
Characterization of Wild Corsican Hops and Assessment of the Performances of German Hops in Corsican Environmental Conditions through a Multidisciplinary Approach




Abstract
Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. SSR Analysis
2.3. Morphological Comparison
2.4. Essential Oils Chemical Characterization
2.5. Data Analysis
3. Results and Discussion
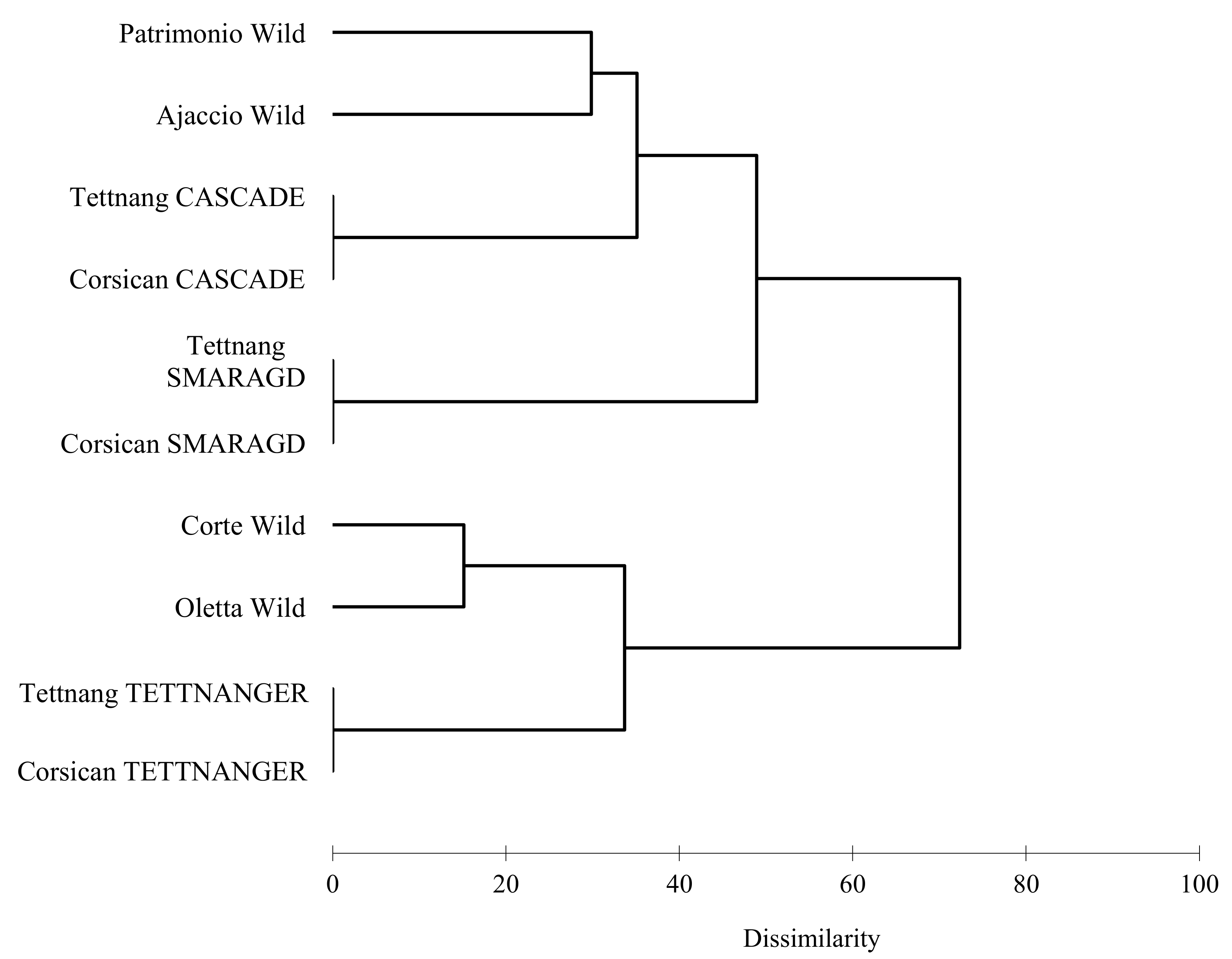
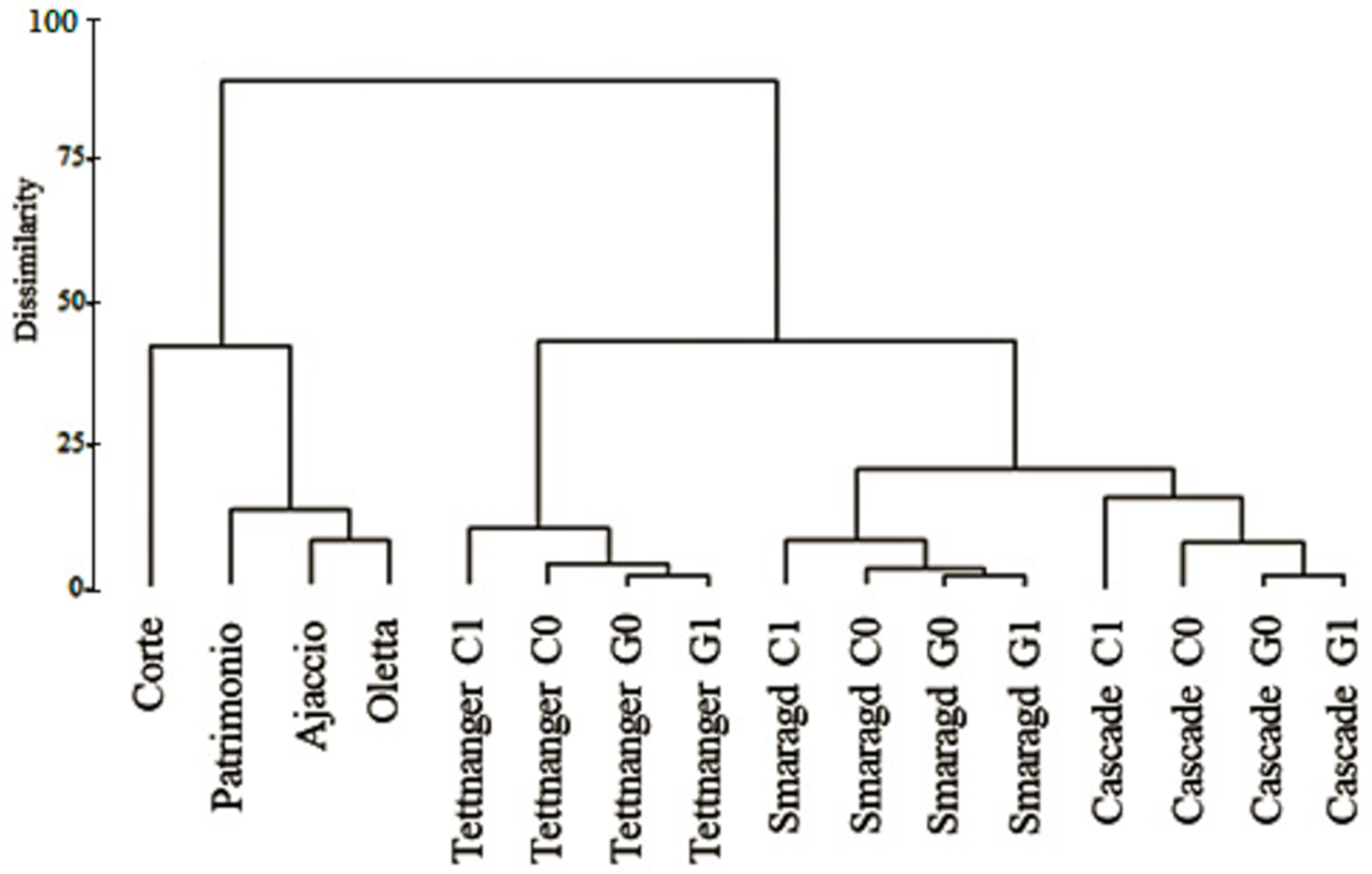
3.1. SSR Analysis
3.2. Morphological Comparison
3.3. Essential Oils Chemical Characterizations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Compounds | Cascade | Smaragd | Tettnanger | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tettnang | Corsica | Tettnang | Corsica | Tettnang | Corsica 2 | |||||||
| Year 0 | Year +1 | Year 0 | Year +1 | Year 0 | Year +1 | Year 0 | Year +1 | Year 0 | Year +1 | Year 0 | Year +1 | |
| Hydrocarbonates | ||||||||||||
| β-Pinene | 2.0 | 2.0 | 1.2 | 0.9 | 1.5 | 1.4 | 1.3 | 1.2 | 1.3 | 1.0 | 1.0 | 0.7 |
| Myrcene | 50.3 | 49.1 | 45.6 | 58.1 | 46.1 | 46.3 | 45.8 | 47.3 | 40.6 | 40.9 | 39.9 | 42.5 |
| p-Cymene | 0.3 | 0.1 | 0.2 | 0.3 | 1.1 | 1.1 | 1.1 | 1.1 | 0.7 | 0.7 | 0.8 | 0.7 |
| Limonene | 0.9 | 1.1 | 1.1 | 0.9 | 0.1 | 0.1 | 0.5 | 0.3 | 0.6 | 0.6 | 0.6 | 0.6 |
| α-Ylangene | 0.2 | 0.1 | 0.4 | 0.2 | 0.2 | 0.1 | 0.1 | 0.2 | 0.5 | 0.3 | 0.8 | 0.6 |
| α-Copaene | - | - | - | - | - | - | - | - | - | - | - | - |
| Isocaryophyllene | - | - | 0.1 | - | - | - | 0.4 | 0.2 | - | - | - | - |
| (E)-β-Caryophyllene | 4.8 | 4.3 | 7.7 | 6.4 | 6.1 | 6.0 | 7.2 | 6.5 | 5.7 | 4.7 | 6.1 | 5.5 |
| β-Copaene | - | - | 0.1 | 0.2 | 0.2 | 0.2 | 0.3 | 0.4 | 0.2 | 0.2 | - | 0.1 |
| (E)-α-Bergamotene | 0.4 | 0.7 | 0.3 | - | 0.3 | 0.2 | 0.4 | - | 0.8 | 0.9 | 0.9 | 0.7 |
| (E)-β-Farnesene | 6.8 | 6.6 | 8.2 | 3.9 | 4.1 | 4.5 | 3.9 | 4.0 | 20.2 | 20.8 | 21.2 | 19.3 |
| α-Humulene | 9.2 | 8.9 | 9.8 | 13.7 | 16.2 | 16.1 | 16.9 | 18.2 | 8.2 | 8.1 | 8.6 | 10.2 |
| 4.5-Di-epi-aristolochene | 0.1 | 0.5 | 0.1 | 0.6 | 0.1 | 0.1 | 0.2 | 0.4 | - | - | - | - |
| γ-Muurolene | 0.2 | 0.1 | 0.8 | 0.1 | 0.2 | 0.2 | 0.5 | 0.3 | 0.9 | 0.7 | 1.2 | 0.7 |
| γ-Himachalene | - | - | 0.1 | 0.1 | 0.2 | 0.4 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| β-Selinene | 1.2 | 1.3 | 1.5 | 1.2 | 5.1 | 5.9 | 4.9 | 5.1 | 0.6 | 0.7 | 0.7 | 0.5 |
| Zingiberene | - | - | 1.1 | 5.1 | - | - | 0.2 | 7.1 | - | - | 2.1 | 7.9 |
| Valencene | 1.1 | 0.8 | 2.1 | 1.2 | 0.5 | 0.3 | 0.7 | 0.5 | 0.1 | 0.1 | - | 0.1 |
| α-Selinene | 0.4 | 0.4 | 0.1 | 0.4 | 0.5 | 0.5 | 0.6 | 0.5 | 0.5 | 0.4 | 0.6 | 0.3 |
| (E.E) α-Farnesene | 0.5 | 0.4 | 0.1 | 0.1 | 0.1 | 0.3 | 0.1 | - | - | - | - | - |
| γ-Cadinene | 0.7 | 0.4 | 0.6 | 0.5 | 1.3 | 1.6 | 1.5 | 1.3 | 0.1 | 0.1 | 0.2 | 0.1 |
| δ-Cadinene | - | - | 1.2 | 0.9 | 0.1 | 0.1 | 0.9 | 0.1 | 0.1 | - | 0.1 | |
| γ-Bisabolene | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 | 0.2 | 0.1 |
| α-Cadinene | 0.3 | 0.1 | 0.4 | 0.2 | - | - | - | - | 0.2 | 0.2 | 0.4 | 0.2 |
| Alcohols | ||||||||||||
| Linalool | 1.0 | 1.4 | 0.4 | 0.3 | 1.5 | 1.4 | 1.0 | 0.5 | 1.5 | 1.2 | 1.1 | 0.4 |
| α-Terpineol | 0.1 | - | 0.1 | - | - | - | - | - | 0.1 | 0.1 | - | - |
| 8-caryolanol | 0.3 | 0.3 | 0.1 | 0.2 | 0.1 | 0.1 | - | 0.1 | 0.1 | 0.1 | 0.2 | 0.1 |
| Viridiflorol | - | - | 0.2 | - | 1.0 | 1.2 | 1.2 | 1.0 | 1.3 | 1.7 | 0.9 | 0.9 |
| Humulol | - | - | 0.1 | 0.2 | - | - | - | 0.1 | - | - | - | - |
| Zingiberenol 1 | - | - | 0.2 | 0.5 | - | - | 0.2 | 0.9 | - | - | 0.5 | 1.2 |
| Zingiberenol 2 | 0.1 | 0.1 | 0.1 | - | 0.1 | 0.1 | 0.2 | 0.1 | - | - | - | - |
| α-Cadinol | 0.4 | 0.3 | 1.7 | 0.7 | 0.1 | 0.2 | 0.1 | 0.2 | 0.2 | 0.2 | 0.3 | 0.1 |
| Eudesm-11-en-4α-ol | 0.1 | 0.2 | 1.1 | 0.1 | - | - | - | - | 0.2 | - | 0.5 | 0.1 |
| α-Bisabolol | - | - | 0.1 | - | - | - | - | - | 0.1 | 0.1 | - | - |
| Ketones | ||||||||||||
| 2-nonanone | 0.1 | 0.2 | 0.1 | - | 0.1 | 0.1 | 0.2 | 0.1 | 0.1 | 0.2 | - | - |
| 2-decanone | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.3 | 0.1 | 0.1 | 0.5 | 0.4 | 0.2 | 0.1 |
| 2-undecanone | 0.6 | 0.8 | 0.4 | 0.2 | 1.0 | 1.1 | 0.8 | 0.4 | 1.7 | 1.4 | 1.6 | 0.9 |
| 2-dodecanone | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| 2-tetradecanone | 0.2 | 0.1 | 0.1 | - | 0.3 | 0.3 | 0.5 | - | 0.1 | 0.1 | 0.2 | 0.1 |
| (Z) 2-pentadec-6-enone | - | - | - | - | 0.1 | 0.4 | - | - | 0.1 | 0.1 | - | - |
| Aldehydes | ||||||||||||
| Nonanal | 0.4 | 0.6 | 0.8 | 0.7 | 0.2 | 0.2 | 0.3 | 0.4 | - | - | - | - |
| Decanal | - | - | 0.1 | 0.1 | - | - | - | - | 0.1 | - | 0.1 | 0.1 |
| Geranial | 0.1 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 | 0.4 | 0.3 | 0.1 | 0.2 | 0.1 | 0.1 |
| Esters | ||||||||||||
| 2-methylbutyl isobutyrate | 0.3 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 | - | 0.1 | 0.1 | 0.1 | - | 0.1 |
| Methyl heptanoate | 1.8 | 1.5 | 0.1 | 0.1 | 0.3 | 0.1 | 0.4 | 0.1 | 0.1 | 0.1 | 0.2 | 0.2 |
| Methyl 6-methylheptanoate | 0.2 | 0.5 | 0.2 | 0.1 | 0.5 | 0.6 | 0.8 | 0.5 | 0.6 | 0.4 | 0.7 | 0.7 |
| Methyl octanoate | 0.1 | 0.3 | 0.2 | 0.1 | 0.7 | 0.7 | 0.5 | 0.7 | 0.1 | - | 0.1 | 0.1 |
| Methyl nonanoate | 0.1 | 0.1 | 0.1 | 0.1 | - | - | - | - | 0.2 | 0.1 | 0.5 | 0.5 |
| Methyl 4-decenoate | 0.6 | 0.8 | 0.7 | 0.6 | 1.7 | 1.9 | 0.8 | 1.4 | 0.7 | 0.9 | 0.4 | 0.7 |
| Methyl geraniate | 0.3 | 0.1 | 0.4 | 0.6 | 0.5 | 0.4 | 0.4 | 0.7 | 0.2 | 0.2 | - | 0.1 |
| Other | ||||||||||||
| Caryophyllene oxide | - | - | 0.1 | 0.1 | - | - | - | - | 0.3 | 0.5 | - | 0.3 |
| Humulene epoxide II | - | - | 0.1 | 0.1 | - | - | - | - | - | - | - | - |
| Humulene epoxide III | 0.8 | 1.0 | 0.4 | 0.1 | - | - | - | - | 0.1 | 0.1 | 0.4 | 0.5 |
| Aromadendrene oxide | 0.1 | 0.1 | 0.1 | 0.1 | - | - | - | - | - | - | - | - |
| Hexadecanoic acid | 0.2 | 0.1 | 0.2 | 0.4 | 0.1 | 0.1 | 0.1 | 0.3 | 0.2 | 0.1 | 0.3 | 0.3 |
| Essential oil rate % | 1.0 | 1.0 | 1.0 | 1.0 | 1.2 | 1.2 | 1.9 | 1.2 | 0.3 | 0.3 | 0.3 | 0.4 |
References
- Cerenak, A.; Pavlovic, M.; Luskar, M.; Kosir, I. Characterization of slovenian hop (Humulus lupulus L.) varieties by analysis of essential oil. Hop Bull. 2011, 18, 27–32. [Google Scholar]
- Howard, G.A.; Slater, C.A. Evaluation of Hops VII. Composition of the Essential Oil of Hops. J. Inst. Brew. 1957, 63, 491–506. [Google Scholar] [CrossRef]
- Katsiotis, S.T.; Langezaal, C.R.; Scheffer, J.J.C.; Verpoorte, R. Comparative study of the essential oils from hops of various Humulus lupulus L. Cultivars. Flavour Fragr. J. 1989, 4, 187–191. [Google Scholar] [CrossRef]
- Kralj, D.; Zupanec, J.; Vasilj, D.; Kralj, S.; Pšeničnik, J. Variability of Essential Oils of Hops, Humulus lupulus L. J. Inst. Brew. 1991, 97, 197–206. [Google Scholar] [CrossRef]
- Nance, M.R.; Setzer, W.N. Volatile Components of Aroma Hops (Humulus lupulus L.) Commonly Used in Beer Brewing. J. Brew. Distilling 2011, 2, 16–22. [Google Scholar]
- Shellie, R.A.; Poynter, S.D.H.; Li, J.; Gathercole, J.L.; Whittock, S.P.; Koutoulis, A. Varietal characterization of hop (Humulus lupulus L.) by GC-MS analysis of hop cone extracts. J. Sep. Sci. 2009, 32, 3720–3725. [Google Scholar] [CrossRef]
- Small, E. A Numerical and Nomenclatural Analysis of Morpho-Geographic Taxa of Humulus. Syst. Bot. 1978, 3, 37–76. [Google Scholar] [CrossRef]
- Small, E. A numerical analysis of morpho-geographic groups of cultivars of Humulus lupulus based on samples of cones. Can. J. Bot. 1981, 59, 311–324. [Google Scholar] [CrossRef]
- Stevens, J.F.; Ivancic, M.; Hsu, V.L.; Deinzer, M.L. Prenylflavonoids from Humulus lupulus. Phytochemistry 1997, 44, 1575–1585. [Google Scholar] [CrossRef]
- Stevens, J.F.; Taylor, A.W.; Nickerson, G.B.; Ivancic, M.; Henning, J.; Haunold, A.; Deinzer, M.L. Prenylflavonoid variation in Humulus lupulus: Distribution and taxonomic significance of xanthogalenol and 4′-O-methylxanthohumol. Phytochemistry 2000, 53, 759–775. [Google Scholar] [CrossRef]
- Henning, J.A.; Steiner, J.J.; Hummer, K.E. Genetic Diversity among World Hop Accessions Grown in the USA. Crop Sci. 2004, 44, 411–417. [Google Scholar] [CrossRef]
- Štěrba, K.; Čejka, P.; Čulík, J.; Jurková, M.; Krofta, K.; Pavlovič, M.; Mikyška, A.; Olšovská, J. Determination of Linalool in Different Hop Varieties Using a New Method Based on Fluidized-Bed Extraction with Gas Chromatographic-Mass Spectrometric Detection. J. Am. Soc. Brew. Chem. 2015, 73, 151–158. [Google Scholar] [CrossRef]
- Martins, Z.; Machado, J.; Cunha, S.; Barata, A.; Ferreira, I. A chemometric approach to compare Portuguese native hops with worldwide commercial varieties. J. Chemom. 2020, 34. [Google Scholar] [CrossRef]
- Green, C.P. Use of a Chromatography Data System to Identify Varieties in Binary Mixtures of Hops. J. Inst. Brew. 1997, 103, 293–296. [Google Scholar] [CrossRef]
- Pluhackova, H.; Ehrenbergerova, J.; Kretek, P.; Kocourkova, B. Hop Essential Oils in the Selected Varieties from Differently Old Hop Yards. Kvasny Prumysl 2011, 57, 266–271. [Google Scholar] [CrossRef][Green Version]
- Matsui, H.; Inui, T.; Oka, K.; Fukui, N. The influence of pruning and harvest timing on hop aroma, cone appearance, and yield. Food Chem. 2016, 202, 15–22. [Google Scholar] [CrossRef]
- Van Holle, A.; Van Landschoot, A.; Roldán-Ruiz, I.; Naudts, D.; De Keukeleire, D. The brewing value of Amarillo hops (Humulus lupulus L.) grown in northwestern USA: A preliminary study of terroir significance. J. Inst. Brew. 2017, 123, 312–318. [Google Scholar] [CrossRef]
- Morcol, T.B.; Negrin, A.; Matthews, P.D.; Kennelly, E.J. Hop (Humulus lupulus L.) terroir has large effect on a glycosylated green leaf volatile but not on other aroma glycosides. Food Chem. 2020, 321, 126644. [Google Scholar] [CrossRef]
- Almeida, A.D.R.; Maciel, M.V.D.O.B.; Gandolpho, B.C.G.; Machado, M.H.; Teixeira, G.L.; Bertoldi, F.C.; Noronha, C.M.; Vitali, L.; Block, J.M.; Barreto, P.L.M. Brazilian Grown Cascade Hop (Humulus lupulus L.): LC-ESI-MS-MS and GC-MS Analysis of Chemical Composition and Antioxidant Activity of Extracts and Essential Oils. J. Am. Soc. Brew. Chem. 2020, 1–11. [Google Scholar] [CrossRef]
- Ganino, T.; Bartolini, G.; Fabbri, A. The classification of olive germplasm. J. Hortic. Sci. Biotechnol. 2006, 81, 319–334. [Google Scholar] [CrossRef]
- Jakse, J.; Satovic, Z.; Javornik, B. Microsatellite variability among wild and cultivated hops (Humulus lupulus L.). Genome 2004, 47. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Darby, P.; Javornik, B.; Pais, M.S.S.; Seigner, E.; Lutz, A.; Svoboda, P. Microsatellite DNA Analysis of Wild Hops, Humulus lupulus L. Genet. Resour. Crop. Evol. 2006, 53. [Google Scholar] [CrossRef]
- Patzak, J.; Nesvadba, V.; Henychová, A.; Krofta, K. Assessment of the genetic diversity of wild hops (Humulus lupulus L.) in Europe using chemical and molecular analyses. Biochem. Syst. Ecol. 2010, 38, 136–145. [Google Scholar] [CrossRef]
- Mongelli, A.; Rodolfi, M.; Ganino, T.; Marieschi, M.; Dall’Asta, C.; Bruni, R. Italian hop germplasm: Characterization of wild Humulus lupulus L. genotypes from Northern Italy by means of phytochemical, morphological traits and multivariate data analysis. Ind. Crop. Prod. 2015, 70, 16–27. [Google Scholar] [CrossRef]
- Rodolfi, M.; Silvanini, A.; Chiancone, B.; Marieschi, M.; Fabbri, A.; Bruni, R.; Ganino, T. Identification and genetic structure of wild Italian Humulus lupulus L. and comparison with European and American hop cultivars using nuclear microsatellite markers. Genet. Resour. Crop. Evol. 2018, 65, 1405–1422. [Google Scholar] [CrossRef]
- Patzak, J.; Henychová, A. Evaluation of genetic variability within actual hop (Humulus lupulus L.) cultivars by an enlarged set of molecular markers. Czech J. Genet. Plant Breed. 2018, 54, 86–91. [Google Scholar] [CrossRef]
- Rodolfi, M.; Chiancone, B.; Liberatore, C.M.; Fabbri, A.; Cirlini, M.; Ganino, T. Changes in chemical profile of Cascade hop cones according to the growing area. J. Sci. Food Agric. 2019, 99, 6011–6019. [Google Scholar] [CrossRef]
- Mafakheri, M.; Kordrostami, M.; Rahimi, M.; Matthews, P.D. Evaluating genetic diversity and structure of a wild hop (Humulus lupulus L.) germplasm using morphological and molecular characteristics. Euphytica 2020, 216, 58. [Google Scholar] [CrossRef]
- Bernath, J. Acta Horticulturae; International Society for Horticultural Science: Leuven, Belgium, 2002; pp. 115–128. [Google Scholar]
- Neve, R.A. Fungal diseases. In Hops; Neve, R.A., Ed.; Springer: Dordrecht, The Netherlands, 1991; pp. 137–173. [Google Scholar]
- DeLyser, D.Y.; Kasper, W.J. Hopped Beer: The Case for Cultivation. Econ. Bot. 1994, 48, 166–170. [Google Scholar] [CrossRef]
- Van Alfen, N.K. Encyclopedia of Agriculture and Food Systems; Elsevier: Amsterdam, The Netherlands, 2004. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Stajner, N.; Jakse, J.; Kozjak, P.; Javornik, B. The isolation and characterisation of microsatellites in hop (Humulus lupulus L.). Plant Sci. 2005, 168, 213–221. [Google Scholar] [CrossRef]
- Council of Europe. European Pharmacopoeia, 3rd ed.; The Stationery Office Books: Strasbourg, France, 1997. [Google Scholar]
- Rabenold, P.P. DNA fingerprinting: An introduction. Trends Ecol. Evol. 1991, 6, 306. [Google Scholar] [CrossRef]
- Beghè, D.; Molano, J.F.G.; Fabbri, A.; Ganino, T. Olive biodiversity in Colombia: A molecular study of local germplasm. Sci. Hortic. 2015, 189, 122–131. [Google Scholar] [CrossRef]
- Ganino, T. Genetic Resources of Olea europaea L. In the Bologna Province (Italy): SSR Analysis and Identification of Local Germplasm; Firenze University Press: Florence, Italy, 2008; pp. 1000–1007. [Google Scholar]
- Belkhir, K. Genetix 4.05. Available online: https://kimura.univ-montp2.fr/genetix/ (accessed on 8 February 2021).
- Patzak, J.P.; Vrba, L.V.; Matoušek, J.M. New STS molecular markers for assessment of genetic diversity and DNA fingerprinting in hop (Humulus lupulus L.). Genome 2007, 50. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147. [Google Scholar] [CrossRef]
- Lisar, S.Y.; Motafakkerazad, R.; Hossain, M.; Rahman, I.M.M. Water Stress in Plants: Causes, Effects and Responses; IntechOpen: London, UK, 2012; pp. 1–14. [Google Scholar]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Bénévent, E. La pluviosité de la Corse. Revue Géographie Alpine 1914, 2, 239–264. [Google Scholar] [CrossRef]
- Matoušek, J.; Kocábek, T.; Patzak, J.; Bříza, J.; Siglová, K.; Mishra, A.K.; Duraisamy, G.S.; Tỳcová, A.; Ono, E.; Krofta, K. The putative role of transcription factors from HlWRKY family in regulation of the final steps of prenylflavonid and bitter acids biosynthesis in hop (Humulus lupulus L.). Plant Mol. Biol. 2016, 92, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Weihrauch, F. A new monitoring approach for the two-spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae) in hop culture. J. Plant Dis. Prot. 2004, 111, 197–205. [Google Scholar] [CrossRef]
- Yang, Y.; Luo, X.; Wei, W.; Fan, Z.; Huang, T.; Pan, X. Analysis of leaf morphology, secondary metabolites and proteins related to the resistance to Tetranychus cinnabarinus in cassava (Manihot esculenta Crantz). Sci. Rep. 2020, 10, 14197. [Google Scholar] [CrossRef]
- Polijakoff-Mayber, A. Morphological and Anatomical Changes in Plants as a Response to Salinity Stress. Plants Saline Environ. 1975, 97–117. [Google Scholar] [CrossRef]
- Joulain, D.; König, W.A. The Atlas of Spectral Data of Sesquiterpene Hydrocarbons; EB-Verlag: Hamburg, Germany, 1998. [Google Scholar]
- Patzak, J. Evaluation of genetic variability of wild hops (Humulus lupulus L.) in Canada and the Caucasus region by chemical and molecular methods. Genome 2010, 53, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Mongelli, A.; Rodolfi, M.; Ganino, T.; Marieschi, M.; Caligiani, A.; Dall’Asta, C.; Bruni, R. Are Humulus lupulus L. ecotypes and cultivars suitable for the cultivation of aromatic hop in Italy? A phytochemical approach. Ind. Crop. Prod. 2016, 83, 693–700. [Google Scholar] [CrossRef]
- Forteschi, M.; Porcu, M.C.; Fanari, M.; Zinellu, M.; Secchi, N.; Buiatti, S.; Passaghe, P.; Bertoli, S.; Pretti, L. Quality assessment of Cascade Hop (Humulus lupulus L.) grown in Sardinia. Eur. Food Res. Technol. 2019, 245, 863–871. [Google Scholar] [CrossRef]
- Maluf, W.R.; Campos, G.A.; Cardoso, M.D.G. Relationships between trichome types and spider mite (Tetranychus evansi) repellence in tomatoes with respect to foliar zingiberene contents. Euphytica 2001, 121, 73–80. [Google Scholar] [CrossRef]
- Alberts, L. Zatec, cradle of Saaz hops and commercial hop cultivation. Brew. Hist. 2020, 181, 43–50. [Google Scholar]


| Accessions/Cultivars | Geographic Coordinates | Soil Type | Altitude | Sea Proximity | Salinity |
|---|---|---|---|---|---|
| Ajaccio Wild | 41.915528 N, 8.819222 E | Clay-sand | 5 m | Very close | High |
| Oletta Wild | 42.666066 N, 9.308305 E | Clay-sand | 20 m | Very close | High |
| Patrimonio Wild | 42.697306 N, 9.361750 E | Silt-sand | 10 m | Very close | High |
| Corte Wild | 42.303591 N, 9.153470 E | Clay-sand | 410 m | Away | Weak |
| Growing Sites | Geographic Coordinates | Soil Type | Altitude | Sea Proximity | Salinity |
|---|---|---|---|---|---|
| Corsica | 42.695583 N, 9.344222 E | Silt-sand | 10 m | Very close | High |
| Tettnang | 47.668861 N, 9.604528 E | Clay-sand | 466 m | Far away | None |
| Accessions/Cultivars | H1ACA3 | H1AGA7 | H1GA23 | HIGT14 | HlGT16 | HIGT17 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SMARAGD Corsica | 211 | 223 | 187 | 212 | 243 | 249 | 165 | 167 | 233 | 233 | 195 | 195 |
| SMARAGD Tettnang | 211 | 223 | 187 | 212 | 243 | 249 | 165 | 167 | 233 | 233 | 195 | 195 |
| CASCADE Corsica | 211 | 211 | 163 | 187 | 241 | 267 | 157 | 169 | 233 | 233 | 177 | 187 |
| CASCADE Tettnang | 211 | 211 | 163 | 187 | 241 | 267 | 157 | 169 | 233 | 233 | 177 | 187 |
| TETTNANGER Corsica | 211 | 239 | 183 | 209 | 287 | 301 | 167 | 167 | 209 | 231 | 179 | 195 |
| TETTNANGER Tettnang | 211 | 239 | 183 | 209 | 287 | 301 | 167 | 167 | 209 | 231 | 179 | 195 |
| Ajaccio Wild | 230 | 236 | 167 | 187 | 243 | 265 | 161 | 167 | 229 | 233 | 179 | 187 |
| Oletta Wild | 211 | 233 | 187 | 187 | 295 | 295 | 165 | 165 | 229 | 229 | 179 | 179 |
| Patrimonio Wild | 211 | 239 | 159 | 181 | 243 | 277 | 165 | 167 | 215 | 234 | 187 | 187 |
| Corte Wild | 209 | 231 | 187 | 187 | 295 | 295 | 165 | 165 | 215 | 234 | 179 | 179 |
| Morphological Characteristics | Corte | Oletta | Patrimonio | Ajaccio |
|---|---|---|---|---|
| Main shoot: anthocyanin coloration | Strong | Absent | Absent | Absent |
| Leaf: size of blade | Medium | Large | Medium | Large |
| Leaf: blistering of upper side of blade | Absent | Absent | Absent | Absent |
| Leaf: color of upper side of blade | Green | Green | Green | Green |
| Leaf: if green, intensity of green color of the upper side of blade | Dark | Light | Dark | Dark |
| Time of flowering | Late | Late | Medium | Late |
| Plant: Growth type | Normal | Normal | Normal | Normal |
| Side shoot from the middle third of the plants: length | Long | Short | Short | Short |
| Side shoot from the upper third of the plants: Length | Long | Short | Short | Short |
| Side shoot from the middle third of the plants: density of foliage | Dense | Dense | Medium | Medium |
| Side shoot from the middle third of the plants: number of cones per node | Many | Few | Medium | Few |
| Side shoot from the middle third of the plants: total number of cones | Many | Few | Few | Few |
| Side shoot from the upper third of the plants: total number of cones | Many | Medium | Medium | Few |
| Time of picking maturity of cones | Medium | Late | Medium | Late |
| Cone: size | Large | Large | Medium | Medium |
| Cone: shape | Broad ovate | Medium ovate | Cylindric | Cylindric |
| Cone: degree of opening of bracts | Clearly open | Just open | Just open | Closed |
| Cone: intensity of green color | Light | Medium | Light | Light |
| Bract: size | Large | Small | Large | Medium |
| Bract: ratio length/width | Medium | Small | Medium | Medium |
| Bract: length of tip | Short | Short | Medium | Short |
| Morphological Characteristics | TETTNANGER | SMARAGD | CASCADE | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tettnang | Corsica | Tettnang | Corsica | Tettnang | Corsica | |||||||
| Year 0 | Year +1 | Year 0 | Year +1 | Year 0 | Year +1 | Year 0 | Year +1 | Year 0 | Year +1 | Year 0 | Year +1 | |
| Main shoot: anthocyanin coloration | Weak | Weak | Weak | Absent | Strong | Strong | Strong | Absent | Medium | Medium | Medium | Absent |
| Leaf: size of blade | Small | Small | Small | Small | Small | Small | Small | Small | Medium | Medium | Medium | Medium |
| Leaf: blistering of upper side of blade | Absent | Absent | Absent | Absent | Absent | Absent | Absent | Absent | Absent | Absent | Absent | Absent |
| Leaf: color of upper side of blade | Green | Green | Green | Green | Green | Green | Green | Green | Green | Green | Green | Green |
| Leaf: if green, intensity of green color of the upper side of blade | Dark | Dark | Dark | Dark | Dark | Dark | Dark | Dark | Dark | Dark | Dark | Dark |
| Time of flowering | Medium | Medium | Medium | Medium | Medium | Medium | Medium | Medium | Early | Early | Early | Early |
| Plant: growth type | Normal | Normal | Normal | Normal | Normal | Normal | Normal | Normal | Normal | Normal | Normal | Normal |
| Side shoot from the middle third of the plants: length | Long | Long | Long | Short | Medium | Medium | Medium | Short | Long | Long | Long | Medium |
| Side shoot from the upper third of the plants: Length | Medium | Medium | Medium | Short | Medium | Medium | Medium | Short | Long | Long | Long | Medium |
| Side shoot from the middle third of the plants: density of foliage | Dense | Dense | Dense | Dense | Sparse | Sparse | Sparse | Sparse | Medium | Medium | Medium | Medium |
| Side shoot from the middle third of the plants: number of cones per node | Few | Few | Few | Few | Few | Few | Few | Few | Medium | Medium | Medium | Medium |
| Side shoot from the middle third of the plants: total number of cones | Medium | Medium | Medium | Few | Medium | Medium | Medium | Few | Many | Many | Many | Medium |
| Side shoot from the upper third of the plants: total number of cones | Medium | Medium | Medium | Medium | Medium | Medium | Medium | Medium | Many | Many | Many | Many |
| Time of picking maturity of cones | Early | Early | Early | Early | Late | Late | Late | Late | Medium | Medium | Medium | Early |
| Cone: size | Small | Small | Small | Small | Small | Small | Small | Small | Large | Large | Large | Large |
| Cone: shape | Circular | Circular | Circular | Circular | Circular | Circular | Circular | Circular | Med. ovate | Med. ovate | Med. ovate | Med. ovate |
| Cone: degree of opening of bracts | Open | Open | Open | Open | Open | Open | Open | Open | Closed | Closed | Closed | Closed |
| Cone: intensity of green color | Light | Light | Light | Light | Dark | Dark | Dark | Dark | Medium | Medium | Medium | Medium |
| Bract: size | Small | Small | Small | Small | Small | Small | Small | Small | Large | Large | Large | Large |
| Ratio length/width | Small | Small | Small | Small | Small | Small | Small | Small | Medium | Medium | Medium | Medium |
| Length of tip | Short | Short | Short | Short | Short | Short | Short | Short | Long | Long | Long | Long |
| Compounds | LRIa | RIa | RIp | Ajaccio | Oletta | Patrimonio | Corte |
|---|---|---|---|---|---|---|---|
| Hydrocarbonates | |||||||
| β-Pinene | 964 | 970 | 1110 | 0.3 | 0.4 | 0.1 | 0.3 |
| Myrcene | 981 | 976 | 1159 | 20.1 | 19.0 | 19.1 | 6.7 |
| p-Cymene | 1010 | 1011 | 1268 | 0.1 | 0.1 | - | 0.1 |
| Limonene | 1029 | 1020 | 1199 | 0.1 | 0.2 | 0.2 | 0.1 |
| α-Ylangene | 1373 | 1375 | 1476 | 0.3 | 0.1 | 0.8 | - |
| α-Copaene | 1381 | 1379 | 1488 | - | 0.3 | 0.1 | 0.1 |
| Isocaryophyllene | 1402 | 1407 | 1571 | 0.2 | 0.1 | 0.1 | 0.5 |
| (E)-β-Caryophyllene | 1414 | 1424 | 1591 | 8.2 | 10.0 | 13.0 | 8.8 |
| β-Copaene | 1432 | 1431 | 1581 | 1.1 | 0.1 | 0.4 | 0.1 |
| (E)-α-Bergamotene | 1432 | 1432 | 1580 | 1.4 | 0.1 | 0.2 | 0.2 |
| (E)-β-Farnesene | 1445 | 1448 | 1661 | 2.0 | 2.5 | 2.8 | 0.1 |
| α-Humulene | 1455 | 1456 | 1665 | 18.3 | 15.2 | 24.9 | 5.4 |
| 4.5-Di-epi-aristolochene | 1469 | 1467 | 1665 | 1.0 | 0.5 | 0.5 | 1.2 |
| γ-Muurolene | 1477 | 1471 | 1681 | 1.7 | - | 2.9 | - |
| γ-Himachalene | 1479 | 1479 | 1693 | 0.2 | 3.6 | - | 1.4 |
| β-Selinene | 1485 | 1483 | 1712 | 6.7 | 10.0 | 2.6 | 26.8 |
| Zingiberene | 1493 | 1489 | 1717 | 11.7 | 11.6 | 5.8 | 1.6 |
| Valencene | 1495 | 1497 | 1719 | 3.1 | - | 1.5 | - |
| α-Selinene | 1505 | 1494 | - | 9.3 | 10.1 | 7.7 | 25.9 |
| (E.E) α-Farnesene | 1506 | 1498 | 1744 | 0.3 | - | 0.1 | 0.1 |
| γ-Cadinene | 1513 | 1507 | 1752 | 2.1 | 0.4 | 1.2 | 0.1 |
| δ-Cadinene | 1524 | 1516 | 1752 | 0.1 | 1.1 | 0.1 | 0.1 |
| γ-Bisabolene | 1529 | 1.9 | 2.3 | 0.2 | 0.1 | ||
| α-Cadinene | 1538 | 1535 | 1743 | 0.8 | 0.1 | 0.7 | 0.1 |
| Alcohols | |||||||
| Linalool | 1087 | 1081 | 1544 | 0.3 | 0.1 | 0.4 | 0.6 |
| α-Terpineol | 1179 | 1179 | 1700 | 0.1 | - | 0.5 | 0.1 |
| 8-caryolanol | 1562 | 1559 | 2044 | 0.1 | 0.1 | 0.2 | 0.2 |
| Viridiflorol | 1593 | 1591 | 2089 | 0.1 | - | 1.7 | - |
| Humulol | 1601 | 1588 | 2165 | 0.5 | - | 0.5 | 0.8 |
| Zingiberenol 1 | 1614 | 1599 | 2109 | 0.3 | 0.1 | 0.3 | 0.1 |
| Zingiberenol 2 | 1620 | 1613 | 2190 | 0.3 | 0.1 | 0.2 | 0.1 |
| α-Cadinol | 1642 | 1641 | 2231 | 0.7 | 1.2 | 1.2 | 2.2 |
| Eudesm-11-en-4α-ol | 1651 | 1642 | 2241 | 0.3 | 2.3 | 1.1 | 3.1 |
| α-Bisabolol | 1680 | 1672 | 2217 | 0.6 | - | 0.3 | 0.1 |
| Ketones | |||||||
| 2-nonanone | 1091 | 1070 | 1388 | 0.4 | 0.4 | - | 0.1 |
| 2-decanone | 1172 | 1176 | 1495 | 0.3 | 0.5 | - | - |
| 2-undecanone | 1291 | 1273 | 1592 | 1.1 | 2.0 | 0.8 | 0.3 |
| 2-dodecanone | 1371 | 1385 | 1711 | 0.1 | 0.3 | 0.1 | 0.2 |
| 2-tetradecanone | 1576 | 1580 | 1909 | 0.4 | 0.1 | 0.2 | 0.3 |
| (Z) 2-pentadec-6-enone | 1652 | 1647 | 0.1 | 0.8 | - | 0.8 | |
| Aldehydes | |||||||
| Nonanal | 1084 | 1083 | 1394 | 0.2 | 0.9 | 0.7 | 0.1 |
| Decanal | 1184 | 1185 | 1498 | 0.1 | 0.1 | 0.1 | - |
| Geranial | 1242 | 1244 | 1731 | 0.2 | 0.1 | 0.3 | 0.1 |
| Esters | |||||||
| 2-methylbutyl isobutyrate | 989 | 1004 | 1176 | 0.7 | 0.2 | - | 0.1 |
| Methyl heptanoate | 1006 | 1010 | - | 0.5 | 0.4 | 0.1 | |
| Methyl 6-methylheptanoate | 1070 | 1068 | 1338 | 0.3 | 0.5 | 0.6 | - |
| Methyl octanoate | 1210 | 1205 | - | 0.1 | 0.2 | 0.1 | |
| Methyl nonanoate | 1290 | 1290 | 1611 | 0.6 | 0.4 | 0.8 | 0.3 |
| Methyl 4-decenoate | - | - | - | - | |||
| Methyl geraniate | 1301 | 1301 | 1680 | 0.1 | 0.1 | 0.1 | 0.1 |
| Other | |||||||
| Caryophyllene oxide | 1571 | 1576 | 1980 | 0.1 | 0.8 | 1.2 | 0.1 |
| Humulene epoxide II | 1602 | 1601 | 2044 | 0.1 | 1.0 | 0.1 | 0.8 |
| Humulene epoxide III | 1626 | 1626 | 0.2 | 0.1 | 0.9 | 0.1 | |
| Aromadendrene oxide | 1650 | 1617 | 2002 | - | 0.8 | 1.5 | 0.1 |
| Hexadecanoic acid | 1961 | 0.2 | 0.3 | 0.2 | 0.1 | ||
| Essential oil rate % | 0.3 | 0.1 | 0.1 | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dabbous-Wach, A.; Rodolfi, M.; Paolini, J.; Costa, J.; Ganino, T. Characterization of Wild Corsican Hops and Assessment of the Performances of German Hops in Corsican Environmental Conditions through a Multidisciplinary Approach. Appl. Sci. 2021, 11, 3756. https://doi.org/10.3390/app11093756
Dabbous-Wach A, Rodolfi M, Paolini J, Costa J, Ganino T. Characterization of Wild Corsican Hops and Assessment of the Performances of German Hops in Corsican Environmental Conditions through a Multidisciplinary Approach. Applied Sciences. 2021; 11(9):3756. https://doi.org/10.3390/app11093756
Chicago/Turabian StyleDabbous-Wach, Axel, Margherita Rodolfi, Julien Paolini, Jean Costa, and Tommaso Ganino. 2021. "Characterization of Wild Corsican Hops and Assessment of the Performances of German Hops in Corsican Environmental Conditions through a Multidisciplinary Approach" Applied Sciences 11, no. 9: 3756. https://doi.org/10.3390/app11093756
APA StyleDabbous-Wach, A., Rodolfi, M., Paolini, J., Costa, J., & Ganino, T. (2021). Characterization of Wild Corsican Hops and Assessment of the Performances of German Hops in Corsican Environmental Conditions through a Multidisciplinary Approach. Applied Sciences, 11(9), 3756. https://doi.org/10.3390/app11093756