
Presence of Systemic Amyloidosis in Mice with Partial Deficiency in Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) in Aging


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Histological Analysis
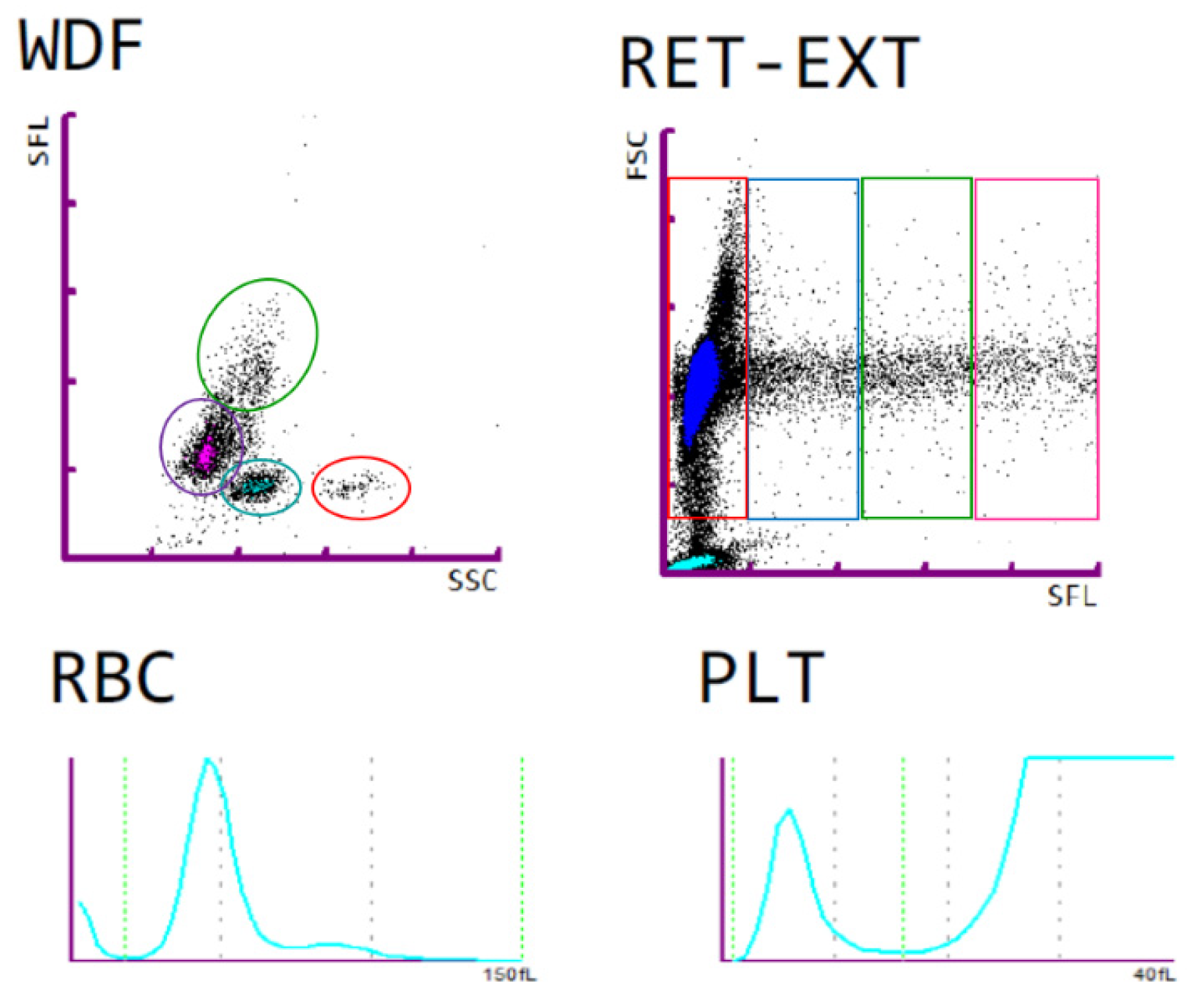
2.3. Blood Laboratory Analysis
3. Results
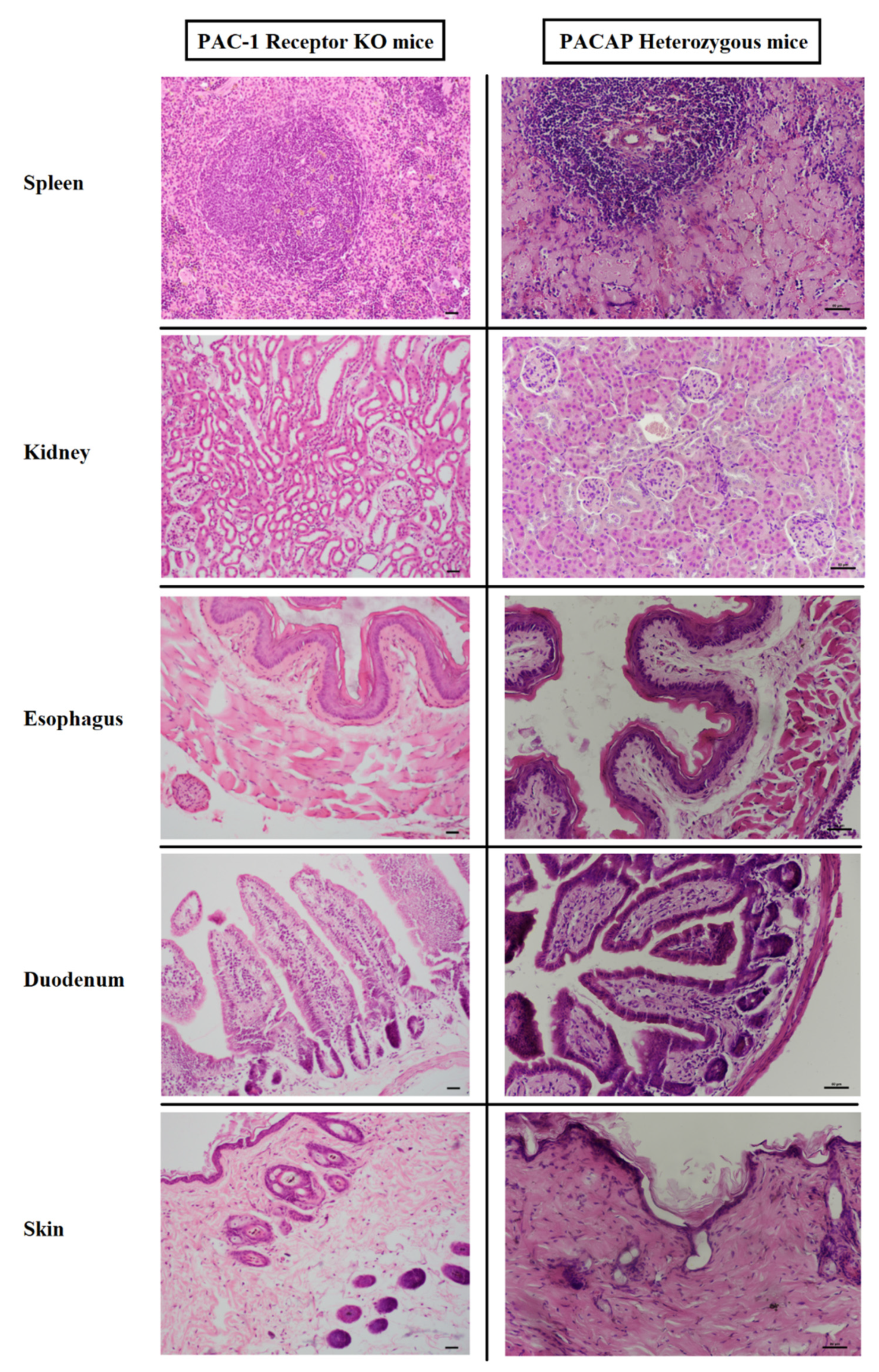
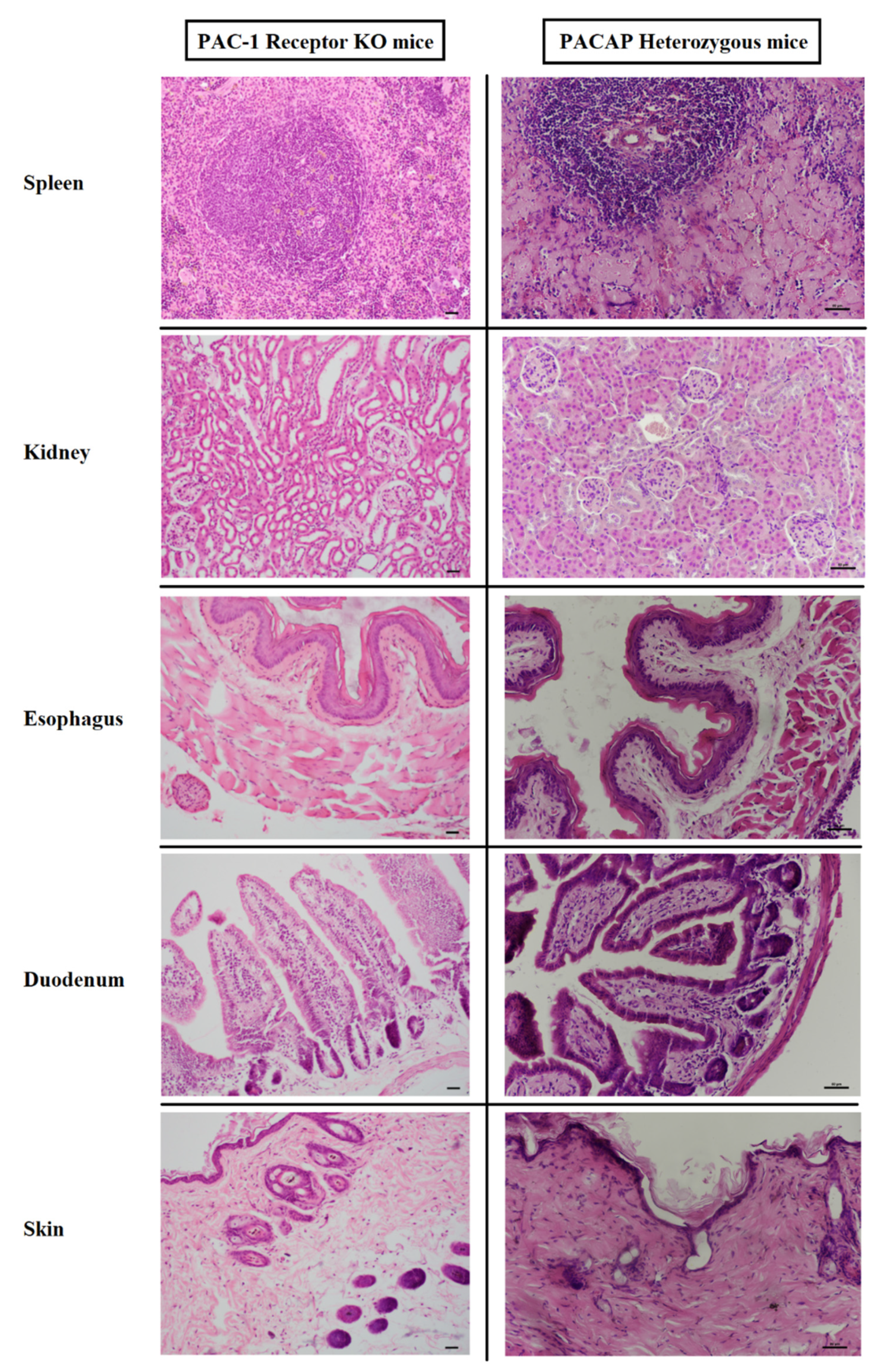
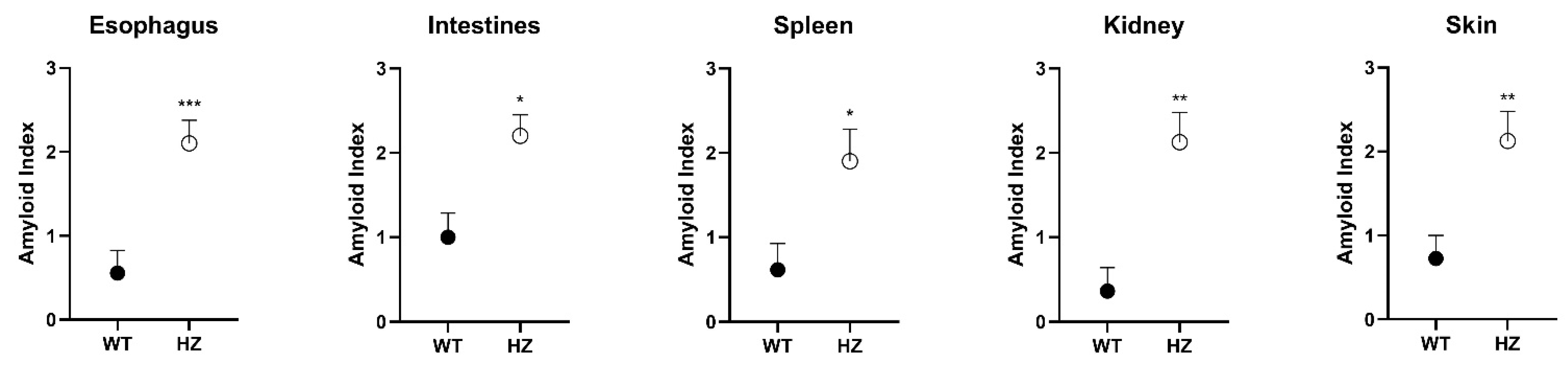
3.1. Histology
3.2. Blood Plasma Analysis and Blood Count
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.C.; Hashimoto, H.; Galas, L.; et al. Pituitary Adenylate Cyclase-Activating Polypeptide and Its Receptors: 20 Years after the Discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- Moody, T.W.; Lee, L.; Jensen, R.T. The G Protein–Coupled Receptor PAC1 Regulates Transactivation of the Receptor Tyrosine Kinase HER3. J. Mol. Neurosci. 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nicot, A.; DiCicco-Bloom, E. Regulation of neuroblast mitosis is determined by PACAP receptor isoform expression. Proc. Natl. Acad. Sci. USA 2001, 98, 4758–4763. [Google Scholar] [CrossRef] [Green Version]
- Lakk, M.; Szabo, B.; Volgyi, B.; Gabriel, R.; Denes, V. Development-related splicing regulates pituitary adenylate cyclase-activating polypeptide (PACAP) receptors in the retina. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7825–7832. [Google Scholar] [CrossRef]
- Dénes, V.; Czotter, N.; Lakk, M.; Berta, G.; Gabriel, R. PAC1-expressing structures of neural retina alter their PAC1 isoform splicing during postnatal development. Cell Tissue Res. 2014, 355, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Girard, B.M.; Campbell, S.E.; Beca, K.I.; Perkins, M.; Hsiang, H.; May, V.; Vizzard, M.A. Intrabladder PAC1 Receptor Antagonist, PACAP(6-38), Reduces Urinary Bladder Frequency and Pelvic Sensitivity in Mice Exposed to Repeated Variate Stress (RVS). J. Mol. Neurosci. 2020, 1–14. [Google Scholar] [CrossRef]
- Parsons, R.L.; May, V. PACAP-Induced PAC1 Receptor Internalization and Recruitment of Endosomal Signaling Regulate Cardiac Neuron Excitability. J. Mol. Neurosci. 2018, 68, 340–347. [Google Scholar] [CrossRef]
- Bozadjieva-Kramer, N.; Ross, R.A.; Johnson, D.Q.; Fenselau, H.; Haggerty, D.L.; Atwood, B.; Lowell, B.; Flak, J.N. The role of mediobasal hypothalamic PACAP in the control of body weight and metabolism. Endocrinology 2021, 162, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Stornetta, D.S.; Reklow, R.J.; Sahu, A.; Wabara, Y.; Nguyen, A.; Li, K.; Zhang, Y.; Perez-Reyes, E.; Ross, R.A.; et al. A brainstem peptide system activated at birth protects postnatal breathing. Nature 2021, 589, 426–430. [Google Scholar] [CrossRef]
- Körtési, T.; Tuka, B.; Tajti, J.; Bagoly, T.; Fülöp, F.; Helyes, Z.; Vécsei, L. Kynurenic acid inhibits the electrical stimulation induced elevated pituitary adenylate cyclase-activating polypeptide expression in the TNC. Front. Neurol. 2018, 8, 745. [Google Scholar] [CrossRef] [Green Version]
- Lauretta, G.; Ravalli, S.; Szychlinska, M.A.; Castorina, A.; Maugeri, G.; D’Amico, A.G.; D’Agata, V.; Musumeci, G. Current knowledge of pituitary adenylate cyclase activating polypeptide (PACAP) in articular cartilage. Histol. Histopathol. 2020, 35, 1251–1262. [Google Scholar] [CrossRef]
- Gargiulo, A.T.; Pirino, B.E.; Curtis, G.R.; Barson, J.R. Effects of pituitary adenylate cyclase-activating polypeptide isoforms in nucleus accumbens subregions on ethanol drinking. Addict. Biol. 2020, 26, e12972. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.; Musumeci, G.; Reglodi, D.; D’Agata, V. Effects of PACAP on Schwann cells: Focus on nerve injury. Int. J. Mol. Sci. 2020, 21, 8233. [Google Scholar] [CrossRef]
- Hayata-Takano, A.; Kamo, T.; Kijima, H.; Seiriki, K.; Ogata, K.; Ago, Y.; Nakazawa, T.; Shintani, Y.; Higashino, K.; Nagayasu, K.; et al. Pituitary adenylate cyclase-activating polypeptide modulates dendritic spine maturation and morphogenesis via microRNA-132 upregulation. J. Neurosci. 2019, 39, 4208–4220. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, J.; Nakamachi, T.; Matsuno, R.; Hayashi, D.; Nakamura, M.; Kikuyama, S.; Nakajo, S.; Shioda, S. Localization, characterization and function of pituitary adenylate cyclase-activating polypeptide during brain development. Peptides 2007, 28, 1713–1719. [Google Scholar] [CrossRef]
- Vincze, A.; Reglodi, D.; Helyes, Z.; Hashimoto, H.; Shintani, N.; Ábrahám, H. Role of endogenous pituitary adenylate cyclase activating polypeptide (PACAP) in myelination of the rodent brain: Lessons from PACAP-deficient mice. Int. J. Dev. Neurosci. 2011, 29, 923–935. [Google Scholar] [CrossRef]
- Raoult, E.; Bénard, M.; Komuro, H.; Lebon, A.; Vivien, D.; Fournier, A.; Vaudry, H.; Vaudry, D.; Galas, L. Cortical-layer-specific effects of PACAP and tPA on interneuron migration during post-natal development of the cerebellum. J. Neurochem. 2014, 130, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Ohtaki, H.; Watanabe, J.; Miyamoto, K.; Murai, N.; Sasaki, S.; Matsumoto, M.; Hashimoto, H.; Hiraizumi, Y.; Numazawa, S.; et al. Pituitary adenylate cyclase-activating polypeptide (PACAP) contributes to the proliferation of hematopoietic progenitor cells in murine bone marrow via PACAP-specific receptor. Sci. Rep. 2016, 6, 22373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abad, C.; Tan, Y.-V. Immunomodulatory Roles of PACAP and VIP: Lessons from knockout mice. J. Mol. Neurosci. 2018, 66, 102–113. [Google Scholar] [CrossRef]
- Juhász, T.; Helgadottir, S.L.; Tamás, A.; Reglődi, D.; Zákány, R. PACAP and VIP signaling in chondrogenesis and osteogenesis. Peptides 2015, 66, 51–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maugeri, G.; D’Amico, A.G.; Morello, G.; Reglodi, D.; Cavallaro, S.; D’Agata, V. Differential Vulnerability of Oculomotor Versus Hypoglossal Nucleus During ALS: Involvement of PACAP. Front. Neurosci. 2020, 14, 805. [Google Scholar] [CrossRef] [PubMed]
- Tóth, D.; Szabo, E.; Tamas, A.; Juhasz, T.; Horvath, G.; Fabian, E.; Opper, B.; Szabo, D.; Maugeri, G.; D’Amico, A.G.; et al. Protective Effects of PACAP in Peripheral Organs. Front. Endocrinol. 2020, 11, 377. [Google Scholar] [CrossRef] [PubMed]
- Toth, D.; Tamas, A.; Reglodi, D. The neuroprotective and biomarker potential of PACAP in human traumatic brain injury. Int. J. Mol. Sci. 2020, 21, 827. [Google Scholar] [CrossRef] [Green Version]
- Reglodi, D.; Renaud, J.; Tamas, A.; Tizabi, Y.; Socías, S.B.; Del-Bel, E.; Raisman-Vozari, R. Novel tactics for neuroprotection in Parkinson’s disease: Role of antibiotics, polyphenols and neuropeptides. Prog. Neurobiol. 2017, 155, 120–148. [Google Scholar] [CrossRef]
- Reglodi, D.; Tamas, A.; Jungling, A.; Vaczy, A.; Rivnyak, A.; Fulop, B.; Szabo, E.; Lubics, A.; Atlasz, T. Protective effects of pituitary adenylate cyclase activating polypeptide against neurotoxic agents. Neurotoxicology 2018, 66, 185–194. [Google Scholar] [CrossRef]
- Reglodi, D.; Tóth, D.; Vicena, V.; Manavalan, S.; Brown, D.; Getachew, B.; Tizabi, Y. Therapeutic potential of PACAP in alcohol toxicity. Neurochem. Int. 2019, 124, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, N.; Banks, W.A.; Shioda, S. Pituitary adenylate cyclase-activating polypeptide: Protective effects in stroke and dementia. Peptides 2020, 130, 170332. [Google Scholar] [CrossRef]
- Reglodi, D.; Jungling, A.; Longuespée, R.; Kriegsmann, J.; Casadonte, R.; Kriegsmann, M.; Juhasz, T.; Bardosi, S.; Tamas, A.; Fulop, B.D.; et al. Accelerated pre-senile systemic amyloidosis in PACAP knockout mice—A protective role of PACAP in age-related degenerative processes. J. Pathol. 2018, 245, 478–490. [Google Scholar] [CrossRef]
- Reglodi, D.; Atlasz, T.; Szabo, E.; Jungling, A.; Tamas, A.; Juhasz, T.; Fulop, B.D.; Bardosi, A. PACAP deficiency as a model of aging. GeroScience 2018, 40, 437–452. [Google Scholar] [CrossRef]
- Otto, C.; Hein, L.; Brede, M.; Jahns, R.; Engelhardt, S.; Gröne, H.-J.; Schütz, G. Pulmonary hypertension and right heart failure in pituitary adenylate cyclase-activating polypeptide type I receptor-deficient mice. Circulation 2004, 110, 3245–3251. [Google Scholar] [CrossRef] [Green Version]
- Splitthoff, P.; Rasbach, E.; Neudert, P.; Bonaterra, G.A.; Schwarz, A.; Mey, L.; Schwarzbach, H.; Eiden, L.E.; Weihe, E.; Kinscherf, R. PAC1 deficiency attenuates progression of atherosclerosis in ApoE deficient mice under cholesterol-enriched diet. Immunobiology 2020, 225, 151930. [Google Scholar] [CrossRef] [PubMed]
- Reglodi, D.; Helyes, Z.; Nemeth, J.; Vass, R.A.; Tamas, A. PACAP as a potential biomarker: Alterations of PACAP levels in human physiological and pathological conditions. In Pituitary Adenylate Cyclase Activating Polypeptide—PACAP (Current Topics in Neurotoxicity, 11); Reglodi, D., Tamas, A., Eds.; Springer: New York, NY, USA, 2016; pp. 815–832. [Google Scholar] [CrossRef] [Green Version]
- Ressler, K.J.; Mercer, K.B.; Bradley, B.; Jovanovic, T.; Mahan, A.; Kerley, K.; Norrholm, S.D.; Kilaru, V.; Smith, A.K.; Myers, A.J.; et al. Post-traumatic stress disorder is associated with PACAP and the PAC1 receptor. Nature 2011, 470, 492–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somogyvari-Vigh, A.; Reglodi, D. Pituitary adenylate cyclase activating polypeptide: A potential neuroprotective peptide. Curr. Pharm. Des. 2004, 10, 2861–2889. [Google Scholar] [CrossRef]
- D’Amico, A.G.; Maugeri, G.; Musumeci, G.; Reglodi, D.; D’Agata, V. PACAP and NAP: Effect of Two Functionally Related Peptides in Diabetic Retinopathy. J. Mol. Neurosci. 2021, 1–11. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, L.; Xie, J.; Shi, L. The emerging role of neuropeptides in Parkinson’s disease. Front. Aging Neurosci. 2021, 13, 646726. [Google Scholar] [CrossRef] [PubMed]
- Baranowska-Bik, A.; Kochanowski, J.; Uchman, D.; Wolinska-Witort, E.; Kalisz, M.; Martyńska, L.; Baranowska, B.; Bik, W. Vasoactive intestinal peptide (VIP) and pituitary adenylate cyclase activating polypeptide (PACAP) in humans with multiple sclerosis. J. Neuroimmunol. 2013, 263, 159–161. [Google Scholar] [CrossRef]
- Han, P.; Liang, W.; Baxter, L.C.; Yin, J.; Tang, Z.; Beach, T.G.; Caselli, R.J.; Reiman, E.M.; Shi, J. Pituitary adenylate cyclase-activating polypeptide is reduced in Alzheimer disease. Neurology 2014, 82, 1724–1728. [Google Scholar] [CrossRef] [Green Version]
- Han, P.; Caselli, R.J.; Baxter, L.; Serrano, G.; Yin, J.; Beach, T.G.; Reiman, E.M.; Shi, J. Association of pituitary adenylate cyclase-activating polypeptide with cognitive decline in mild cognitive impairment due to Alzheimer disease. JAMA Neurol. 2015, 72, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Ohtaki, H.; Satoh, A.; Nakamachi, T.; Yofu, S.; Dohi, K.; Mori, H.; Ohara, K.; Miyamoto, K.; Hashimoto, H.; Shintani, N.; et al. Regulation of oxidative stress by pituitary adenylate cyclase-activating polypeptide (PACAP) mediated by PACAP receptor. J. Mol. Neurosci. 2010, 42, 397–403. [Google Scholar] [CrossRef]
- Nakamachi, T.; Ohtaki, H.; Seki, T.; Yofu, S.; Kagami, N.; Hashimoto, H.; Shintani, N.; Baba, A.; Mark, L.; Lanekoff, I.; et al. PACAP suppresses dry eye signs by stimulating tear secretion. Nat. Commun. 2016, 7, 12034. [Google Scholar] [CrossRef]
- Hashimoto, H.; Shintani, N.; Tanaka, K.; Mori, W.; Hirose, M.; Matsuda, T.; Sakaue, M.; Miyazaki, J.-I.; Niwa, H.; Tashiro, F.; et al. Altered psychomotor behaviors in mice lacking pituitary adenylate cyclase-activating polypeptide (PACAP). Proc. Natl. Acad. Sci. USA 2001, 98, 13355–13360. [Google Scholar] [CrossRef] [Green Version]
- Farkas, J.; Kovács, L.; Gáspár, L.; Nafz, A.; Gaszner, T.; Ujvári, B.; Kormos, V.; Csernus, V.; Hashimoto, H.; Reglődi, D.; et al. Construct and face validity of a new model for the three-hit theory of depression using PACAP mutant mice on CD1 background. Neuroscience 2017, 354, 11–29. [Google Scholar] [CrossRef] [PubMed]
- Laird, P.W.; Zijderveld, A.; Linders, K.; Rudnicki, M.A.; Jaenisch, R.; Berns, A. Simplified mammalian DNA isolation procedure. Nucleic Acids Res. 1991, 19, 4293. [Google Scholar] [CrossRef] [Green Version]
- Jamen, F.; Persson, K.; Bertrand, G.; Rodriguez-Henche, N.; Puech, R.; Bockaert, J.; Ahrén, B.; Brabet, P. PAC1 receptor-deficient mice display impaired insulinotropic response to glucose and reduced glucose tolerance. J. Clin. Investig. 2000, 105, 1307–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May, V.; Johnson, G.C.; Hammack, S.E.; Braas, K.M.; Parsons, R.L. PAC1 Receptor Internalization and Endosomal MEK/ERK Activation Is Essential for PACAP-Mediated Neuronal Excitability. J. Mol. Neurosci. 2021, 1–7. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Castrogiovanni, P.; Saccone, S.; Federico, C.; Reibaldi, M.; Russo, A.; Bonfiglio, V.; Avitabile, T.; Longo, A.; et al. PACAP through EGFR transactivation preserves human corneal endothelial integrity. J. Cell. Biochem. 2019, 120, 10097–10105. [Google Scholar] [CrossRef]
- Ekblad, E.; Jongsma, H.; Brabet, P.; Bockaert, J.; Sundler, F. Characterization of intestinal receptors for VIP and PACAP in rat and in PAC1 receptor knockout mouse. Ann. N. Y. Acad. Sci. 2000, 921, 137–147. [Google Scholar] [CrossRef]
- Jamen, F.; Rodriguez-Henche, N.; Pralong, F.; Jegou, B.; Gaillard, R.; Bockaert, J.; Brabet, P. PAC1 null females display decreased fertility. Ann. N. Y. Acad. Sci. 2006, 921, 400–404. [Google Scholar] [CrossRef]
- Jamen, F.; Alonso, G.; Shibuya, I.; Widmer, H.; Vacher, C.-M.; Calas, A.; Bockaert, J.; Brabet, P.; Dayanithi, G. Impaired somatodendritic responses to pituitary adenylate cyclase-activating polypeptide (PACAP) of supraoptic neurones in PACAP type I -receptor deficient mice. J. Neuroendocrinol. 2003, 15, 871–881. [Google Scholar] [CrossRef]
- Barrett, K.T.; Hasan, S.U.; Scantlebury, M.H.; Wilson, R. Impaired neonatal cardiorespiratory responses to hypoxia in mice lacking PAC1 or VPAC2 receptors. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 316, R594–R606. [Google Scholar] [CrossRef] [PubMed]
- Barrett, K.T.; Hasan, S.U.; Scantlebury, M.H.; Wilson, R.J.A. Impaired cardiorespiratory responses to hypercapnia in neonatal mice lacking PAC1 but not VPAC2 receptors. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 320, R116–R128. [Google Scholar] [CrossRef]
- Sauvage, M.; Brabet, P.; Holsboer, F.; Bockaert, J.; Steckler, T. Mild deficits in mice lacking pituitary adenylate cyclase-activating polypeptide receptor type 1 (PAC1) performing on memory tasks. Mol. Brain Res. 2000, 84, 79–89. [Google Scholar] [CrossRef]
- Zink, M.; Otto, C.; Zörner, B.; Zacher, C.; Schütz, G.; Henn, F.A.; Gass, P. Reduced expression of brain-derived neurotrophic factor in mice deficient for pituitary adenylate cyclase activating polypeptide type-I-receptor. Neurosci Lett. 2004, 360, 106–108. [Google Scholar] [CrossRef] [PubMed]
- Otto, C.; Martin, M.; Wolfer, D.P.; Lipp, H.-P.; Maldonado, R.; Schütz, G. Altered emotional behavior in PACAP-type-I-receptor-deficient mice. Mol. Brain Res. 2001, 92, 78–84. [Google Scholar] [CrossRef]
- Nicot, A.; Otto, T.; Brabet, P.; DiCicco-Bloom, E.M. Altered social behavior in pituitary adenylate cyclase-activating polypeptide type I receptor-deficient mice. J. Neurosci. 2004, 24, 8786–8795. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.; Otto, C.; Santamarta, M.T.; Torrecilla, M.; Pineda, J.; Schütz, G.; Maldonado, R. Morphine withdrawal is modified in pituitary adenylate cyclase-activating polypeptide type I-receptor-deficient mice. Mol. Brain Res. 2003, 110, 109–118. [Google Scholar] [CrossRef]
- Jongsma, H.; Pettersson, L.M.E.; Zhang, Y.-Z.; Reimer, M.K.; Kanje, M.; Waldenström, A.; Sundler, F.; Danielsen, N. Markedly reduced chronic nociceptive response in mice lacking the PAC1 receptor. NeuroReport 2001, 12, 2215–2219. [Google Scholar] [CrossRef]
- Engelund, A.; Fahrenkrug, J.; Harrison, A.; Luuk, H.; Hannibal, J. Altered pupillary light reflex in PACAP receptor 1-deficient mice. Brain Res. 2012, 1453, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Georg, B.; Hannibal, J.; Fahrenkrug, J. Lack of the PAC1 receptor alters the circadian expression of VIP mRNA in the suprachiasmatic nucleus of mice. Brain Res. 2007, 1135, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Hannibal, J.; Jamen, F.; Nielsen, H.S.; Journot, L.; Brabet, P.; Fahrenkrug, J. Dissociation between light-induced phase shift of the circadian rhythm and clock gene expression in mice lacking the pituitary adenylate cyclase activating polypeptide type 1 receptor. J. Neurosci. 2001, 21, 4883–4890. [Google Scholar] [CrossRef] [Green Version]
- Hannibal, J.; Georg, B.; Fahrenkrug, J. Altered circadian food anticipatory activity rhythms in PACAP receptor 1 (PAC1) deficient mice. PLoS ONE 2016, 11, e0146981. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Matsuzaki, S.; Hattori, T.; Kuwahara, R.; Taniguchi, M.; Hashimoto, H.; Shintani, N.; Baba, A.; Kumamoto, N.; Yamada, K.; et al. Increased stathmin1 expression in the dentate gyrus of mice causes abnormal axonal arborizations. PLoS ONE 2010, 5, e8596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maasz, G.; Pirger, Z.; Reglodi, D.; Petrovics, D.; Schmidt, J.; Kiss, P.; Rivnyak, A.; Hashimoto, H.; Avar, P.; Jambor, E.; et al. Comparative protein composition of the brains of PACAP-deficient mice using mass spectrometry-based proteomic analysis. J. Mol. Neurosci. 2014, 54, 310–319. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.; Kambe, Y.; Kurihara, T.; Nakamachi, T.; Shintani, N.; Hashimoto, H.; Miyata, A. Pituitary adenylate cyclase-activating polypeptide in the ventromedial hypothalamus is responsible for food intake behavior by modulating the expression of Agouti-related peptide in mice. Mol. Neurobiol. 2020, 57, 2101–2114. [Google Scholar] [CrossRef]
- Reglodi, D.; Kiss, P.; Szabadfi, K.; Atlasz, T.; Gabriel, R.; Horvath, G.; Szakaly, P.; Sandor, B.; Lubics, A.; Laszlo, E.; et al. PACAP is an endogenous protective factor-insights from PACAP-deficient mice. J. Mol. Neurosci. 2012, 48, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Reglodi, D.; Vaczy, A.; Beltrán, A.E.R.; MaassenVanDenBrink, A. Protective effects of PACAP in ischemia. J. Headache Pain 2018, 19, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtaki, H.; Nakamachi, T.; Dohi, K.; Aizawa, Y.; Takaki, A.; Hodoyama, K.; Yofu, S.; Hashimoto, H.; Shintani, N.; Baba, A.; et al. Pituitary adenylate cyclase-activating polypeptide (PACAP) decreases ischemic neuronal cell death in association with IL-6. Proc. Natl. Acad. Sci. USA 2006, 103, 7488–7493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szakaly, P.; Horvath, G.; Kiss, P.; Láśzló, E.; Farkas, J.; Furjes, G.; Nemeth, J.; Reglodi, D. Changes in pituitary adenylate cyclase-activating polypeptide following renal ischemia-reperfusion in rats. Transplant. Proc. 2010, 42, 2283–2286. [Google Scholar] [CrossRef] [PubMed]
- Szabadfi, K.; Atlasz, T.; Kiss, P.; Danyadi, B.; Tamas, A.; Helyes, Z.; Hashimoto, H.; Shintani, N.; Baba, A.; Toth, G.; et al. Mice deficient in pituitary adenylate cyclase activating polypeptide (PACAP) are more susceptible to retinal ischemic injury in vivo. Neurotox. Res. 2012, 21, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, B.; Abad, C.; Chhith, S.; Cheung-Lau, G.; Hajji, O.; Nobuta, H.; Waschek, J. Impaired nerve regeneration and enhanced neuroinflammatory response in mice lacking pituitary adenylyl cyclase activating peptide. Neuroscience 2008, 151, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Józsa, G.; Fülöp, B.D.; Kovács, L.; Czibere, B.; Szegeczki, V.; Kiss, T.; Hajdú, T.; Tamás, A.; Helyes, Z.; Zákány, R.; et al. Lack of pituitary adenylate cyclase-activating polypeptide (PACAP) disturbs callus formation. J. Mol. Neurosci. 2019, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Endo, K.; Nakamachi, T.; Seki, T.; Kagami, N.; Wada, Y.; Nakamura, K.; Kishimoto, K.; Hori, M.; Tsuchikawa, D.; Shinntani, N.; et al. Neuroprotective effect of PACAP against NMDA-induced retinal damage in the mouse. J. Mol. Neurosci. 2011, 43, 22–29. [Google Scholar] [CrossRef]
- Schaler, A.W.; Runyan, A.M.; Clelland, C.L.; Sydney, E.J.; Fowler, S.L.; Figueroa, H.Y.; Shioda, S.; Santa-Maria, I.; Duff, K.E.; Myeku, N. PAC1 receptor-mediated clearance of tau in postsynaptic compartments attenuates tau pathology in mouse brain. Sci. Transl. Med. 2021, 13, eaba7394. [Google Scholar] [CrossRef]
- Mori, H.; Nakamachi, T.; Ohtaki, H.; Yofu, S.; Sato, A.; Endo, K.; Iso, Y.; Suzuki, H.; Takeyama, Y.; Shintani, N.; et al. Cardioprotective effect of endogenous pituitary adenylate cyclase-activating polypeptide on Doxorubicin-induced cardiomyopathy in mice. Circ. J. 2010, 74, 1183–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, S.L.; Cummings, K.J.; Jirik, F.R.; Sherwood, N.M. Targeted disruption of the pituitary adenylate cyclase-activating polypeptide gene results in early postnatal death associated with dysfunction of lipid and carbohydrate metabolism. Mol. Endocrinol. 2001, 15, 1739–1747. [Google Scholar] [CrossRef] [PubMed]
- Åkesson, L.; Ahrén, B.; Manganiello, V.C.; Holst, L.S.; Edgren, G.; Degerman, E. Dual effects of pituitary adenylate cyclase-activating polypeptide and isoproterenol on lipid metabolism and signaling in primary rat adipocytes. Endocrinology 2003, 144, 5293–5299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Åkesson, L.; Ahren, B.; Edgren, G.; Degerman, E. VPAC2-R mediates the lipolytic effects of pituitary adenylate cyclase-activating polypeptide/vasoactive intestinal polypeptide in primary rat adipocytes. Endocrinology 2005, 146, 744–750. [Google Scholar] [CrossRef] [Green Version]
- Prandota, J. Possible pathomechanisms of sudden infant death syndrome: Key role of chronic hypoxia, infection/inflammation states, cytokine irregularities, and metabolic trauma in genetically predisposed infants. Am. J. Ther. 2004, 11, 517–546. [Google Scholar] [CrossRef] [PubMed]
- Kawai, K.; Yokota, C.; Ohashi, S.; Isobe, K.; Suzuki, S.; Nakai, T.; Yamashita, K. Pituitary adenylate cyclase-activating polypeptide: Effects on pancreatic-adrenal hormone secretion and glucose-lipid metabolism in normal conscious dogs. Metabolism 1994, 43, 739–744. [Google Scholar] [CrossRef]
- Rasbach, E.; Splitthoff, P.; Bonaterra, G.A.; Schwarz, A.; Mey, L.; Schwarzbach, H.; Eiden, L.E.; Weihe, E.; Kinscherf, R. PACAP deficiency aggravates atherosclerosis in ApoE deficient mice. Immunobiology 2019, 224, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Qiu, P.; Gong, H.; Chen, X.; Sun, Y.; Hong, A.; Ma, Y. PACAP ameliorates hepatic metabolism and inflammation through up-regulating FAIM in obesity. J. Cell. Mol. Med. 2019, 23, 5970–5980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freson, K.; Peeters, K.; De Vos, R.; Wittevrongel, C.; Thys, C.; Hoylaerts, M.F.; Vermylen, J.; Van Geet, C. PACAP and its receptor VPAC1 regulate megakaryocyte maturation: Therapeutic implications. Blood 2008, 111, 1885–1893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peeters, K.; Loyen, S.; Van Kerckhoven, S.; Stoffels, K.; Hoylaerts, M.F.; Van Geet, C.; Freson, K. Thrombopoietic effect of VPAC1 inhibition during megakaryopoiesis. Br. J. Haematol. 2010, 151, 54–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PACAP WT Mice | PACAP HZ Mice | |
|---|---|---|
| Creatinin (µmol/L) | 9.50 ± 1.64 | 8.10 ± 0.68 |
| Na+ (mmol/L) | 160 ± 0.5 | 157.10 ± 0.6 |
| K+ (mmol/L) | 4.13 ± 0.09 | 4.17 ± 0.08 |
| ALP (U/L) | 51.20 ± 2.1 | 53.45 ± 2.32 |
| Cholesterol (mmol/L) | 2.21 ± 0.09 | 2.57 ± 0.08 ** |
| TG (mmol/L) | 1.06 ± 0.1 | 0.91 ± 0.07 |
| LDL (mmol/L) | 0.26 ± 0.01 | 0.37 ± 0.02 *** |
| HDL (mmol/L) | 1.86 ± 0.08 | 2.22 ± 0.07 ** |
| PACAP WT | PACAP HZ Mice | |
|---|---|---|
| White blood cell count (WBC) [109/L] | 3.56 ± 0.42 | 3.95 ± 0.66 |
| Red blood cell count (RBC) [1012/L] | 8.40 ± 0.36 | 7.91 ± 0.20 |
| Hemoglobin concentration (HGB) [g/L] | 123.91 ± 3.36 | 121.90 ± 2.21 |
| Haematocrit (HCT) [%] | 37.92 ± 1.27 | 36.31 ± 0.80 |
| Mean corpuscular volume (MCV) [fL] | 45.50 ± 1.16 | 45.91 ± 0.24 |
| Mean corpuscular hemoglobin (MCH) [pg] | 14.88 ± 0.34 | 15.45 ± 0.15 |
| Mean corpuscular hemoglobin concetration (MCHC) [g/L] | 327.64 ± 3.05 | 335.90 ± 1.66 * |
| Platelet count measured with optical method (PLT&Q) [109/L] | 1324 ± 260.25 | 1394.1 ± 66.01 |
| Red cell distribution width—standard deviation (RDW-SD) [fL] | 27.15 ± 1.86 | 23.2 ± 0.24 * |
| Red cell distribution width—coefficient of variation (RDW-CV) [%] | 18.89 ± 1.24 | 15.87 ± 0.31 * |
| Platelet distribution width (PDW) [fL] | 7.06 ± 0.15 | 6.81 ± 0.06 |
| Mean platelet volume (MPV) [fL] | 7.26 ± 0.27 | 6.93 ± 0.03 |
| Platelet large cell ratio (P-LCR) [%] | 9.62 ± 3.05 | 4.63 ± 0.27 |
| Plateletcrit (PCT) [%] | 0.75 ± 0.09 | 1.03 ± 0.06 ** |
| Nucleated red blood cell (NRBC) [109/L] | 0.06 ± 0.01 | 0.03 ± 0.01 ** |
| Neutrophil granulocyte count (NEUT) [109/L] | 0.93 ± 0.12 | 0.78 ± 0.08 |
| Lymphocyte count (LYMPH) [109/L] | 2.16 ± 0.28 | 2.63 ± 0.51 |
| Monocyte count (MONO) [109/L] | 0.30 ± 0.04 | 0.33 ± 0.06 |
| Eosinophil garnulocyte count (EO) [109/L] | 0.15 ± 0.01 | 0.17 ± 0.03 |
| Basophil granulocyte count (BASO) [109/L] | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Reticulocyte count (RET) [%] | 4.19 ± 0.19 | 3.73 ± 0.01 * |
| Immature reticulocyte fraction (IRF) [%] | 52.45 ± 1.06 | 51.26 ± 0.46 |
| Reticulocytes with low fluorescence (LFR) [%] | 47.55 ± 1.06 | 48.74 ± 0.46 |
| Reticulocytes with median fluorescence (MFR) [%] | 12.60 ± 0.38 | 11.10 ± 0.16 ** |
| Reticulocytes with high fluorescence (HFR) [%] | 39.86 ± 0.89 | 40.16 ± 0.53 |
| Reticulocyte hemoglobin equivalent (RET-HE) [pg] | 17.34 ± 0.33 | 18.45 ± 0.08 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sparks, J.; Jungling, A.; Kiss, G.; Hiripi, L.; Pham, D.; Tamas, A.; Hoffmann, O.; Bardosi, S.; Miseta, A.; Reglodi, D. Presence of Systemic Amyloidosis in Mice with Partial Deficiency in Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) in Aging. Appl. Sci. 2021, 11, 7373. https://doi.org/10.3390/app11167373
Sparks J, Jungling A, Kiss G, Hiripi L, Pham D, Tamas A, Hoffmann O, Bardosi S, Miseta A, Reglodi D. Presence of Systemic Amyloidosis in Mice with Partial Deficiency in Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) in Aging. Applied Sciences. 2021; 11(16):7373. https://doi.org/10.3390/app11167373
Chicago/Turabian StyleSparks, Jason, Adel Jungling, Gabriella Kiss, Laszlo Hiripi, Daniel Pham, Andrea Tamas, Orsolya Hoffmann, Sebastian Bardosi, Attila Miseta, and Dora Reglodi. 2021. "Presence of Systemic Amyloidosis in Mice with Partial Deficiency in Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) in Aging" Applied Sciences 11, no. 16: 7373. https://doi.org/10.3390/app11167373
APA StyleSparks, J., Jungling, A., Kiss, G., Hiripi, L., Pham, D., Tamas, A., Hoffmann, O., Bardosi, S., Miseta, A., & Reglodi, D. (2021). Presence of Systemic Amyloidosis in Mice with Partial Deficiency in Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) in Aging. Applied Sciences, 11(16), 7373. https://doi.org/10.3390/app11167373