binding protein, transthyretin. Gen. Comp. Endocrinol. 2000, 119, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Liu, Z.; Gao, Y.; Jia, D.; Tang, R.; Li, L.; Li, D. Waterborne exposure to microcystin-LR alters thyroid hormone levels, iodothyronine deiodinase activities, and gene transcriptions in juvenile zebrafish (Danio rerio). Chemosphere 2020, 241, 125037. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, Z.; Jia, D.; Hu, Q.; Li, L.; Tang, R.; Li, D. Acute microcystin-LR exposure interfere thyroid hormones homeostasis in adult zebrafish (Danio rerio). Chemosphere 2020, 243, 125258. [Google Scholar] [CrossRef]
- Nair, P.C.; McKinnon, R.A.; Miners, J.O. Cytochrome P450 structure-function: Insights from molecular dynamics simulations. Drug Metab. Rev. 2016, 48, 434–452. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Suganuma, M. Tumor Promoters—Microcystin-LR, Nodularin and TNF-α and Human Cancer Development. Anticancer Agents Med. Chem. 2011, 11, 4–18. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Y.; Zuo, X.; Ding, N.; Zeng, H.; Zou, X.; Han, X. Microcystin (-LR) induced testicular cell apoptosis via up-regulating apoptosis-related genes in vivo. Food Chem. Toxicol. 2013, 60, 309–317. [Google Scholar] [CrossRef]
- Dias, E.; Andrade, M.; Alverca, E.; Pereira, P.; Batoréu, M.C.C.; Jordan, P.; Silva, M.J. Comparative study of the cytotoxic effect of microcistin-LR and purified extracts from Microcystis aeruginosa on a kidney cell line. Toxicon 2009, 53, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Karthikeyan, C.; Moorthy, N.S.H.N.; Waiker, D.K.; Jain, A.K.; Trivedi, P. Human CDC2-like kinase 1 (CLK1): A novel target for Alzheimer’s disease. Curr. Drug Targets 2014, 15, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Honma, M.; Shibuya, T.; Fujii, M.; Iinuma, S.; Ishida-Yamamoto, A. Aberrant LIM-kinase 1 expression in hyperproliferative psoriatic epidermis. J. Dermatol. 2017, 44, 91–92. [Google Scholar] [CrossRef]
- Chen, Q.; Gimple, R.C.; Li, G.; Chen, J.; Wu, H.; Li, R.; Xie, J.; Xu, B. LIM kinase 1 acts as a profibrotic mediator in permanent atrial fibrillation patients with valvular heart disease. J. Biosci. 2019, 44, 16. [Google Scholar] [CrossRef] [PubMed]
- Cebula, M.; Schmidt, E.E.; Arnér, E.S.J. TrxR1 as a Potent Regulator of the Nrf2-Keap1 Response System. Antioxid. Redox Signal. 2015, 23, 823–853. [Google Scholar] [CrossRef]
- Woynarowska, B.A.; Woynarowski, J.M. Preferential targeting of apoptosis in tumor versus normal cells. Biochim. Biophys. Acta 2002, 1587, 309–317. [Google Scholar] [CrossRef]
- Schreidah, C.M.; Ratnayake, K.; Senarath, K.; Karunarathne, A. Microcystins: Biogenesis, Toxicity, Analysis, and Control. Chem. Res. Toxicol. 2020, 33, 2225–2246. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
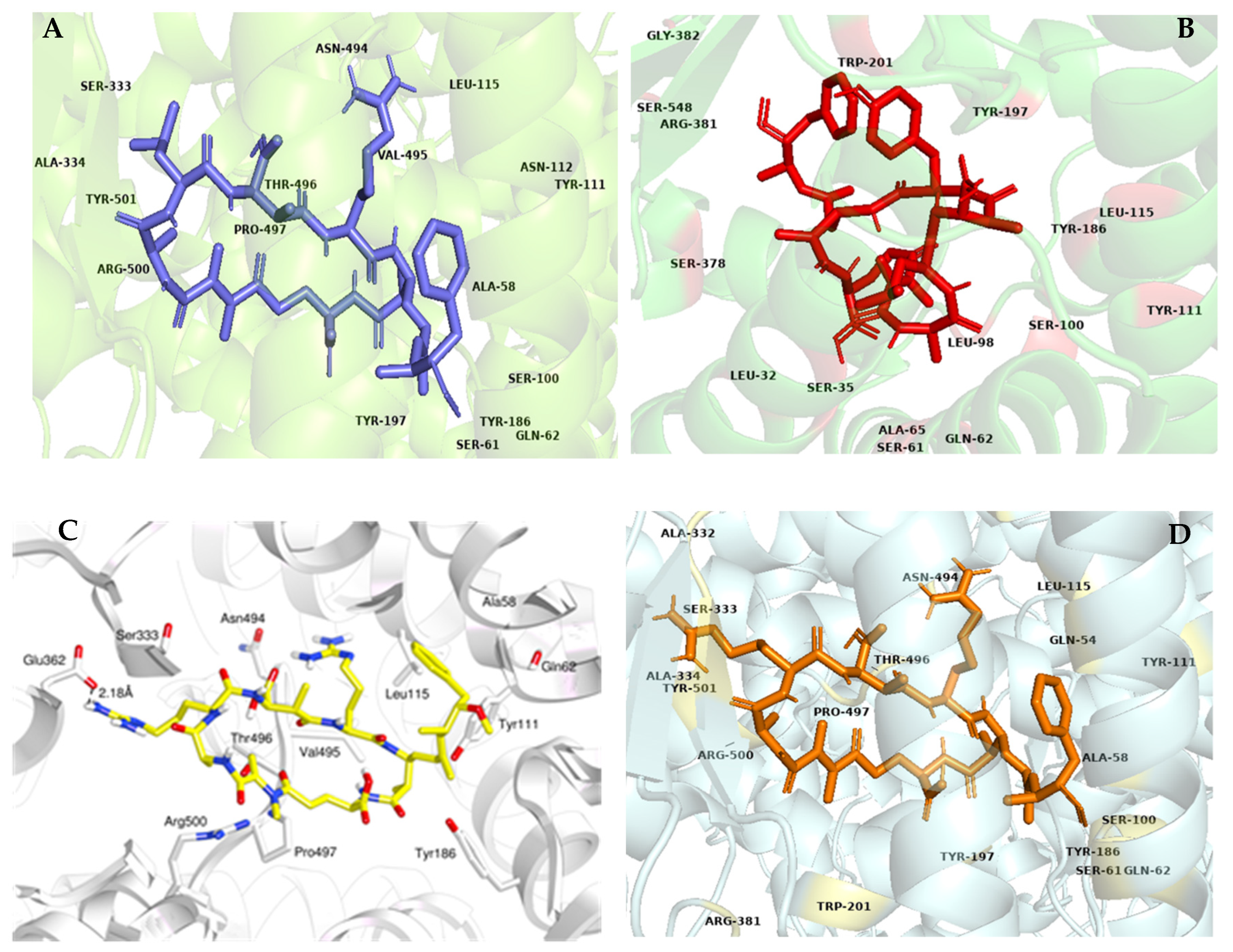{kind=link}
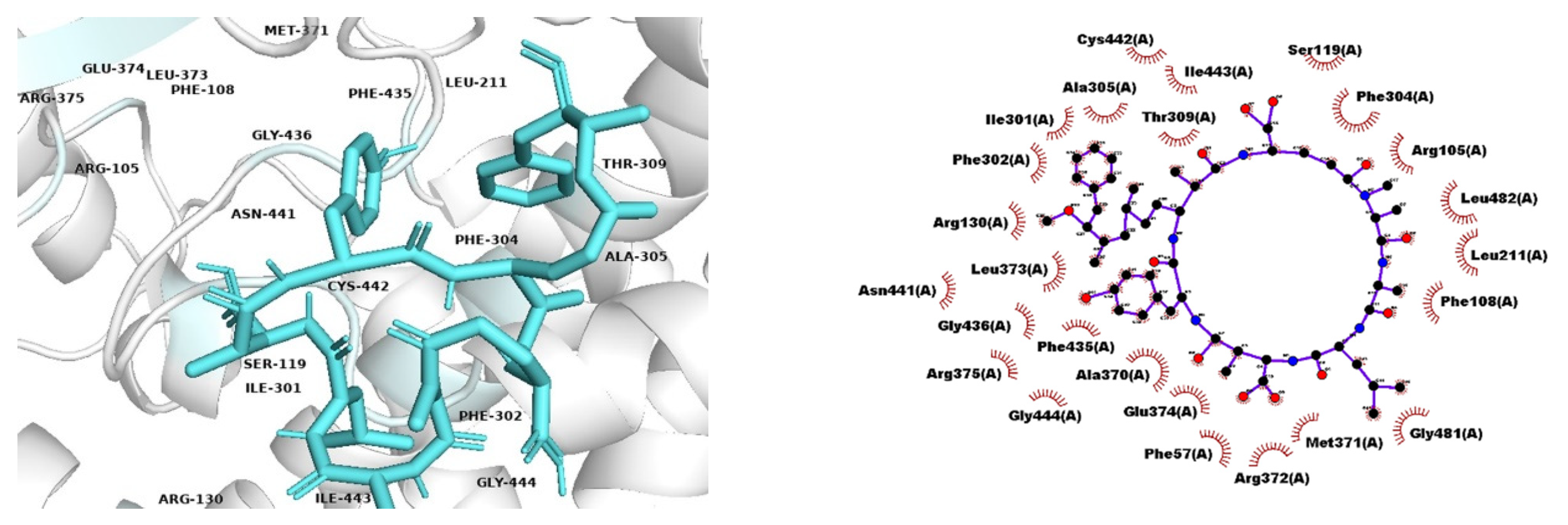{kind=link}
| Short Name | PDB-ID | MC-LA | MC-LR | MC-RR | MC-LY | |
|---|---|---|---|---|---|---|
| ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase 1 | CD38 | 3ofs | −10.1 | −10.1 | −10.1 | −11.5 |
| Butyrylcholine esterase | BChE | 4xii | −10.5 | −10.4 | −10.4 | −11.0 |
| Toll-like receptor 4 | TLR4 | 3ula | −10.3 | −10.0 | −10.1 | −11.4 |
| Toll-like receptor 8 | TLR8 | 3w3j | −11.5 | −10.4 | −10.3 | −11.3 |
| Angiotensin-I-converting enzyme | ACE | 3nxq | −11.1 | −11.3 | −11.5 | −11.3 |
| DNA topoisomerase 2-alpha | TOP2A | 1zxm | −10.7 | −10.6 | −10.6 | −11.1 |
| Induced myeloid leukemia cell differentiation protein Mcl-1 | MCL-1 | 5lof | −10.7 | −10.9 | −10.2 | −10.6 |
| Hemoglobin subunit alpha | HBA | 3b75 | −11.2 | −10.2 | −11.4 | −11.0 |
| Apoptosis regulator Bcl-2 | BCL2 | 2w3l | −10.6 | −10.6 | −10.2 | −10.9 |
| Heat shock protein HSP 90-alpha | HSP90 | 2qg0 | −10.2 | −10.1 | −10.9 | −10.9 |
| Serine/threonine-protein kinase 26 | MST-4 | 3ggf | −11.0 | −10.5 | −10.2 | −10.9 |
| Catalase | CAT | 1dgb | −10.7 | −10.4 | −10.4 | −10.7 |
| Transthyretin | TTR | 1ict | −10.8 | −10.2 | −10.2 | −10.6 |
| Calcium/calmodulin-dependent serine/threonine kinase | DAPK | 1jkl | −10.0 | −10.5 | −10.2 | −10.6 |
| Aminopeptidase N | AMPN | 4fyr | −10.5 | −10.3 | −10.3 | −10.3 |
| MC-LA, MC-LR MC-RR, MC-LY | MC-LA, MC-LR, MC-RR | MC-LR, MC-RR, MC-LY | MC-LR, MC-RR | MC-LA, MC-RR | MC-LR, MC-LY |
|---|---|---|---|---|---|
| Gln 62 | Ala 58 | Ser 61 | Gln 54 | * Gln 362 | Tyr 197 |
| * Tyr 111 | Ser 333 | Ser 100 | Ala 332 | * Thr 496 | |
| Leu 115 | Asn 494 | Trp 201 | Ala 334 | ||
| Tyr 186 | Val 495 | * Arg 381 | Tyr 501 | ||
| Pro 497 | |||||
| Arg 500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tapia-Larios, C.; Mendoza, E.G.; Rubio-Armendáriz, M.C.; Olivero-Verbel, J. Identification of Novel Molecular Targets of Four Microcystin Variants by High-Throughput Virtual Screening. Appl. Sci. 2022, 12, 2612. https://doi.org/10.3390/app12052612
Tapia-Larios C, Mendoza EG, Rubio-Armendáriz MC, Olivero-Verbel J. Identification of Novel Molecular Targets of Four Microcystin Variants by High-Throughput Virtual Screening. Applied Sciences. 2022; 12(5):2612. https://doi.org/10.3390/app12052612
Chicago/Turabian StyleTapia-Larios, Claudia, Ernesto García Mendoza, María Carmen Rubio-Armendáriz, and Jesus Olivero-Verbel. 2022. "Identification of Novel Molecular Targets of Four Microcystin Variants by High-Throughput Virtual Screening" Applied Sciences 12, no. 5: 2612. https://doi.org/10.3390/app12052612
APA StyleTapia-Larios, C., Mendoza, E. G., Rubio-Armendáriz, M. C., & Olivero-Verbel, J. (2022). Identification of Novel Molecular Targets of Four Microcystin Variants by High-Throughput Virtual Screening. Applied Sciences, 12(5), 2612. https://doi.org/10.3390/app12052612