
Benincasa hispida Extract Promotes Proliferation, Differentiation, and Mineralization of MC3T3-E1 Preosteoblasts and Inhibits the Differentiation of RAW 246.7 Osteoclast Precursors


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sample Preparation
2.3. Chemical Component Characterization
2.4. HPLC Analysis of 2-Furoic Acid
2.5. Osteoblast Cell Culture
2.6. Osteoblast Cell Proliferation
2.7. Analysis of ALP Activity
2.8. Western Blotting
2.9. Osteoclast Cell Culture
2.10. Cell Viability
2.11. TRAP Assay
2.12. Statistical Analyses
3. Results
3.1. Identification of Chemical Components in HR1901-W by Preparative HPLC and NMR
3.2. Analysis of the 2-Furoic Acid Concentration in HR1901-W
3.3. Effect of HR1901-W on the Proliferation of MC3T3-E1 Preosteoblasts and ALP Activity
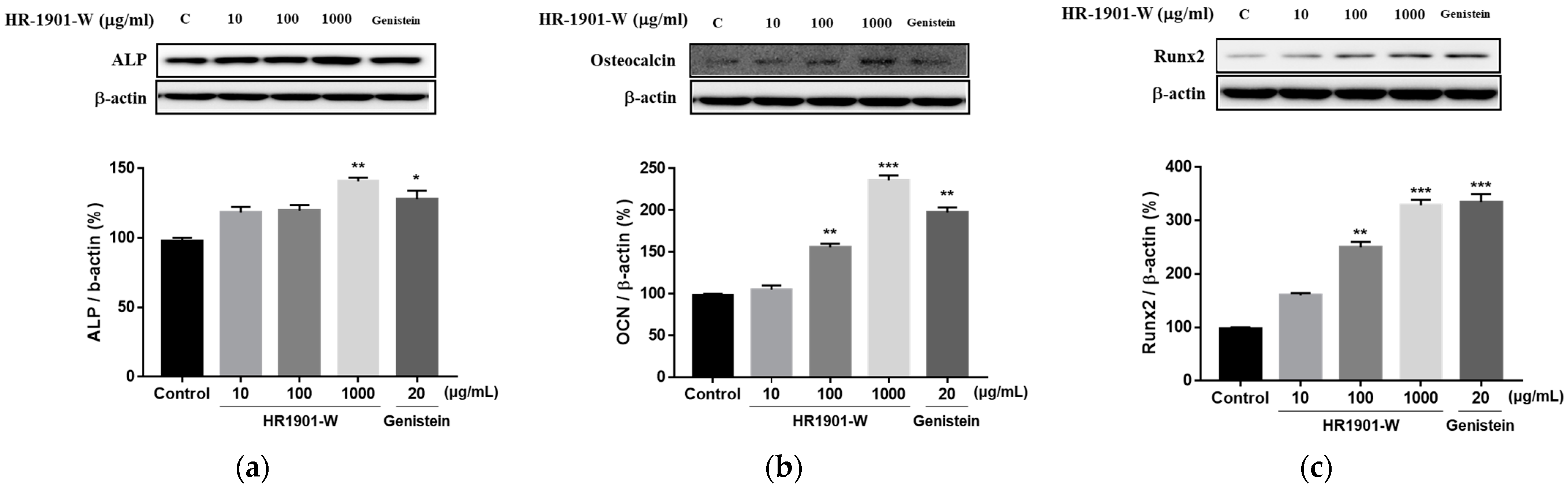
3.4. Effects of HR1901-W on Expression Levels of Factors Related to Osteoblast Differentiation
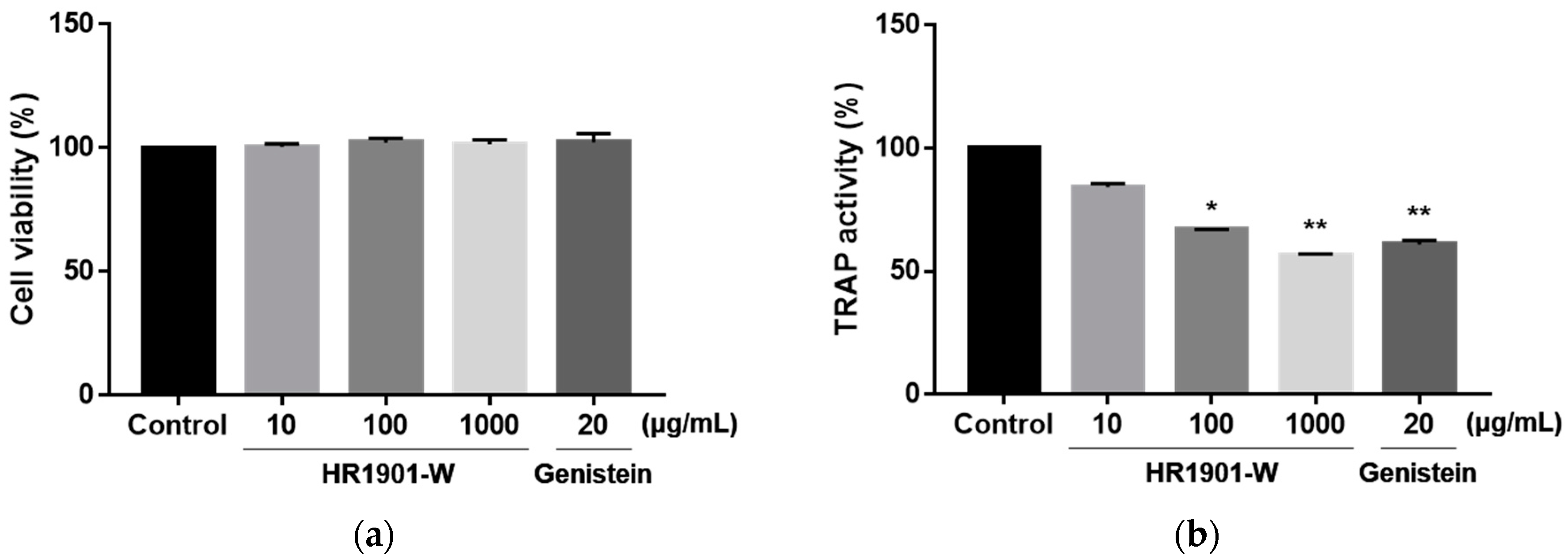
3.5. Effects of HR1901-W on Osteoclast Cell Viability and TRAP Activity
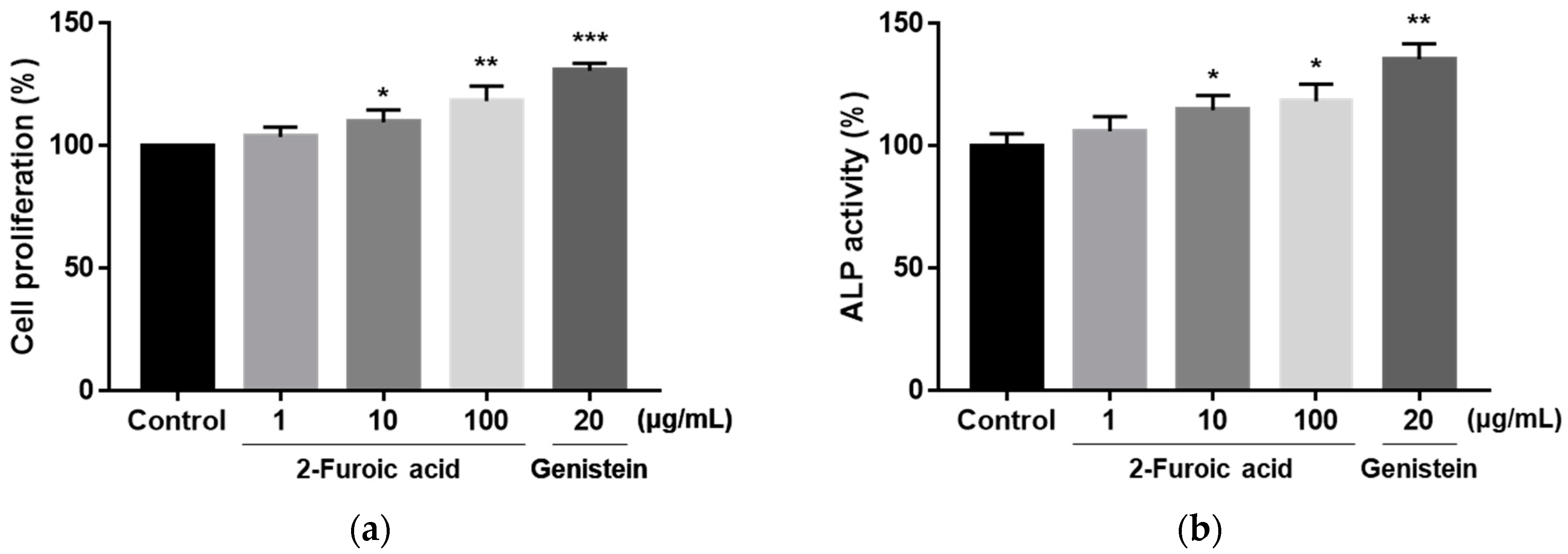
3.6. Effect of 2-Furoic Acid on the Proliferation of MC3T3-E1 Preosteoblasts and ALP Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bonewald, L.F. The amazing osteocyte. J. Bone Miner. Res. 2011, 26, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Datta, H.K.; Ng, W.F.; Walker, J.A.; Tuck, S.P.; Varanasi, S.S. The cell biology of bone metabolism. J. Clin. Pathol. 2008, 61, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast–osteoclast interactions. Connect. Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. Osteoclasts: What do they do and how do they do it? Am. J. Pathol. 2007, 170, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Hernlund, E.; Svedbom, A.; Ivergård, M.; Compston, J.; Cooper, C.; Stenmark, J.; McCloskey, E.V.; Jönsson, B.; Kanis, J.A. Osteoporosis in the european union: Medical management, epidemiology and economic burden. Arch. Osteoporos. 2013, 8, 136. [Google Scholar] [CrossRef]
- Tolar, J.; Teitelbaum, S.L.; Orchard, P.J. Osteopetrosis. N. Engl. J. Med. 2004, 351, 2839–2849. [Google Scholar] [CrossRef]
- Freedman, K.B.; Kaplan, F.S.; Bilker, W.B.; Strom, B.L.; Lowe, R.A. Treatment of osteoporosis: Are physicians missing an opportunity? JBJS 2000, 82, 1063. [Google Scholar] [CrossRef]
- Marcus, R.; Wong, M.; Heath, H., III; Stock, J.L. Antiresorptive treatment of postmenopausal osteoporosis: Comparison of study designs and outcomes in large clinical trials with fracture as an endpoint. Endocr. Rev. 2002, 23, 16–37. [Google Scholar] [CrossRef]
- McClung, M.R.; Geusens, P.; Miller, P.D.; Zippel, H.; Bensen, W.G.; Roux, C.; Adami, S.; Fogelman, I.; Diamond, T.; Eastell, R.; et al. Effect of risedronate on the risk of hip fracture in elderly women. N. Engl. J. Med. 2001, 344, 333–340. [Google Scholar] [CrossRef]
- Woo, S.; Hellstein, J.W.; Kalmar, J.R. Systematic review: Bisphosphonates and osteonecrosis of the jaws. Ann. Intern. Med. 2006, 144, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, R.; Hata, K.; Matsubara, T.; Wakabayashi, M.; Yoneda, T. Regulation of bone and cartilage development by network between BMP signalling and transcription factors. J. Biochem. 2012, 151, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Tsang, K.Y.; Tang, H.C.; Chan, D.; Cheah, K.S. Hypertrophic chondrocytes can become osteoblasts and osteocytes in endochondral bone formation. Proc. Natl. Acad. Sci. USA 2014, 111, 12097–12102. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of osteoblast differentiation by Runx2. In Osteoimmunology; Springer: Berlin/Heidelberg, Germany, 2009; pp. 43–49. [Google Scholar]
- Komori, T. Regulation of bone development and extracellular matrix protein genes by RUNX2. Cell Tissue Res. 2010, 339, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Franzen, A.; Heinegård, D. Isolation and characterization of two sialoproteins present only in bone calcified matrix. Biochem. J. 1985, 232, 715–724. [Google Scholar] [CrossRef]
- Owen, T.A.; Aronow, M.; Shalhoub, V.; Barone, L.M.; Wilming, L.; Tassinari, M.S.; Kennedy, M.B.; Pockwinse, S.; Lian, J.B.; Stein, G.S. Progressive development of the rat osteoblast phenotype in vitro: Reciprocal relationships in expression of genes associated with osteoblast proliferation and differentiation during formation of the bone extracellular matrix. J. Cell. Physiol. 1990, 143, 420–430. [Google Scholar] [CrossRef]
- Beertsen, W.; Van den Bos, T. Alkaline phosphatase induces the mineralization of sheets of collagen implanted subcutaneously in the rat. J. Clin. Investig. 1992, 89, 1974–1980. [Google Scholar] [CrossRef]
- Delmas, P.D.; Eastell, R.; Garnero, P.; Seibel, M.J.; Stepan, J.; Committee of Scientific Advisors of the International Osteoporosis Foundation. The use of biochemical markers of bone turnover in osteoporosis. Osteoporos. Int. 2000, 11, S2. [Google Scholar] [CrossRef]
- Mizokami, A.; Kawakubo-Yasukochi, T.; Hirata, M. Osteocalcin and its endocrine functions. Biochem. Pharmacol. 2017, 132, 1–8. [Google Scholar] [CrossRef]
- Watanabe, K.; Ikeda, K. Osteoblast differentiation and bone formation. Nihon Rinsho. Jpn. J. Clin. Med. 2009, 67, 879–886. [Google Scholar]
- Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kurata, K.; Oh-Hora, M.; Feng, J.Q.; Bonewald, L.F.; Kodama, T.; Wutz, A.; Wagner, E.F.; et al. Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat. Med. 2011, 17, 1231–1234. [Google Scholar] [CrossRef]
- Xiong, J.; Onal, M.; Jilka, R.L.; Weinstein, R.S.; Manolagas, S.C.; O’brien, C.A. Matrix-embedded cells control osteoclast formation. Nat. Med. 2011, 17, 1235–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karsdal, M.A.; Hjorth, P.; Henriksen, K.; Kirkegaard, T.; Nielsen, K.L.; Lou, H.; Delaissé, J.-M.; Foged, N.T. Transforming growth factor-β controls human osteoclastogenesis through the p38 MAPK and regulation of RANK expression. J. Biol. Chem. 2003, 278, 44975–44987. [Google Scholar] [PubMed]
- Chopra, R.N. Glossary of Indian Medicinal Plants; CSIR: New Delhi, India, 1956. [Google Scholar]
- Nanjing University of Traditional Chinese Medicine. Dictionary of Traditional Chinese Medicine; Nanjing University of Traditional Chinese Medicine: Nanjing, China, 2006. [Google Scholar]
- Sahu, P.K.; Sharma, D.; Nair, S.K. Performance of ash gourd genotypes for earliness and yield under chhattisgarh plains, india. Plant Arch. 2015, 15, 1157–1160. [Google Scholar]
- Siong, T.E.; Noor, M.I.; Azudin, M.N.; Idris, K. Nutrient Composition of Malaysian Foods; ASEAN Sub-Committee on Protein: Kuala Lumpur, Malaysia, 1988. [Google Scholar]
- Du, Q.; Zhang, Q.; Ito, Y. Isolation and identification of phenolic compounds in the fruit of benincasa hispida by HSCCC. J. Liq. Chromatogr. Relat. Technol. 2005, 28, 137–144. [Google Scholar] [CrossRef]
- Gill, N.S.; Dhiman, K.; Bajwa, J.; Sharma, P.; Sood, S. Evaluation of free radical scavenging, anti-inflammatory and analgesic potential of benincasa hispida seed extract. IJP-Int. J. Pharmacol. 2010, 6, 652–657. [Google Scholar] [CrossRef]
- Yoshizumi, S.; Murakami, T.; Kadoya, M.; Matsuda, H.; Yamahara, J.; Yoshikawa, M. Medicinal foodstuffs. XI. histamine release inhibitors from wax gourd, the fruits of benincasa hispida cogn. Yakugaku Zasshi J. Pharm. Soc. Jpn. 1998, 118, 188–192. [Google Scholar] [CrossRef]
- Grover, J.K.; Adiga, G.; Vats, V.; Rathi, S.S. Extracts of benincasa hispida prevent development of experimental ulcers. J. Ethnopharmacol. 2001, 78, 159–164. [Google Scholar] [CrossRef]
- Grover, J.K.; Rathi, S.S. Benincasa hispida: An anti-inflammatory agent with cytoprotective activity (abs). Can. J. Physiol. Pharmaol. 1994, 72, 269. [Google Scholar]
- Patil, R.N.; Patil, R.Y.; Ahirwar, B.; Ahirwar, D. Evaluation of antidiabetic and related actions of some indian medicinal plants in diabetic rats. Asian Pac. J. Trop. Med. 2011, 4, 20–23. [Google Scholar]
- Rachchh, M.A.; Jain, S.M. Gastroprotective effect of benincasa hispida fruit extract. Indian J. Pharmacol. 2008, 40, 271. [Google Scholar] [CrossRef]
- Roy, C.; Guha, D. Role of benincasa hispida linn. on brain electrical activity in colchicine induced experimental rat model of alzheimer’s disease: Possible involvements of antioxidants. World J. Pharm. Res. 2017, 6, 1439–1443. [Google Scholar]
- Mandal, U.; De, D.; Ali, K.M.; Biswas, A.; Ghosh, D. Effect of different solvent extracts of benincasa hispida T. on experimental hypochlorhydria in rat. J. Adv. Pharm. Technol. Res. 2012, 3, 41. [Google Scholar] [PubMed]
- Tan, Z.B.; Tonks, C.E.; O’Donnell, G.E.; Geyer, R. An improved HPLC analysis of the metabolite furoic acid in the urine of workers occupationally exposed to furfural. J. Anal. Toxicol. 2003, 27, 43–46. [Google Scholar] [PubMed]
- Vimalraj, V.; Vijayalakshmi, S.; Umayaparvathi, S.; Krishnan, A.R. Vibrational, NMR spectral studies of 2-furoic hydrazide by DFT and ab initio HF methods. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2011, 78, 670–675. [Google Scholar]
- Feng, X.; McDonald, J.M. Disorders of bone remodeling. Annu. Rev. Pathol. 2011, 6, 121. [Google Scholar]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar]
- Jiang, X.; Kuang, F.; Kong, F.; Yan, C. Prediction of the antiglycation activity of polysaccharides from Benincasa hispida using a response surface methodology. Carbohydr. Polym. 2016, 151, 358–363. [Google Scholar]
- Fatariah, Z.; Zulkhairuazha, T.Y.T.G.; Rosli, W.W. Quantitative HPLC analysis of gallic acid in Benincasa hispida prepared with different extraction techniques. Sains Malays. 2014, 43, 1181–1187. [Google Scholar]
- Hall, I.H.; Williams, W.L., Jr.; Rhyne, K.A.; Knowles, M. The hypolipidemic activity of furoic acid and furylacrylic acid derivatives in rodents. Pharm. Res. 1985, 2, 233–238. [Google Scholar]
- Hall, I.H.; Wong, O.T.; Reynolds, D.J.; Chang, J.J. Hypolipidemic Effects of 2-Furoic Acid in Sprague-Dawley Rats. Arch. Pharm. 1993, 326, 15–23. [Google Scholar]
- Kim, M.K.; Kim, H.D.; Park, J.H.; Lim, J.I.; Yang, J.S.; Kwak, W.Y.; Sung, S.Y.; Kim, S.H.; Lee, C.H.; Shim, J.Y.; et al. An orally active cathepsin K inhibitor, furan-2-carboxylic acid, 1-{1-[4-fluoro-2-(2-oxo-pyrrolidin-1-yl)-phenyl]-3-oxo-piperidin-4-ylcarbamoyl}-cyclohexyl)-amide (OST-4077), inhibits osteoclast activity in vitro and bone loss in ovariectomized rats. J. Pharmacol. Exp. Ther. 2006, 318, 555–562. [Google Scholar] [PubMed]
- Kandikattu, H.K.; Venuprasad, M.P.; Pal, A.; Khanum, F. Phytochemical analysis and exercise enhancing effects of hydroalcoholic extract of Celastrus paniculatus Willd. Ind. Crops Prod. 2014, 55, 217–224. [Google Scholar]
- Gullón, B.; Eibes, G.; Dávila, I.; Moreira, M.T.; Labidi, J.; Gullón, P. Hydrothermal treatment of chestnut shells (Castanea sativa) to produce oligosaccharides and antioxidant compounds. Carbohydr. Polym. 2018, 192, 75–83. [Google Scholar]
- Avila-Gaxiola, E.; Avila-Gaxiola, J.; Velarde-Escobar, O.; Ramos-Brito, F.; Atondo-Rubio, G.; Yee-Rendon, C. Effect of drying temperature on Agave tequilana leaves: A pretreatment for releasing reducing sugars for biofuel production. J. Food Process Eng. 2017, 40, e12455. [Google Scholar]
- Avila-Gaxiola, J.C.; Avila-Gaxiola, E. Ethanol production from Agave tequilana leaves powder by Saccharomyces cerevisiae yeast applying enzymatic saccharification without detoxification. Ind. Crops Prod. 2022, 177, 114515. [Google Scholar]
- Bonnick, S.L. Osteoporosis in men and women. Clin. Cornerstone 2006, 8, 28–39. [Google Scholar]
- Papaioannou, A.; Morin, S.; Cheung, A.M.; Atkinson, S.; Brown, J.P.; Feldman, S.; Hanley, D.A.; Hodsman, A.; Jamal, S.A.; Kaiser, S.M.; et al. 2010 clinical practice guidelines for the diagnosis and management of osteoporosis in Canada: Summary. CMAJ 2010, 182, 1864–1873. [Google Scholar] [CrossRef] [Green Version]
- Hooshmand, S.; Juma, S.; Arjmandi, B.H. Combination of genistin and fructooligosaccharides prevents bone loss in ovarian hormone deficiency. J. Med. Food 2010, 13, 320–325. [Google Scholar]
- Zittermann, A.; Geppert, J.; Zehn, N.; Gouni-Berthold, I.; Berthold, H.K.; Reinsberg, J.; Stehle, P.; Baier, S. Short-term effects of high soy supplementation on sex hormones, bone markers, and lipid parameters in young female adults. Eur. J. Nutr. 2004, 43, 100–108. [Google Scholar]
- Morabito, N.; Crisafulli, A.; Vergara, C.; Gaudio, A.; Lasco, A.; Frisina, N.; D’Anna, R.; Corrado, F.; Pizzoleo, M.A.; Cincotta, M.; et al. Effects of genistein and hormone-replacement therapy on bone loss in early postmenopausal women: A randomized double-blind placebo-controlled study. J. Bone Miner. Res. 2002, 17, 1904–1912. [Google Scholar]
- Filipović, B.; Šošić-Jurjević, B.; Ajdžanović, V.; Živanović, J.; Manojlović-Stojanoski, M.; Nestorović, N.; Ristić, N.; Trifunović, S.; Milošević, V. The phytoestrogen genistein prevents trabecular bone loss and affects thyroid follicular cells in a male rat model of osteoporosis. J. Anat. 2018, 233, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Marini, H.; Minutoli, L.; Polito, F.; Bitto, A.; Altavilla, D.; Atteritano, M.; Gaudio, A.; Mazzaferro, S.; Frisina, A.; Frisina, N.; et al. Effects of the phytoestrogen genistein on bone metabolism in osteopenic postmenopausal women: A randomized trial. Ann. Intern. Med. 2007, 146, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Kuboki, Y. Osteoblast-related gene expression of bone marrow cells during the osteoblastic differentiation induced by type I collagen. J. Biochem. 2001, 129, 133–138. [Google Scholar] [CrossRef]
- Jensen, E.D.; Gopalakrishnan, R.; Westendorf, J.J. Regulation of gene expression in osteoblasts. Biofactors 2010, 36, 25–32. [Google Scholar] [CrossRef]
- Jafary, F.; Hanachi, P.; Gorjipour, K. Osteoblast differentiation on collagen scaffold with immobilized alkaline phosphatase. Int. J. Organ Transplant. Med. 2017, 8, 195. [Google Scholar] [PubMed]
- Brandao-Burch, A.; Utting, J.C.; Orriss, I.R.; Arnett, T.R. Acidosis inhibits bone formation by osteoblasts in vitro by preventing mineralization. Calcif. Tissue Int. 2005, 77, 167–174. [Google Scholar] [CrossRef]
- Ling, L.; Dombrowski, C.; Foong, K.M.; Haupt, L.M.; Stein, G.S.; Nurcombe, V.; van Wijnen, A.J.; Cool, S.M. Synergism between Wnt3a and heparin enhances osteogenesis via a phosphoinositide 3-kinase/akt/RUNX2 pathway. J. Biol. Chem. 2010, 285, 26233–26244. [Google Scholar] [CrossRef]
- Stein, G.S.; Lian, J.B.; Owen, T.A. Relationship of cell growth to the regulation of tissue-specific gene expression during osteoblast differentiation. FASEB J. 1990, 4, 3111–3123. [Google Scholar] [CrossRef]
- Neve, A.; Corrado, A.; Cantatore, F.P. Osteoblast physiology in normal and pathological conditions. Cell Tissue Res. 2011, 343, 289–302. [Google Scholar] [CrossRef]
- Huang, W.; Yang, S.; Shao, J.; Li, Y. Signaling and transcriptional regulation in osteoblast commitment and differentiation. Front. Biosci. A J. Virtual Libr. 2007, 12, 3068. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Gong, Y.; Xu, L.; Zhou, M.; Li, J.; Song, J. Bidirectional regulation of osteogenic differentiation by the FOXO subfamily of forkhead transcription factors in mammalian MSCs. Cell Prolif. 2019, 52, e12540. [Google Scholar] [CrossRef] [PubMed]
- Enke, C.G.; Nagels, L.J. Undetected components in natural mixtures: How many? What concentrations? Do they account for chemical noise? What is needed to detect them? Anal. Chem. 2011, 83, 2539–2546. [Google Scholar] [CrossRef] [PubMed]
- Junio, H.A.; Sy-Cordero, A.A.; Ettefagh, K.A.; Burns, J.T.; Micko, K.T.; Graf, T.N.; Richter, S.J.; Cannon, R.E.; Oberlies, N.H.; Cech, N.B. Synergy-directed fractionation of botanical medicines: A case study with goldenseal (Hydrastis canadensis). J. Nat. Prod. 2011, 74, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Stermitz, F.R.; Scriven, L.N.; Tegos, G.; Lewis, K. Two flavonols from Artemisa annua which potentiate the activity of berberine and norfloxacin against a resistant strain of Staphylococcus aureus. Planta Med. 2002, 68, 1140–1141. [Google Scholar] [CrossRef]
- Wagner, H.; Ulrich-Merzenich, G. Synergy research: Approaching a new generation of phytopharmaceuticals. Phytomedicine 2009, 16, 97–110. [Google Scholar] [CrossRef]
- Indo, Y.; Takeshita, S.; Ishii, K.; Hoshii, T.; Aburatani, H.; Hirao, A.; Ikeda, K. Metabolic regulation of osteoclast differentiation and function. J. Bone Miner. Res. 2013, 28, 2392–2399. [Google Scholar] [CrossRef]
- Suda, T.; Takahashi, N.; Udagawa, N.; Jimi, E.; Gillespie, M.T.; Martin, T.J. Modulation of osteoclast differentiation and function by the new members of the tumor necrosis factor receptor and ligand families. Endocr. Rev. 1999, 20, 345–357. [Google Scholar] [CrossRef]
- Karsenty, G.; Wagner, E.F. Reaching a genetic and molecular understanding of skeletal development. Dev. Cell 2002, 2, 389–406. [Google Scholar] [CrossRef]
- Arai, F.; Miyamoto, T.; Ohneda, O.; Inada, T.; Sudo, T.; Brasel, K.; Miyata, T.; Anderson, D.M.; Suda, T. Commitment and differentiation of osteoclast precursor cells by the sequential expression of c-fms and receptor activator of nuclear factor κB (RANK) receptors. J. Exp. Med. 1999, 190, 1741–1754. [Google Scholar] [CrossRef]
- Teitelbaum, S.L.; Ross, F.P. Genetic regulation of osteoclast development and function. Nat. Rev. Genet. 2003, 4, 638–649. [Google Scholar] [CrossRef]
- Li, D.; Zhang, Q.; Dong, X.; Li, H.; Ma, X. Treatment with hydrogen molecules prevents RANKL-induced osteoclast differentiation associated with inhibition of ROS formation and inactivation of MAPK, AKT and NF-kappa B pathways in murine RAW264. 7 cells. J. Bone Miner. Metab. 2014, 32, 494–504. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.-E.; Yang, J.-M.; Cho, J.-H. Benincasa hispida Extract Promotes Proliferation, Differentiation, and Mineralization of MC3T3-E1 Preosteoblasts and Inhibits the Differentiation of RAW 246.7 Osteoclast Precursors. Appl. Sci. 2022, 12, 8849. https://doi.org/10.3390/app12178849
Choi Y-E, Yang J-M, Cho J-H. Benincasa hispida Extract Promotes Proliferation, Differentiation, and Mineralization of MC3T3-E1 Preosteoblasts and Inhibits the Differentiation of RAW 246.7 Osteoclast Precursors. Applied Sciences. 2022; 12(17):8849. https://doi.org/10.3390/app12178849
Chicago/Turabian StyleChoi, Ye-Eun, Jung-Mo Yang, and Ju-Hyun Cho. 2022. "Benincasa hispida Extract Promotes Proliferation, Differentiation, and Mineralization of MC3T3-E1 Preosteoblasts and Inhibits the Differentiation of RAW 246.7 Osteoclast Precursors" Applied Sciences 12, no. 17: 8849. https://doi.org/10.3390/app12178849
APA StyleChoi, Y.-E., Yang, J.-M., & Cho, J.-H. (2022). Benincasa hispida Extract Promotes Proliferation, Differentiation, and Mineralization of MC3T3-E1 Preosteoblasts and Inhibits the Differentiation of RAW 246.7 Osteoclast Precursors. Applied Sciences, 12(17), 8849. https://doi.org/10.3390/app12178849