
Overcoming Metabolic Constraints in the MEP-Pathway Enrich Salvia sclarea Hairy Roots in Therapeutic Abietane Diterpenes

 , ,
, , 
Abstract
:1. Introduction
2. Methodologies
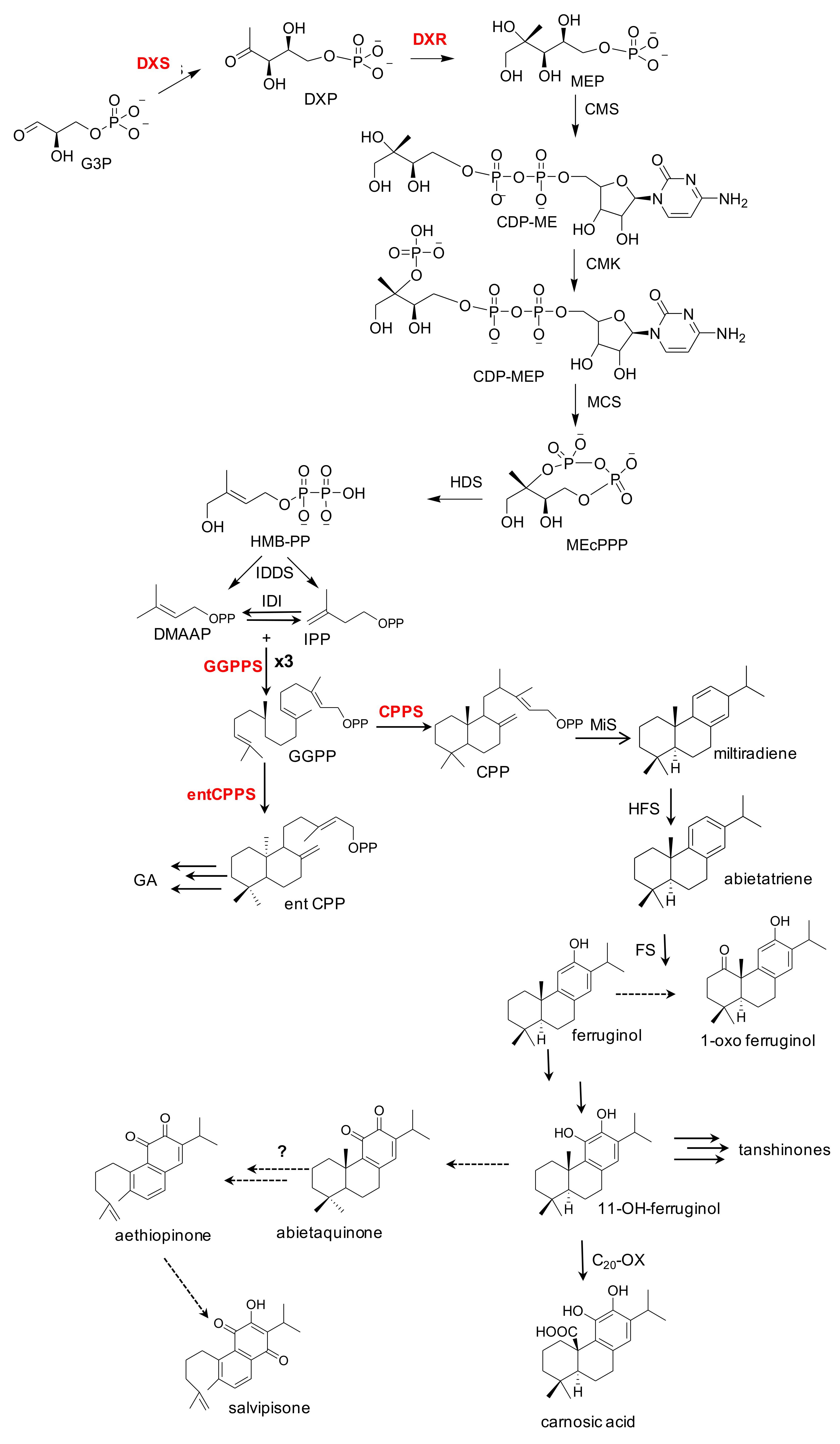
3. S. sclarea Roots Contain Bioactive Abietane Diterpenes
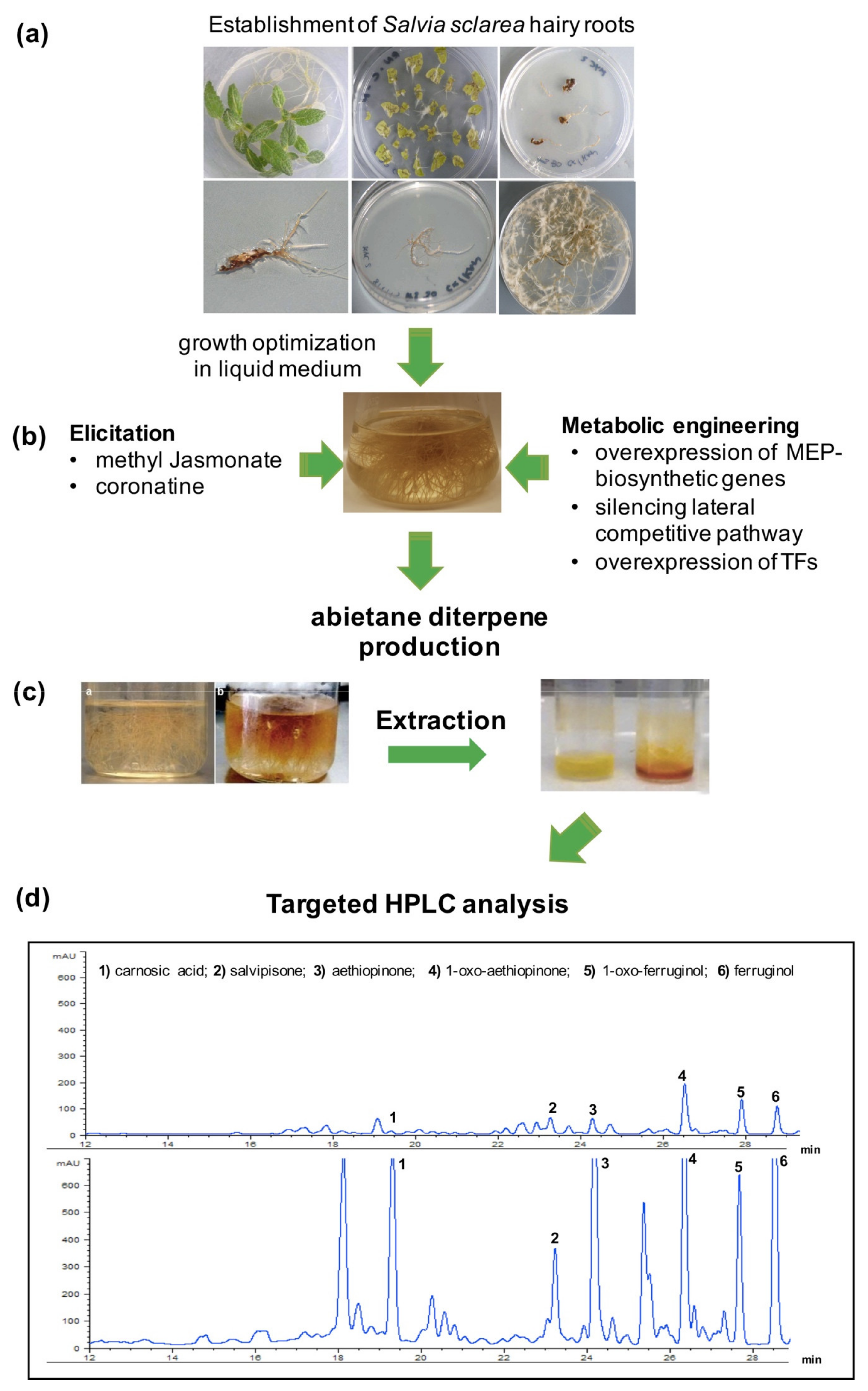
4. A Hairy Root Platform for the Production of Bioactive Abietane Diterpenes
5. Boosting the Biosynthesis of Abietane Diterpenes in S. sclarea HRs by Elicitation and Metabolic Engineering
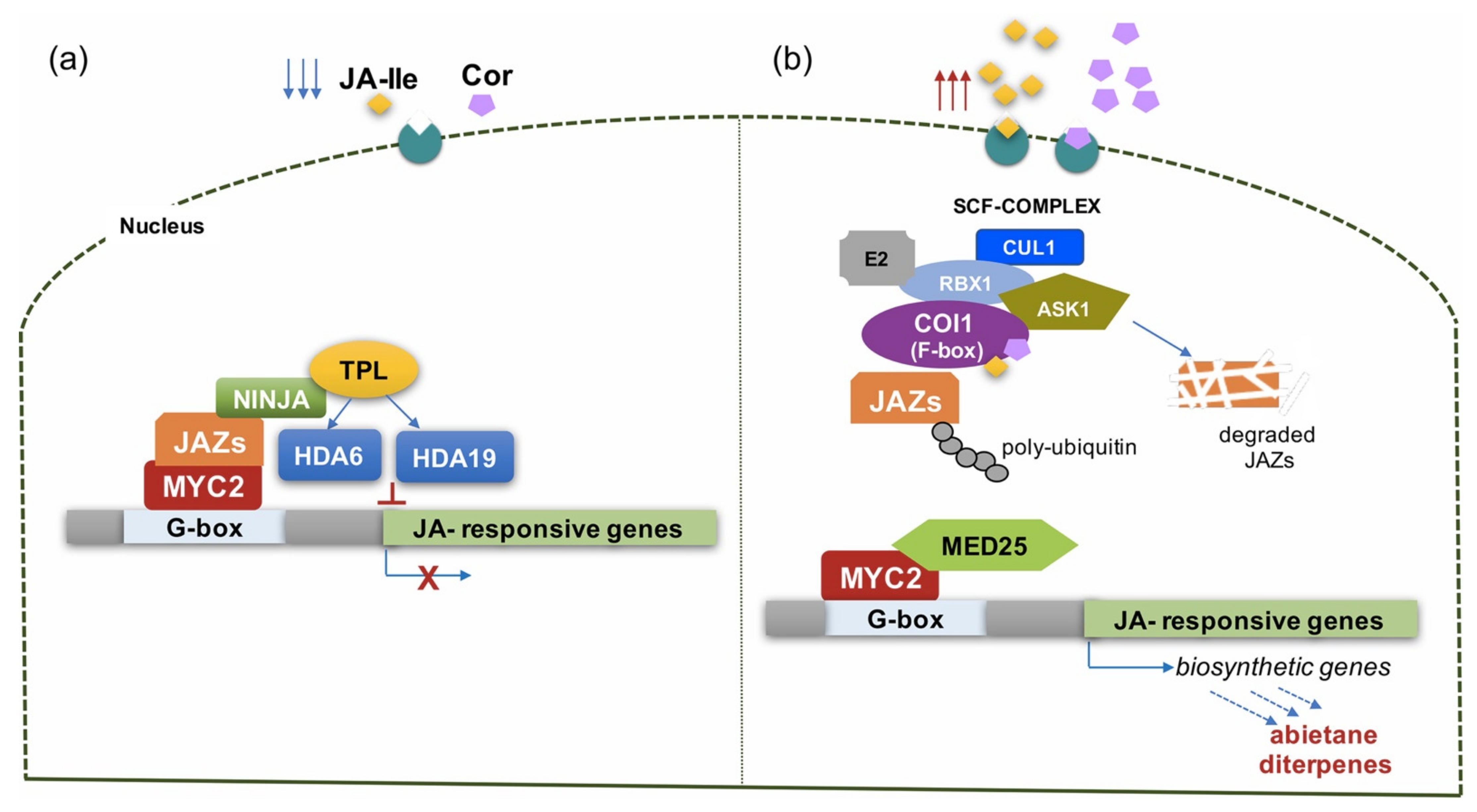
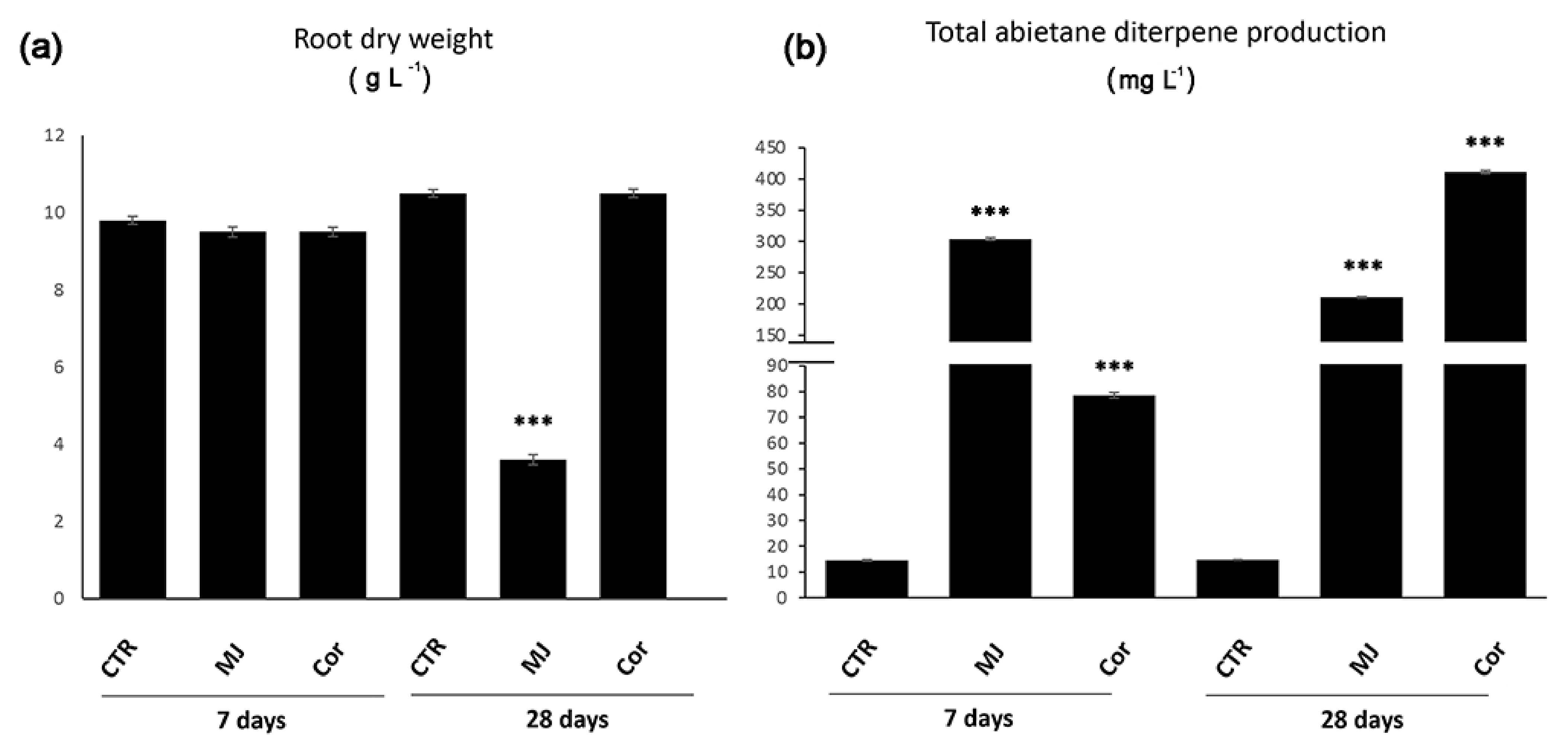
5.1. Elicitation with Methyl Jasmonate and Its Analogue Coronatine
5.1.1. MJ-Induced Accumulation of Bioactive Abietane Diterpenes in S. sclarea Is Due to the Transcriptional Regulation of Genes of the MEP-Pathway
5.1.2. Coronatine as Alternative Elicitor of the Synthesis of Abietane Diterpenes
5.2. Metabolic Engineering to Enrich the Content of Abietane Diterpenes in S. sclarea Hairy Roots
5.2.1. Overexpression of DXS and DXR Genes, encoding the First Two Enzymes Acting Up-Stream the MEP-Pathway
5.2.2. GGPP Availability Limits the Accumulation of Abietane Diterpenes in S. sclarea HRs
5.2.3. CPP Is a Precursor of Abietane Diterpenes in S. sclarea and Overexpression of SsCPPS Also Enhances Their Content
5.2.4. Co-Expression of GGPPS and CPPS Genes
5.2.5. Orchestrating the Expression of Multiple Biosynthetic Genes of the MEP-Pathway by TFs
6. Conclusions and Perspectives
- i.
- the possibility of engineering enzymes, removing catalytic constraints in key enzymes or knocking repressors, by CRISPRS-Cas9 genome editing (as reviewed in [83];
- ii.
- many secondary metabolites are synthesized/accumulated in response to a plethora of abiotic and biotic stress. Epigenetic control of plant stress response is well documented, but the role of epigenetics in regulating the secondary metabolism has so far been largely overlooked [84].
Author Contributions
Funding
Conflicts of Interest
References
- Marchev, A.S.; Yordanova, Z.P.; Georgiev, M.I. Green (cell) factories for advanced production of plant secondary metabolites. Crit. Rev. Biotechnol. 2020, 40, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Orhan, I.E.; Banach, M.; Rollinger, J.M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E.A.; et al. Natural Products in Drug Discovery: Advances and Opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Tetali, S.D. Terpenes and isoprenoids: A wealth of compounds for global use. Planta 2019, 249, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Courdavault, V.; O’Connor, S.E.; Jensen, M.K.; Papon, N. Metabolic engineering for plant natural products biosynthesis: New procedures, concrete achievements and remaining limits. Nat. Prod. Rep. 2021, 38, 2145–2153. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Liu, X.; Tian, M.; Ma, Y.; Jin, B.; Gao, W.; Cui, G.; Guo, J.; Huang, L. Recent progress and new perspectives for diterpenoid biosynthesis in medicinal plants. Med. Res. Rev. 2021, 41, 2971–2997. [Google Scholar] [CrossRef]
- Johnson, S.R.; Bhat, W.W.; Bibik, J.; Turmo, A.; Hamberger, B.; Evolutionary Mint Genomics Consortium; Hamberger, B. A database-driven approach identifies additional diterpene synthase activities in the mint family (Lamiaceae). J. Biol. Chem. 2019, 294, 1349–1362. [Google Scholar] [CrossRef] [Green Version]
- Bisio, A.; Pedrelli, F.; D’Ambola, M.; Labanca, F.; Schito, A.M.; Govaerts, R.; De Tommasi, N.; Milella, L. Quinone Diterpenes from Salvia Species: Chemistry, Botany, and Biological Activity. Phytochem. Rev. 2019, 18, 665–842. [Google Scholar] [CrossRef]
- Devappa, R.K.; Makkar, H.P.S.; Becker, K. Jatropha Toxicity—A Review. J. Toxicol. Environ. Health Part B 2010, 13, 476–507. [Google Scholar] [CrossRef]
- Vaccaro, M.; Malafronte, N.; Alfieri, M.; De Tommasi, N.; Leone, A. Enhanced biosynthesis of bioactive abietane diterpenes by overexpressing AtDXS or AtDXR genes in Salvia sclarea hairy roots. Plant Cell Tissue Organ Cult. (PCTOC) 2014, 119, 65–77. [Google Scholar] [CrossRef]
- Aiyelaagbe, O.O.; Adesogan, K.; Ekundayo, O.; Gloer, J.B. Antibacterial diterpenoids from Jatropha podagrica Hook. Phytochemistry 2007, 68, 2420–2425. [Google Scholar] [CrossRef]
- Clarkson, C.; Musonda, C.C.; Chibale, K.; Campbell, W.E.; Smith, P. Synthesis of totarol amino alcohol derivatives and their antiplasmodial activity and cytotoxicity. Bioorganic Med. Chem. 2003, 11, 4417–4422. [Google Scholar] [CrossRef]
- Sutthivaiyakit, S.; Mongkolvisut, W.; Ponsitipiboon, P.; Prabpai, S.; Kongsaeree, P.; Ruchirawat, S.; Mahidol, C. A novel 8,9-seco-rhamnofolane and a new rhamnofolane endoperoxide from Jatropha integerrima roots. Tetrahedron Lett. 2003, 44, 3637–3640. [Google Scholar] [CrossRef]
- Schmeda-Hirschmann, G.; Razmilic, I.; Sauvain, M.; Moretti, C.; Munoz, V.; Ruiz, E.; Balanza, E.; Fournet, A. Antiprotozoal Activity of Jatrogrossidione from Jatropha Grossidentata and Jatrophone from Jatropha Isabellii. Phytother. Res. 1996, 10, 375–378. [Google Scholar] [CrossRef]
- Schmeda-Hirschmann, G.; Tsichritzis, F.; Jakupovic, J. Diterpenes and a lignan from Jatropha grossidentata. Phytochemistry 1992, 31, 1731–1735. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, S. Overview of Medicinally Important Diterpenoids Derived from Plastids. Mini-Rev. Med. Chem. 2017, 17, 988–1001. [Google Scholar] [CrossRef] [PubMed]
- Caniard, A.; Zerbe, P.; Legrand, S.; Cohade, A.; Valot, N.; Magnard, J.-L.; Bohlmann, J.; Legendre, L. Discovery and Functional Characterization of Two Diterpene Synthases for Sclareol Biosynthesis in Salvia sclarea (L.) and Their Relevance for Perfume Manufacture. BMC Plant Biol. 2012, 12, 119. [Google Scholar] [CrossRef] [Green Version]
- Walencka, E.; Rozalska, S.; Wysokinska, H.; Rozalski, M.; Kuzma, L.; Rozalska, B. Salvipisone and Aethiopinone from Salvia sclarea Hairy Roots Modulate Staphylococcal Antibiotic Resistance and Express Anti-Biofilm Activity. Planta Med. 2007, 73, 545–551. [Google Scholar] [CrossRef]
- Hernández-Pérez, M.; Rabanal, R.M.; de la Torre, M.C.; Rodríguez, B. Analgesic, Anti-Inflammatory, Antipyretic and Haematological Effects of Aethiopinone, an o-Naphthoquinone Diterpenoid from Salvia Aethiopis Roots and Two Hemisynthetic Derivatives. Planta Med. 1995, 61, 505–509. [Google Scholar] [CrossRef]
- Różalski, M.; Kuźma, Ł.; Wysokińska, H.; Krajewska, U. Cytotoxic and Proapoptotic Activity of Diterpenoids from in vitro Cultivated Salvia sclarea Roots. Studies on the Leukemia Cell Lines. Z. Nat. C 2006, 61, 483–488. [Google Scholar] [CrossRef] [Green Version]
- Pereyra, C.E.; Dantas, R.F.; Ferreira, S.; Gomes, L.P.; Silva-Jr, F.P. The diverse mechanisms and anticancer potential of naphthoquinones. Cancer Cell Int. 2019, 19, 207. [Google Scholar] [CrossRef] [Green Version]
- Winder, M.; Virós, A. Mechanisms of drug resistance in melanoma BT. In Mechanisms of Drug Resistance in Cancer Therapy; Mandalà, M., Romano, E., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 91–108. ISBN 978-3-030-10507-5. [Google Scholar]
- Wu, T.; Kerbler, S.M.; Fernie, A.R.; Zhang, Y. Plant cell cultures as heterologous bio-factories for secondary metabolite production. Plant Commun. 2021, 2, 100235. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Valdes, N.; Häkkinen, S.T.; Lemasson, C.; Guillet, M.; Oksman-Caldentey, K.-M.; Ritala, A.; Cardon, F. Hairy Root Cultures—A Versatile Tool with Multiple Applications. Front. Plant Sci. 2020, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, H.H.; Firuzi, O.; Jassbi, A.R. Diterpenoids from Roots of Salvia lachnocalyx; In-silico and In-vitro Toxicity against Human Cancer Cell Lines. Iran. J. Pharm. Res. IJPR 2020, 19, 85–94. [Google Scholar] [CrossRef]
- Benrezzouk, R.; Terencio, M.; Ferrandiz, M.; Hernandez-Perez, M.; Rabanal, R.; Alcaraz, M. Inhibition of 5-lipoxygenase activity by the natural anti-inflammatory compound aethiopinone. Agents Actions 2001, 50, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T.; Trudler, D.; Oh, C.-K.; Lipton, S.A. Potential Therapeutic Use of the Rosemary Diterpene Carnosic Acid for Alzheimer’s Disease, Parkinson’s Disease, and Long-COVID through NRF2 Activation to Counteract the NLRP3 Inflammasome. Antioxidants 2022, 11, 124. [Google Scholar] [CrossRef]
- Wei, K.; Louis, H.; Emori, W.; Idante, P.S.; Agwamba, E.C.; Cheng, C.-R.; Eno, E.A.; Unimuke, T.O. Antispasmodic Activity of Carnosic Acid Extracted from Rosmarinus Officinalis: Isolation, Spectroscopic Characterization, DFT Studies, and in Sili-co Molecular Docking Investigations. J. Mol. Struct. 2022, 1260, 132795. [Google Scholar] [CrossRef]
- Irtegun Kandemir, S.; Fidan, H.S.; Yener, I.; Mete, N.; Ertas, A.; Topcu, G.; Kolak, U. Investigation of Cytotoxic and Apoptotic Effects of 63 Compounds Obtained from Salvia Species: Promising Anticancer Agents. J. Food Biochem. 2022, e14226. [Google Scholar] [CrossRef] [PubMed]
- Min, F.; Liu, X.; Li, Y.; Dong, M.; Qu, Y.; Liu, W. Carnosic Acid Suppresses the Development of Oral Squamous Cell Carcinoma via Mitochondrial-Mediated Apoptosis. Front. Oncol. 2021, 11, 760861. [Google Scholar] [CrossRef]
- Jiang, S.; Qiu, Y.; Wang, Z.; Ji, Y.; Zhang, X.; Yan, X.; Zhan, Z. Carnosic Acid Induces Antiproliferation and Anti-Metastatic Property of Esophageal Cancer Cells via MAPK Signaling Pathways. J. Oncol. 2021, 2021, 4451533. [Google Scholar] [CrossRef]
- Ossikbayeva, S.; Khanin, M.; Sharoni, Y.; Trachtenberg, A.; Tuleukhanov, S.; Sensenig, R.; Rom, S.; Danilenko, M.; Orynbayeva, Z. Curcumin and Carnosic Acid Cooperate to Inhibit Proliferation and Alter Mitochondrial Function of Metastatic Prostate Cancer Cells. Antioxidants 2021, 10, 1591. [Google Scholar] [CrossRef]
- Kolak, U.S.; Arı, Ş.; Birman, H.; Hasançebi, S.; Ulubelen, A. Cardioactive Diterpenoids from the Roots of Salvia amplexicaulis. Planta Med. 2001, 67, 761–763. [Google Scholar] [CrossRef] [PubMed]
- Ulubelen, A.; Birman, H.; Öksüz, S.; Topçu, G.; Kolak, U.; Barla, A.; Voelter, W. Cardioactive Diterpenes from the Roots of Salvia eriophora. Planta Med. 2002, 68, 818–821. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Cao, J.; Wang, X.; Zhang, Y.; Sun, Q.; Jiang, Y.; Yao, J.; Li, C.; Wang, Y.; Wang, W. Ferruginol Restores SIRT1-PGC-1α-Mediated Mitochondrial Biogenesis and Fatty Acid Oxidation for the Treatment of DOX-Induced Cardiotoxicity. Front. Pharmacol. 2021, 12, 773834. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cao, G.; Ding, D.; Li, F.; Zhao, X.; Wang, J.; Yang, Y. Ferruginol Prevents Degeneration of Dopaminergic Neurons by Enhancing Clearance of α-Synuclein in Neuronal Cells. Fitoterapia 2022, 156, 105066. [Google Scholar] [CrossRef]
- Luo, G.; Zhou, J.; Li, G.; Hu, N.; Xia, X.; Zhou, H. Ferruginol Diterpenoid Selectively Inhibits Human Thyroid Cancer Growth by Inducing Mitochondrial Dependent Apoptosis, Endogenous Reactive Oxygen Species (ROS) Production, Mitochondrial Membrane Potential Loss and Suppression of Mitogen-Activated Protein. Med. Sci. Monit. 2019, 25, 2935–2942. [Google Scholar] [CrossRef]
- Ho, S.-T.; Tung, Y.-T.; Kuo, Y.-H.; Lin, C.-C.; Wu, J.-H. Ferruginol Inhibits Non–Small Cell Lung Cancer Growth by Inducing Caspase-Associated Apoptosis. Integr. Cancer Ther. 2015, 14, 86–97. [Google Scholar] [CrossRef]
- Kuźma, L.; Bruchajzer, E.; Wysokińska, H. Diterpenoid Production in Hairy Root Culture of Salvia sclarea L. Z. Nat. C J. Biosci. 2008, 63, 621–624. [Google Scholar] [CrossRef]
- Ma, X.-H.; Ma, Y.; Tang, J.-F.; He, Y.-L.; Liu, Y.-C.; Ma, X.-J.; Shen, Y.; Cui, G.-H.; Lin, H.-X.; Rong, Q.-X.; et al. The Biosyn-thetic Pathways of Tanshinones and Phenolic Acids in Salvia Miltiorrhiza. Molecules 2015, 20, 16235–16254. [Google Scholar] [CrossRef] [Green Version]
- Vaccaro, M.C.; Alfieri, M.; De Tommasi, N.; Moses, T.; Goossens, A.; Leone, A. Boosting the Synthesis of Pharmaceutically Active Abietane Diterpenes in S. sclarea Hairy Roots by Engineering the GGPPS and CPPS Genes. Front. Plant Sci. 2020, 11, 924. [Google Scholar] [CrossRef]
- Vaccaro, M.; Bernal, V.O.; Malafronte, N.; De Tommasi, N.; Leone, A. High Yield of Bioactive Abietane Diterpenes in Salvia sclarea Hairy Roots by Overexpressing Cyanobacterial DXS or DXR Genes. Planta Med. 2019, 85, 973–980. [Google Scholar] [CrossRef] [Green Version]
- Alfieri, M.; Vaccaro, M.C.; Cappetta, E.; Ambrosone, A.; De Tommasi, N.; Leone, A. Coactivation of MEP-biosynthetic genes and accumulation of abietane diterpenes in Salvia sclarea by heterologous expression of WRKY and MYC2 transcription factors. Sci. Rep. 2018, 8, 11009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Estrada, K.; Vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.M.; Palazon, J. Elicitation, an Effective Strategy for the Biotechnological Production of Bioactive High-Added Value Compounds in Plant Cell Factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef] [PubMed]
- Halder, M.; Sarkar, S.; Jha, S. Elicitation: A biotechnological tool for enhanced production of secondary metabolites in hairy root cultures. Eng. Life Sci. 2019, 19, 880–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, X.; Jin, L.; Shimada, M.; Kim, M.G.; Mackey, D. The phytotoxin coronatine is a multifunctional component of the virulence armament of Pseudomonas syringae. Planta 2014, 240, 1149–1165. [Google Scholar] [CrossRef] [Green Version]
- Durand, A.N.; Pauwels, L.; Goossens, A. The Ubiquitin System and Jasmonate Signaling. Plants 2016, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Ho, T.-T.; Murthy, H.N.; Park, S.-Y. Methyl Jasmonate Induced Oxidative Stress and Accumulation of Secondary Metabolites in Plant Cell and Organ Cultures. Int. J. Mol. Sci. 2020, 21, 716. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Wang, H.; Yang, J.; Deng, K.; Wang, T. RNA sequencing on Amomum villosum Lour. induced by MeJA identifies the genes of WRKY and terpene synthases involved in terpene biosynthesis. Genome 2018, 61, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Hampel, D.; Mosandl, A.A.; Wüst, M. Induction of de Novo Volatile Terpene Biosynthesis via Cytosolic and Plastidial Pathways by Methyl Jasmonate in Foliage of Vitis vinifera L. J. Agric. Food Chem. 2005, 53, 2652–2657. [Google Scholar] [CrossRef]
- Martin, D.M.; Gershenzon, J.; Bohlmann, J. Induction of Volatile Terpene Biosynthesis and Diurnal Emission by Methyl Jasmonate in Foliage of Norway Spruce. Plant Physiol. 2003, 132, 1586–1599. [Google Scholar] [CrossRef] [Green Version]
- Vaccaro, M.C.; Mariaevelina, A.; Malafronte, N.; De Tommasi, N.; Leone, A. Increasing the Synthesis of Bioactive Abietane Diterpenes in Salvia sclarea Hairy Roots by Elicited Transcriptional Reprogramming. Plant Cell Rep. 2017, 36, 375–386. [Google Scholar] [CrossRef]
- Wang, Q.; Quan, S.; Xiao, H. Towards efficient terpenoid biosynthesis: Manipulating IPP and DMAPP supply. Bioresour. Bioprocess. 2019, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Shi, M.; Luo, X.; Ju, G.; Li, L.; Huang, S.; Zhang, T.; Wang, H.; Kai, G. Enhanced Diterpene Tanshinone Accumulation and Bioactivity of Transgenic Salvia miltiorrhiza Hairy Roots by Pathway Engineering. J. Agric. Food Chem. 2016, 64, 2523–2530. [Google Scholar] [CrossRef] [PubMed]
- Scheler, U.; Brandt, W.; Porzel, A.; Rothe, K.; Manzano, D.; Božić, D.; Papaefthimiou, D.; Balcke, G.U.; Henning, A.; Lohse, S.; et al. Elucidation of the biosynthesis of carnosic acid and its reconstitution in yeast. Nat. Commun. 2016, 7, 12942. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.J. Two rings in them all: The labdane-related diterpenoids. Nat. Prod. Rep. 2010, 27, 1521–1530. [Google Scholar] [CrossRef] [Green Version]
- Xing, B.; Yang, D.; Liu, L.; Han, R.; Sun, Y.; Liang, Z. Phenolic acid production is more effectively enhanced than tanshinone production by methyl jasmonate in Salvia miltiorrhiza hairy roots. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 134, 119–129. [Google Scholar] [CrossRef]
- Zhou, W.; Huang, Q.; Wu, X.; Zhou, Z.; Ding, M.; Shi, M.; Huang, F.; Li, S.; Wang, Y.; Kai, G. Comprehensive transcriptome profiling of Salvia miltiorrhiza for discovery of genes associated with the biosynthesis of tanshinones and phenolic acids. Sci. Rep. 2017, 7, 10554. [Google Scholar] [CrossRef] [Green Version]
- Akhgari, A.; Laakso, I.; Maaheimo, H.; Choi, Y.H.; Seppänen-Laakso, T.; Oksman-Caldentey, K.-M.; Rischer, H. Methyljasmonate Elicitation Increases Terpenoid Indole Alkaloid Accumulation in Rhazya stricta Hairy Root Cultures. Plants 2019, 8, 534. [Google Scholar] [CrossRef] [Green Version]
- Shoji, T.; Kajikawa, M.; Hashimoto, T. Clustered Transcription Factor Genes Regulate Nicotine Biosynthesis in Tobacco. Plant Cell 2010, 22, 3390–3409. [Google Scholar] [CrossRef] [Green Version]
- Xiang, L.; Zhu, S.; Zhao, T.; Zhang, M.; Liu, W.; Chen, M.; Lan, X.; Liao, Z. Enhancement of artemisinin content and relative expression of genes of artemisinin biosynthesis in Artemisia annua by exogenous MeJA treatment. Plant Growth Regul. 2015, 75, 435–441. [Google Scholar] [CrossRef]
- Lu, X.; Tang, K.; Li, P. Plant Metabolic Engineering Strategies for the Production of Pharmaceutical Terpenoids. Front. Plant Sci. 2016, 7, 1647. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Wang, Y.; Xing, J.; Zhang, Y.; Duan, L.; Zhang, M.; Li, Z. Coronatine Modulated the Generation of Reactive Oxygen Species for Regulating the Water Loss Rate in the Detaching Maize Seedlings. Agriculture 2021, 11, 685. [Google Scholar] [CrossRef]
- Singh, P.; Arif, Y.; Siddiqui, H.; Hayat, S. Jasmonate: A versatile messenger in plants. In Jasmonates and Salicylates Signaling in Plants; Aftab, T., Yusuf, M., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 129–158. ISBN 978-3-030-75805-9. [Google Scholar]
- Barone, R.P.; Knittel, D.K.; Ooka, J.K.; Porter, L.N.; Smith, N.T.; Owens, D.K. The production of plant natural products beneficial to humanity by metabolic engineering. Curr. Plant Biol. 2020, 24, 100121. [Google Scholar] [CrossRef]
- Tian, S.; Wang, D.; Yang, L.; Zhang, Z.; Liu, Y. A systematic review of 1-Deoxy-D-xylulose-5-phosphate synthase in terpenoid biosynthesis in plants. Plant Growth Regul. 2021, 96, 221–235. [Google Scholar] [CrossRef]
- Carretero-Paulet, L.; Cairó, A.; Botella-Pavía, P.; Besumbes, O.; Campos, N.; Boronat, A.; Rodríguez-Concepción, M. Enhanced flux through the methylerythritol 4-phosphate pathway in Arabidopsis plants overexpressing deoxyxylulose 5-phosphate reductoisomerase. Plant Mol. Biol. 2006, 62, 683–695. [Google Scholar] [CrossRef]
- Estévez, J.M.; Cantero, A.; Reindl, A.; Reichler, S.; León, P. 1-Deoxy-d-xylulose-5-phosphate Synthase, a Limiting Enzyme for Plastidic Isoprenoid Biosynthesis in Plants. J. Biol. Chem. 2001, 276, 22901–22909. [Google Scholar] [CrossRef] [Green Version]
- Kai, G.; Xu, H.; Zhou, C.; Liao, P.; Xiao, J.; Luo, X.; You, L.; Zhang, L. Metabolic engineering tanshinone biosynthetic pathway in Salvia miltiorrhiza hairy root cultures. Metab. Eng. 2011, 13, 319–327. [Google Scholar] [CrossRef]
- Peebles, C.A.; Sander, G.W.; Hughes, E.H.; Peacock, R.; Shanks, J.V.; San, K.-Y. The expression of 1-deoxy-d-xylulose synthase and geraniol-10-hydroxylase or anthranilate synthase increases terpenoid indole alkaloid accumulation in Catharanthus roseus hairy roots. Metab. Eng. 2011, 13, 234–240. [Google Scholar] [CrossRef]
- Vranová, E.; Coman, D.; Gruissem, W. Network Analysis of the MVA and MEP Pathways for Isoprenoid Synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Boya, M.T.; Valverde, S. An orthoquinone isolated from Salvia aethiopis. Phytochemistry 1981, 20, 1367–1368. [Google Scholar] [CrossRef]
- Zi, J.; Peters, R.J. Characterization of CYP76AH4 clarifies phenolic diterpenoid biosynthesis in the Lamiaceae. Org. Biomol. Chem. 2013, 11, 7650–7652. [Google Scholar] [CrossRef] [Green Version]
- Božić, D.; Papaefthimiou, D.; Brückner, K.; De Vos, R.C.H.; Tsoleridis, C.A.; Katsarou, D.; Papanikolaou, A.; Pateraki, I.; Chatzopoulou, F.M.; Dimitriadou, E.; et al. Towards Elucidating Carnosic Acid Biosynthesis in Lamiaceae: Functional Characterization of the Three First Steps of the Pathway in Salvia Fruticosa and Rosmarinus Officinalis. PLoS ONE 2015, 10, e0124106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, P.; Tong, Y.; Cheng, Q.; Hu, Y.; Zhang, M.; Yang, J.; Teng, Z.; Gao, W.; Huang, L. Functional characterization of ent-copalyl diphosphate synthase, kaurene synthase and kaurene oxidase in the Salvia miltiorrhiza gibberellin biosynthetic pathway. Sci. Rep. 2016, 6, 23057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Ni, X.; Ji, Q.; Teng, X.; Yang, Y.; Wu, C.; Zekria, D.; Zhang, D.; Kai, G. Co-overexpression of geraniol-10-hydroxylase and strictosidine synthase improves anti-cancer drug camptothecin accumulation in Ophiorrhiza pumila. Sci. Rep. 2015, 5, 08227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strader, L.; Weijers, D.; Wagner, D. Plant transcription factors—Being in the right place with the right company. Curr. Opin. Plant Biol. 2022, 65, 102136. [Google Scholar] [CrossRef]
- Wani, S.H.; Anand, S.; Singh, B.; Bohra, A.; Joshi, R. WRKY transcription factors and plant defense responses: Latest discoveries and future prospects. Plant Cell Rep. 2021, 40, 1071–1085. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Pang, S.; Lu, Z.; Jin, B. Function and Mechanism of WRKY Transcription Factors in Abiotic Stress Responses of Plants. Plants 2020, 9, 1515. [Google Scholar] [CrossRef]
- Yu, H.; Guo, W.; Yang, D.; Hou, Z.; Liang, Z. Transcriptional Profiles of SmWRKY Family Genes and Their Putative Roles in the Biosynthesis of Tanshinone and Phenolic Acids in Salvia miltiorrhiza. Int. J. Mol. Sci. 2018, 19, 1593. [Google Scholar] [CrossRef] [Green Version]
- Kazan, K.; Manners, J.M. MYC2: The Master in Action. Mol. Plant 2013, 6, 686–703. [Google Scholar] [CrossRef] [Green Version]
- Goossens, J.; Fernández-Calvo, P.; Schweizer, F.; Goossens, A. Jasmonates: Signal transduction components and their roles in environmental stress responses. Plant Mol. Biol. 2016, 91, 673–689. [Google Scholar] [CrossRef]
- De Geyter, N.; Gholami, A.; Goormachtig, S.; Goossens, A. Transcriptional machineries in jasmonate-elicited plant secondary metabolism. Trends Plant Sci. 2012, 17, 349–359. [Google Scholar] [CrossRef]
- Sabzehzari, M.; Zeinali, M.; Naghavi, M.R. CRISPR-based metabolic editing: Next-generation metabolic engineering in plants. Gene 2020, 759, 144993. [Google Scholar] [CrossRef] [PubMed]
- Selma, S.; Orzáez, D. Perspectives for epigenetic editing in crops. Transgenic Res. 2021, 30, 381–400. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diterpene | Plant Species | Biological Activity | References |
|---|---|---|---|
| aethiopinone | Salvia sclarea Salvia aethiopis Salvia lachnocalyx | Antimelanoma Antimicrobial Cytotoxic Antiinflammatory | [9] [17] [19,24] [25] |
| carnosic acid | Rosmarinus officinalis Salvia sclarea | Neuroprotective Citotoxic Antioxidant | [26,27] [28,29,30] [31] |
| ferruginol | Salvia amplexicaulis Salvia eriophora Salvia sclarea | Cardioactive Neuroprotective Cytotoxic | [32,33,34] [35] [28,36,37] |
| 1-oxo-aethiopinone | Salvia sclarea | Antimicrobial | [17,38] |
| salvipisone | Salvia sclarea | Antimicrobial Cytotoxic | [17,38] [19,28] |
| Gene | Modification/ Elicitation | Total Abietane Diterpenes mg g−1 Dry Weight | Fold- Increase | Aethiopinone Content mg g−1 Dry Weight | References |
|---|---|---|---|---|---|
| HRs control | Not transformed/not elicited | 1.2 ± 0.12 | _ | 0.5 ± 0.03 | |
| AtDXS | overexpression | ± 0.22 ** | 2.1 | 1.01 ± 0.25 * | [9] |
| AtDXR | overexpression | 3.38 ± 1.02 *** | 2.8 | 1.54 ± 0.4 ** | |
| Ssent -CPPS | RNA interference | 4.5 ± 0.50 *** | 3.7 | 1.86 ± 0.11 ** | [40] |
| SsGGPPS | overexpression | 25.64 ± 1.22 *** | 21.3 | 7.8 ± 0.71 *** | |
| SsCPPS | overexpression | 37 ± 1.60 *** | 30.8 | 10.44 ± 0.21 *** | |
| SsGGPPS / SsCPPS | overexpression | 10.24 ± 0.68 *** | 8.5 | 5.2 ± 0.12 *** | |
| cyanobacterial DXS | overexpression | 2.9 ± 0.13 ** | 2.4 | 1.21 ± 0.11 ** | [41] |
| cyanobacterial DXR | overexpression | 4.5 ± 0.24 *** | 3.75 | 2.27 ± 0.22 *** | |
| AtWRKY40 | overexpression | 4.8 ± 0.43 ** | 4.0 | 2.9 ± 0.12 ** | [42] |
| AtWRKY18 | overexpression | 2.5 ± 0.03 * | 2.1 | 0.77 ± 0.01 * | |
| AtMYC2 | overexpression | 6.5 ± 1.62 ** | 5.4 | 3.1 ± 0.10 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfieri, M.; Ambrosone, A.; Vaccaro, M.; De Tommasi, N.; Leone, A. Overcoming Metabolic Constraints in the MEP-Pathway Enrich Salvia sclarea Hairy Roots in Therapeutic Abietane Diterpenes. Appl. Sci. 2022, 12, 7116. https://doi.org/10.3390/app12147116
Alfieri M, Ambrosone A, Vaccaro M, De Tommasi N, Leone A. Overcoming Metabolic Constraints in the MEP-Pathway Enrich Salvia sclarea Hairy Roots in Therapeutic Abietane Diterpenes. Applied Sciences. 2022; 12(14):7116. https://doi.org/10.3390/app12147116
Chicago/Turabian StyleAlfieri, Mariaevelina, Alfredo Ambrosone, Mariacarmela Vaccaro, Nunziatina De Tommasi, and Antonietta Leone. 2022. "Overcoming Metabolic Constraints in the MEP-Pathway Enrich Salvia sclarea Hairy Roots in Therapeutic Abietane Diterpenes" Applied Sciences 12, no. 14: 7116. https://doi.org/10.3390/app12147116
APA StyleAlfieri, M., Ambrosone, A., Vaccaro, M., De Tommasi, N., & Leone, A. (2022). Overcoming Metabolic Constraints in the MEP-Pathway Enrich Salvia sclarea Hairy Roots in Therapeutic Abietane Diterpenes. Applied Sciences, 12(14), 7116. https://doi.org/10.3390/app12147116