Physical, Chemical, and Genetic Techniques for Diatom Frustule Modification: Applications in Nanotechnology



Abstract
1. Introduction
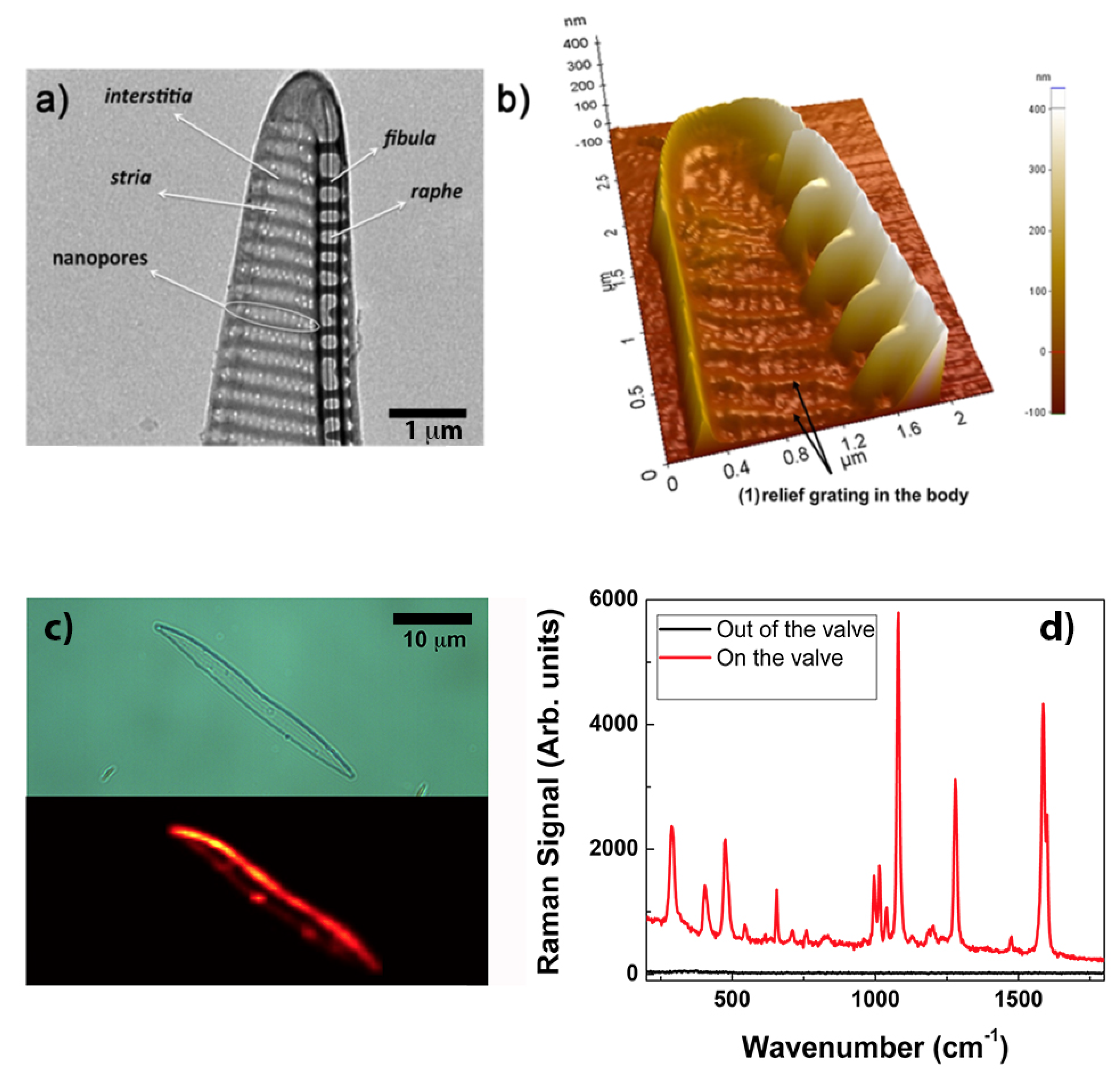
2. The Anatomy of the Frustule
3. Frustule Modification: Techniques and Applications
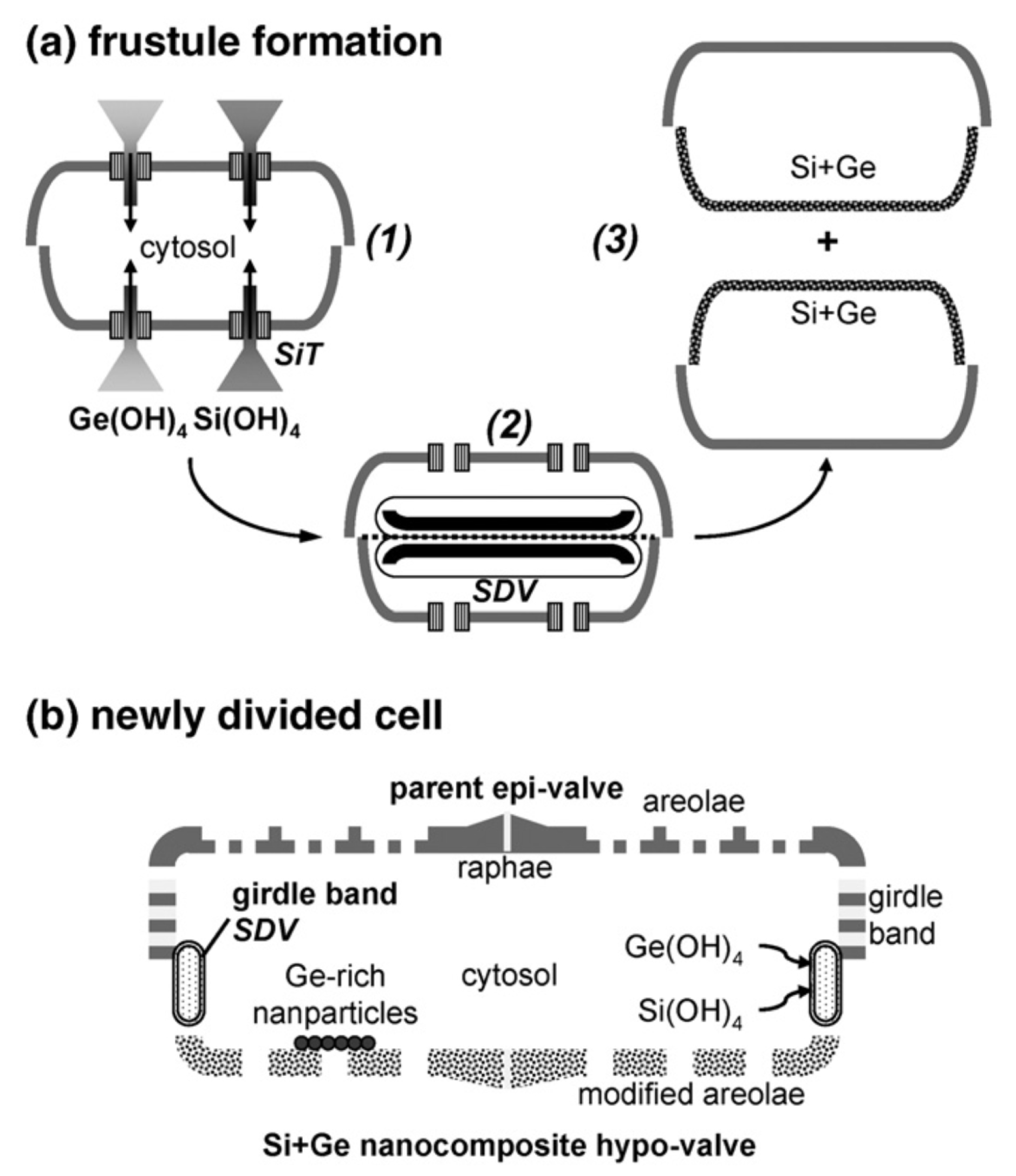
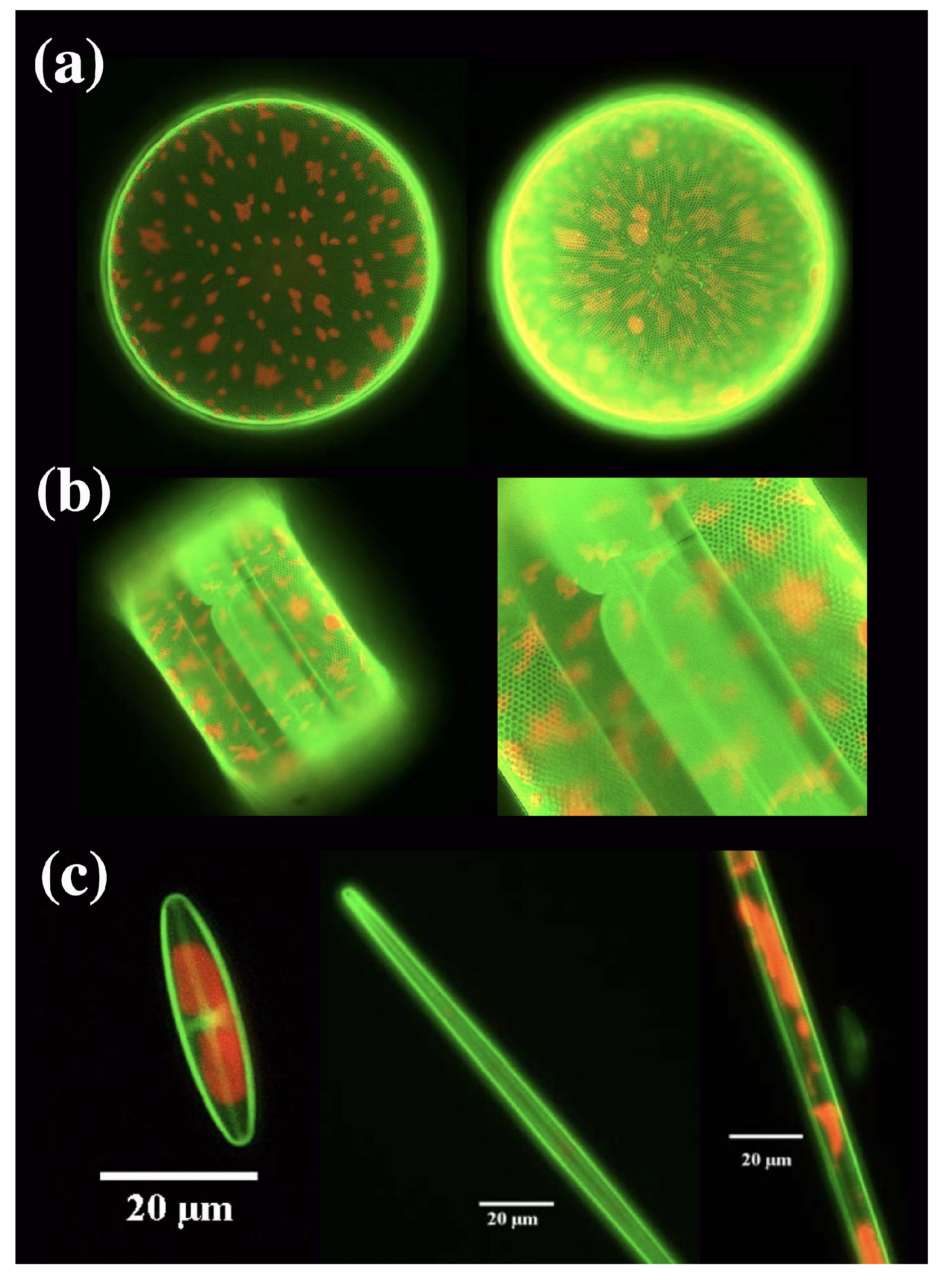
3.1. Frustule Metabolic Doping
3.2. Frustule Metalization for Applications in Plasmonics
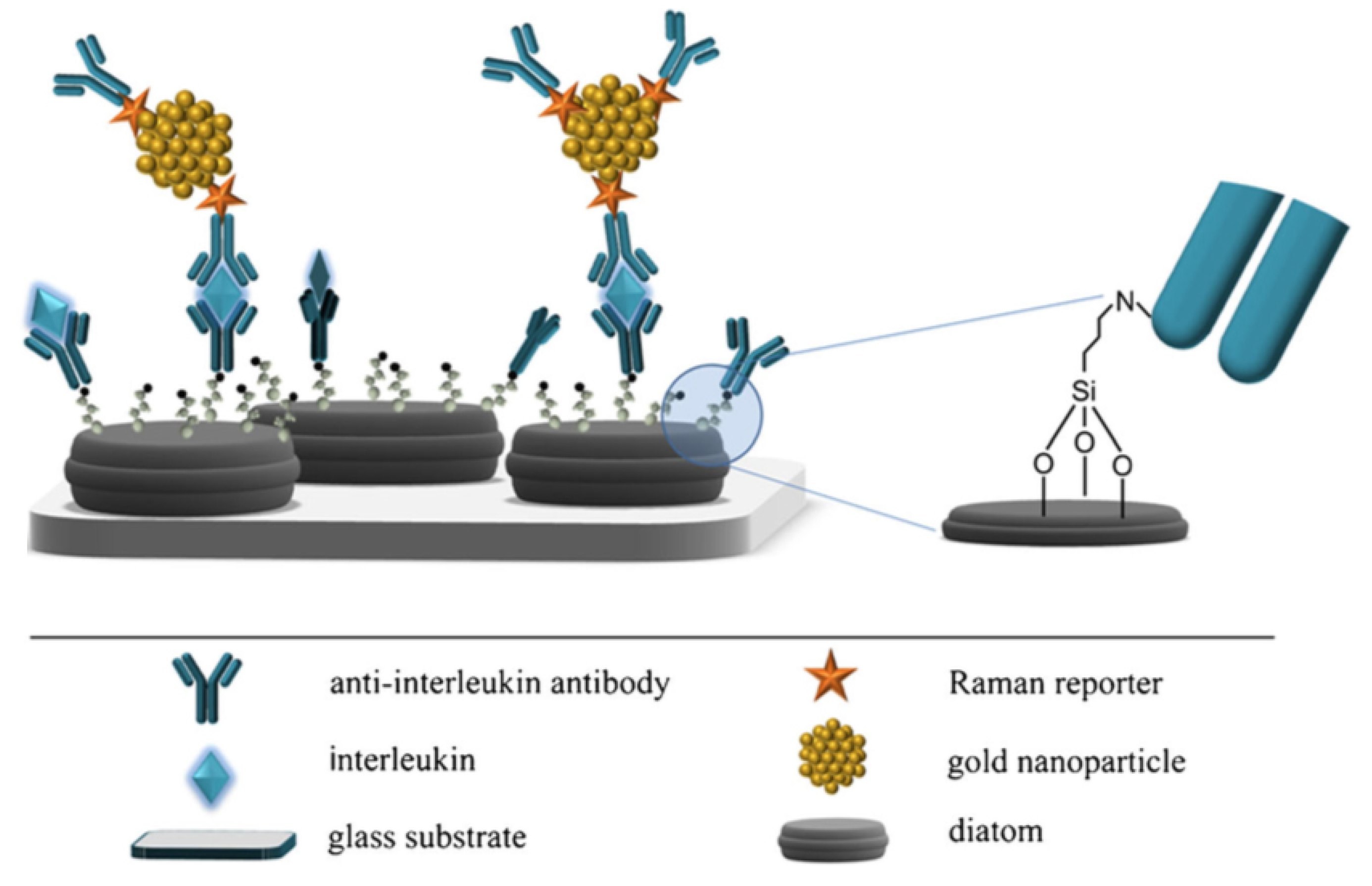
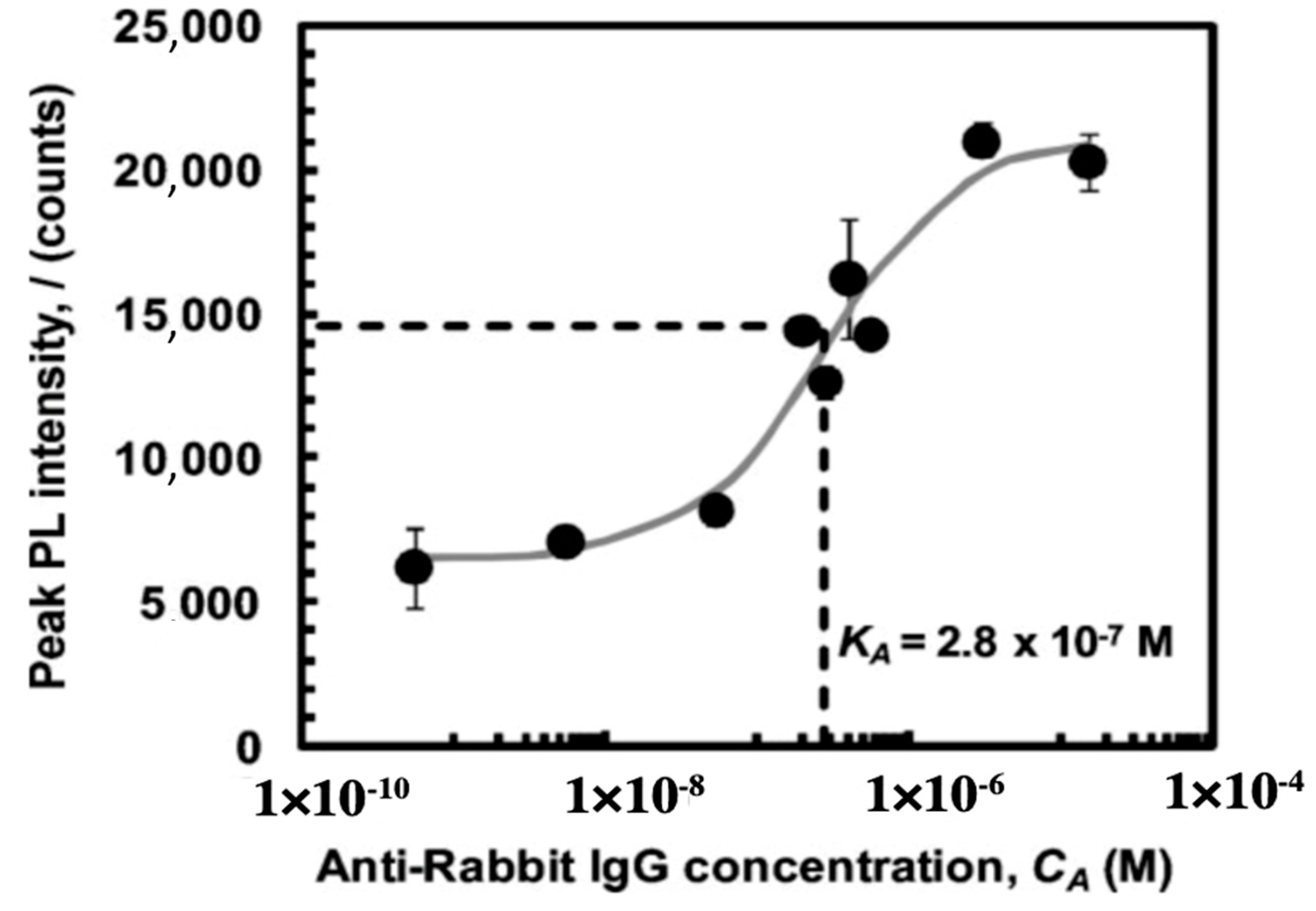
3.3. Fustule Functionalization for Protein Immobilization
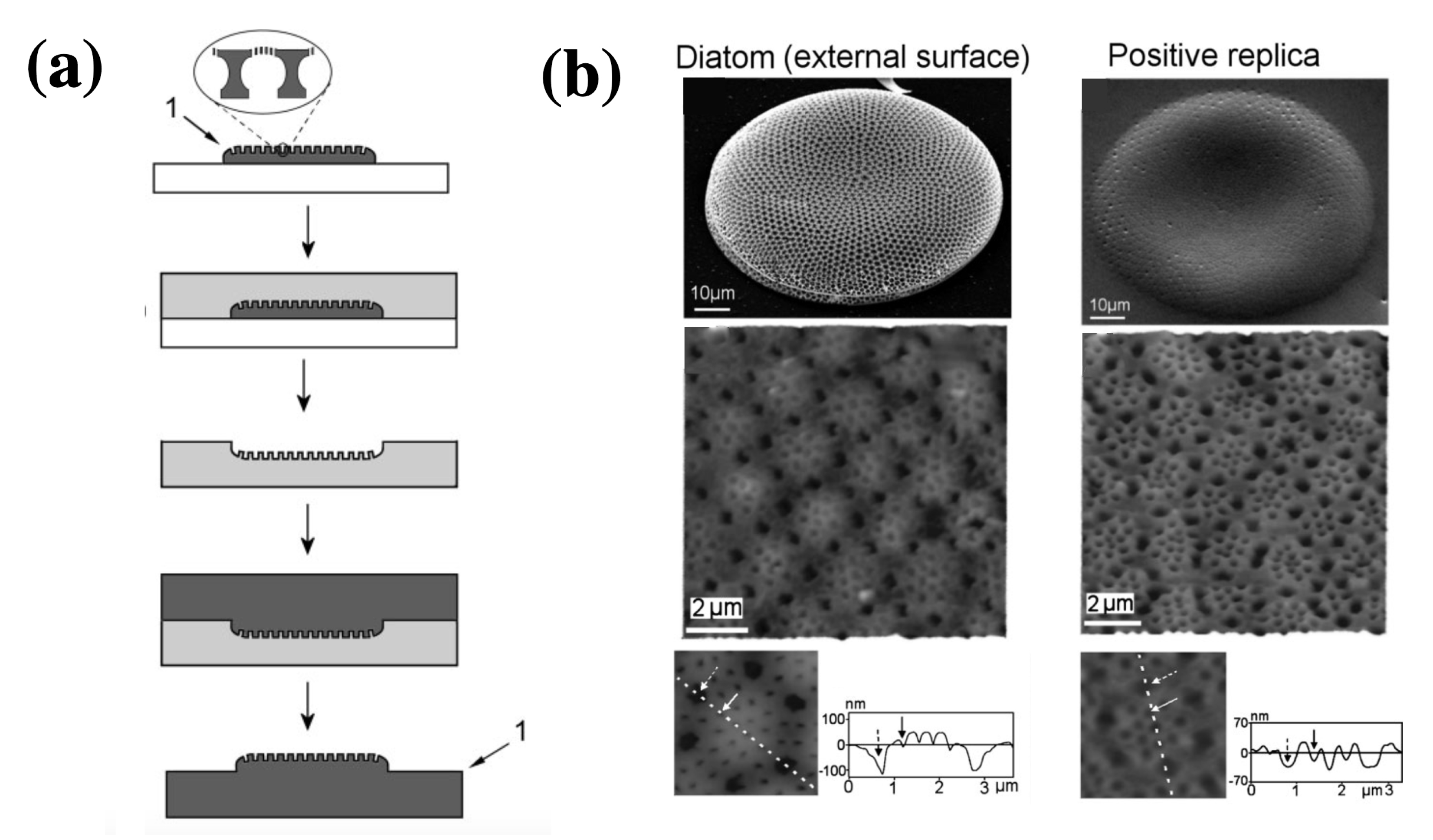
3.4. Frustule Replicas
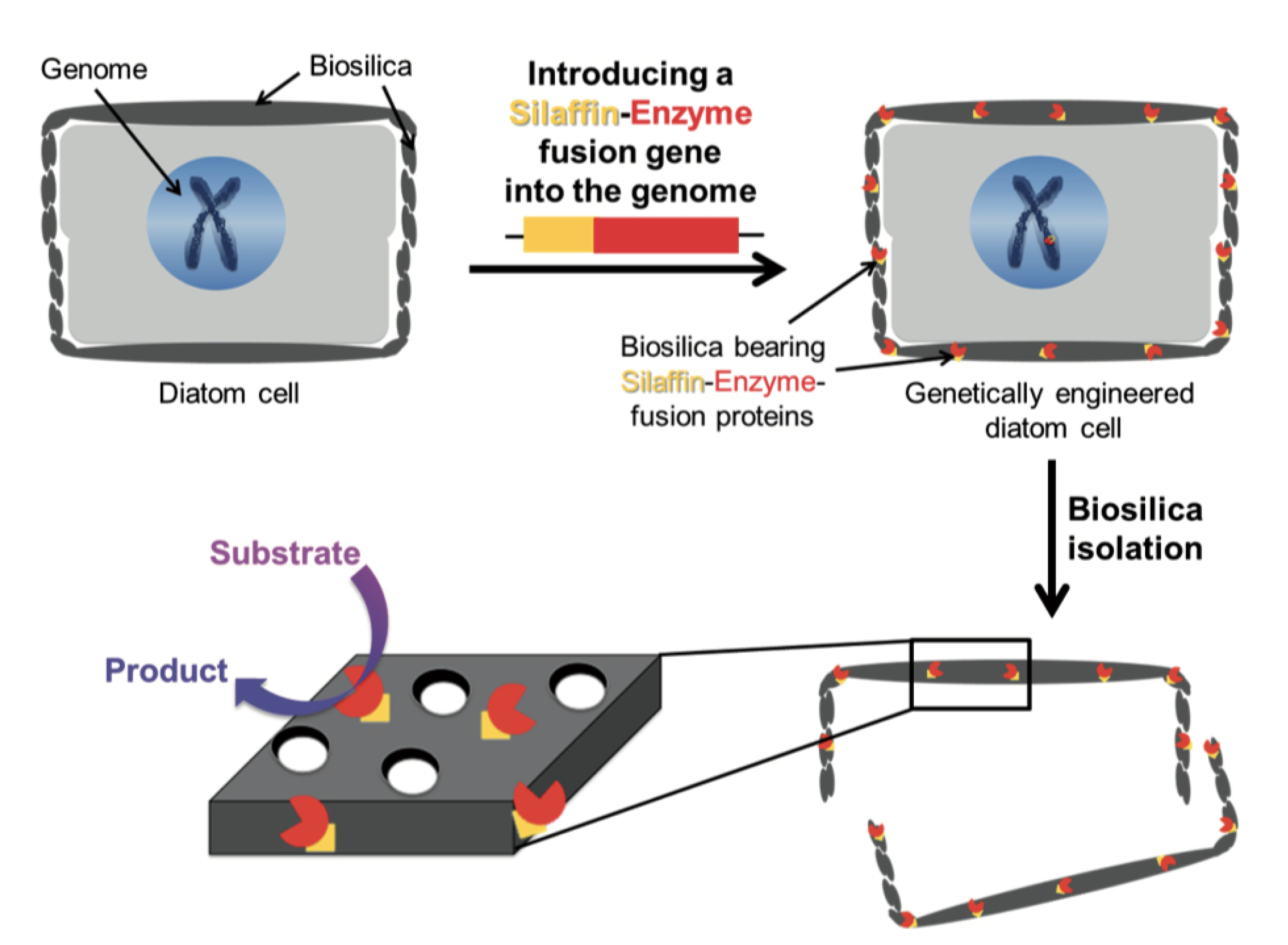
3.5. Frustule Genetic Modification
3.6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kumari, E.; Görlich, S.; Poulsen, N.; Kröger, N. Genetically Programmed Regioselective Immobilization of Enzymes in Biosilica Microparticles. Adv. Funct. Mater. 2020, 30, 2000442. [Google Scholar] [CrossRef]
- Livage, J. Bioinspired nanostructured materials. Comptes Rendus Chim. 2018, 21, 969–973. [Google Scholar] [CrossRef]
- Otzen, D. The role of proteins in biosilicification. Scientifica 2012, 2012, 867562. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.C.; Keeling-Tucker, T. Biosilicification: The role of the organic matrix in structure control. JBIC J. Biol. Inorg. Chem. 2000, 5, 537–550. [Google Scholar] [CrossRef]
- Hildebrand, M.; Lerch, S.J.; Shrestha, R.P. Understanding diatom cell wall silicification—Moving forward. Front. Mar. Sci. 2018, 5, 125. [Google Scholar] [CrossRef]
- Round, F.E.; Crawford, R.M.; Mann, D.G. Diatoms: Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Nelson, D.M.; Tréguer, P.; Brzezinski, M.A.; Leynaert, A.; Quéguiner, B. Production and dissolution of biogenic silica in the ocean: Revised global estimates, comparison with regional data and relationship to biogenic sedimentation. Glob. Biogeochem. Cycles 1995, 9, 359–372. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Raven, J.A. Aquatic Photosynthesis; Princeton University Press: Princeton, NJ, USA, 2013. [Google Scholar]
- Heintze, C.; Formanek, P.; Pohl, D.; Hauptstein, J.; Rellinghaus, B.; Kröger, N. An intimate view into the silica deposition vesicles of diatoms. BMC Mater. 2020, 2, 11. [Google Scholar] [CrossRef]
- De Tommasi, E.; Gielis, J.; Rogato, A. Diatom frustule morphogenesis and function: A multidisciplinary survey. Mar. Genom. 2017, 35, 1–18. [Google Scholar] [CrossRef]
- Hamm, C.E.; Merkel, R.; Springer, O.; Jurkojc, P.; Maier, C.; Prechtel, K.; Smetacek, V. Architecture and material properties of diatom shells provide effective mechanical protection. Nature 2003, 421, 841–843. [Google Scholar] [CrossRef]
- Hale, M.S.; Mitchell, J.G. Functional morphology of diatom frustule microstructures: Hydrodynamic control of Brownian particle diffusion and advection. Aquat. Microb. Ecol. 2001, 24, 287–295. [Google Scholar] [CrossRef]
- Hale, M.S.; Mitchell, J.G. Effects of particle size, flow velocity, and cell surface microtopography on the motion of submicrometer particles over diatoms. Nano Lett. 2002, 2, 657–663. [Google Scholar] [CrossRef]
- Waite, A.; Fisher, A.; Thompson, P.A.; Harrison, P.J. Sinking rate versus cell volume relationships illuminate sinking rate control mechanisms in marine diatoms. Mar. Ecol. Prog. Ser. 1997, 157, 97–108. [Google Scholar] [CrossRef]
- De Tommasi, E. Light manipulation by single cells: The case of diatoms. J. Spectrosc. 2016, 2016, 2490128. [Google Scholar] [CrossRef]
- De Tommasi, E.; De Luca, A.; Lavanga, L.; Dardano, P.; De Stefano, M.; De Stefano, L.; Langella, C.; Rendina, I.; Dholakia, K.; Mazilu, M. Biologically enabled sub-diffractive focusing. Opt. Express 2014, 22, 27214–27227. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhen, L.; Ren, F.; Campbell, J.; Rorrer, G.L.; Wang, A.X. Ultra-sensitive immunoassay biosensors using hybrid plasmonic-biosilica nanostructured materials. J. Biophotonics 2015, 8, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Huang, C.H.; Shian, S.; Sandhage, K.H. Rapid Hydrolysis of Organophosphorous Esters Induced by Nanostructured, Fluorine-Doped Titania Replicas of Diatom Frustules. J. Am. Ceram. Soc. 2007, 90, 1632–1636. [Google Scholar] [CrossRef]
- Squire, K.; Kong, X.; LeDuff, P.; Rorrer, G.L.; Wang, A.X. Photonic crystal enhanced fluorescence immunoassay on diatom biosilica. J. Biophotonics 2018, 11, e201800009. [Google Scholar] [CrossRef]
- Toster, J.; Iyer, K.S.; Xiang, W.; Rosei, F.; Spiccia, L.; Raston, C.L. Diatom frustules as light traps enhance DSSC efficiency. Nanoscale 2013, 5, 873–876. [Google Scholar] [CrossRef]
- Aw, M.S.; Simovic, S.; Addai-Mensah, J.; Losic, D. Silica microcapsules from diatoms as new carrier for delivery of therapeutics. Nanomedicine 2011, 6, 1159–1173. [Google Scholar] [CrossRef]
- Gordon, R.; Losic, D.; Tiffany, M.A.; Nagy, S.S.; Sterrenburg, F.A. The glass menagerie: Diatoms for novel applications in nanotechnology. Trends Biotechnol. 2009, 27, 116–127. [Google Scholar] [CrossRef]
- Friedrichs, L.; Maier, M.; Hamm, C. A new method for exact three-dimensional reconstructions of diatom frustules. J. Microsc. 2012, 248, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Morales, L.V.; Sigman, D.M.; Horn, M.G.; Robinson, R.S. Cleaning methods for the isotopic determination of diatombound nitrogen in non-fossil diatom frustules. Limnol. Oceanogr. Methods 2013, 11, 101–112. [Google Scholar] [CrossRef]
- Romann, J.; Chauton, M.S.; Hanetho, S.M.; Vebner, M.; Heldal, M.; Thaulow, C.; Vadstein, O.; Tranell, G.; Einarsrud, M.A. Diatom frustules as a biomaterial: Effects of chemical treatment on organic material removal and mechanical properties in cleaned frustules from two Coscinodiscus species. J. Porous Mater. 2016, 23, 905–910. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, D.; Cai, J.; Pan, J.; Chen, M.; Li, A.; Jiang, Y. Biosilica structures obtained from Nitzschia, Ditylum, Skeletonema, and Coscinodiscus diatom by a filtration-aided acid cleaning method. Appl. Microbiol. Biotechnol. 2012, 95, 1165–1178. [Google Scholar] [CrossRef]
- Gholami, P.; Khataee, A.; Bhatnagar, A. Environmentally superior cleaning of diatom frustules using sono-Fenton process: Facile fabrication of nanoporous silica with homogeneous morphology and controlled size. Ultrason. Sonochem. 2020, 64, 105044. [Google Scholar] [CrossRef]
- Losic, D.; Short, K.; Mitchell, J.G.; Lal, R.; Voelcker, N.H. AFM nanoindentations of diatom biosilica surfaces. Langmuir 2007, 23, 5014–5021. [Google Scholar] [CrossRef]
- Brzozowska, W.; Sprynskyy, M.; Wojtczak, I.; Dąbek, P.; Witkowski, A.; Buszewski, B. “Outsourcing” Diatoms in Fabrication of Metal-Doped 3D Biosilica. Materials 2020, 13, 2576. [Google Scholar] [CrossRef]
- Jeffryes, C.; Gutu, T.; Jiao, J.; Rorrer, G.L. Two-stage photobioreactor process for the metabolic insertion of nanostructured germanium into the silica microstructure of the diatom Pinnularia sp. Mater. Sci. Eng. C 2008, 28, 107–118. [Google Scholar] [CrossRef]
- De Tommasi, E.; Congestri, R.; Dardano, P.; De Luca, A.C.; Managò, S.; Rea, I.; De Stefano, M. UV-shielding and wavelength conversion by centric diatom nanopatterned frustules. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Qin, T.; Gutu, T.; Jiao, J.; Chang, C.H.; Rorrer, G.L. Photoluminescence of silica nanostructures from bioreactor culture of marine diatom Nitzschia frustulum. J. Nanosci. Nanotechnol. 2008, 8, 2392–2398. [Google Scholar] [CrossRef]
- Jeffryes, C.; Solanki, R.; Rangineni, Y.; Wang, W.; Chang, C.h.; Rorrer, G.L. Electroluminescence and photoluminescence from nanostructured diatom frustules containing metabolically inserted germanium. Adv. Mater. 2008, 20, 2633–2637. [Google Scholar] [CrossRef]
- Gössling, J.W. Biophotonics of Diatoms: Linking Frustule Structure to Photobiology. Ph.D. Thesis, Department of Biology, Faculty of Science, University of Copenhagen, Copenhagen, Denmark, 2017. [Google Scholar]
- Townley, H.E.; Woon, K.L.; Payne, F.P.; White-Cooper, H.; Parker, A.R. Modification of the physical and optical properties of the frustule of the diatom Coscinodiscus wailesii by nickel sulfate. Nanotechnology 2007, 18, 295101. [Google Scholar] [CrossRef]
- Jeffryes, C.; Gutu, T.; Jiao, J.; Rorrer, G.L. Metabolic insertion of nanostructured TiO2 into the patterned biosilica of the diatom Pinnularia sp. by a two-stage bioreactor cultivation process. ACS Nano 2008, 2, 2103–2112. [Google Scholar] [CrossRef] [PubMed]
- Jeffryes, C.; Campbell, J.; Li, H.; Jiao, J.; Rorrer, G. The potential of diatom nanobiotechnology for applications in solar cells, batteries, and electroluminescent devices. Energy Environ. Sci. 2011, 4, 3930–3941. [Google Scholar] [CrossRef]
- Gautam, S.; Kashyap, M.; Gupta, S.; Kumar, V.; Schoefs, B.; Gordon, R.; Jeffryes, C.; Joshi, K.B.; Vinayak, V. Metabolic engineering of tio 2 nanoparticles in nitzschia palea to form diatom nanotubes: An ingredient for solar cells to produce electricity and biofuel. RSC Adv. 2016, 6, 97276–97284. [Google Scholar] [CrossRef]
- Bandara, T.; Furlani, M.; Albinsson, I.; Wulff, A.; Mellander, B.E. Diatom frustules enhancing the efficiency of gel polymer electrolyte based dye-sensitized solar cells with multilayer photoelectrodes. Nanoscale Adv. 2020, 2, 199–209. [Google Scholar] [CrossRef]
- Xiao, X.; Zhang, X.; Su, H.; Chen, S.; He, Z.; Zhao, C.; Yang, S. A Visible-NIR Responsive Dye-Sensitized Solar Cell Based on Diatom Frustules and Cosensitization of Photopigments from Diatom and Purple Bacteria. J. Chem. 2020, 2020, 1710989. [Google Scholar] [CrossRef]
- Machill, S.; Köhler, L.; Ueberlein, S.; Hedrich, R.; Kunaschk, M.; Paasch, S.; Schulze, R.; Brunner, E. Analytical studies on the incorporation of aluminium in the cell walls of the marine diatom Stephanopyxis turris. BioMetals 2013, 26, 141–150. [Google Scholar] [CrossRef]
- Zhang, G.; Jiang, W.; Wang, L.; Liao, X.; Liu, P.; Deng, X.; Li, J. Preparation of silicate-based red phosphors with a patterned nanostructure via metabolic insertion of europium in marine diatoms. Mater. Lett. 2013, 110, 253–255. [Google Scholar] [CrossRef]
- Leone, G.; Vona, D.; Presti, M.L.; Urbano, L.; Cicco, S.; Gristina, R.; Palumbo, F.; Ragni, R.; Farinola, G. Ca 2+-in vivo doped biosilica from living Thalassiosira weissflogii diatoms: Investigation on Saos-2 biocompatibility. MRS Adv. 2017, 2, 1047–1058. [Google Scholar] [CrossRef]
- Basharina, T.N.; Danilovtseva, E.N.; Zelinskiy, S.N.; Klimenkov, I.V.; Likhoshway, Y.V.; Annenkov, V.V. The effect of titanium, zirconium and tin on the growth of diatom Synedra acus and morphology of its silica valves. Silicon 2012, 4, 239–249. [Google Scholar] [CrossRef]
- Gannavarapu, K.P.; Ganesh, V.; Thakkar, M.; Mitra, S.; Dandamudi, R.B. Nanostructured Diatom-ZrO2 composite as a selective and highly sensitive enzyme free electrochemical sensor for detection of methyl parathion. Sens. Actuators B Chem. 2019, 288, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Kucki, M. Biological Photonic Crystals: Diatoms Dye Functionalization of Biological Silica Nanostructures. Ph.D. Thesis, University of Kassel, Kassel, Germany, 2009. [Google Scholar]
- Kucki, M.; Fuhrmann-Lieker, T. Staining diatoms with rhodamine dyes: Control of emission colour in photonic biocomposites. J. R. Soc. Interface 2012, 9, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Megens, M.; Wijnhoven, J.E.; Lagendijk, A.; Vos, W.L. Fluorescence lifetimes and linewidths of dye in photonic crystals. Phys. Rev. A 1999, 59, 4727. [Google Scholar] [CrossRef]
- Yoshie, T.; Shchekin, O.; Chen, H.; Deppe, D.; Scherer, A. Quantum dot photonic crystal lasers. Electron. Lett. 2002, 38, 967–968. [Google Scholar] [CrossRef]
- Ragni, R.; Scotognella, F.; Vona, D.; Moretti, L.; Altamura, E.; Ceccone, G.; Mehn, D.; Cicco, S.R.; Palumbo, F.; Lanzani, G.; et al. Hybrid photonic nanostructures by in vivo incorporation of an organic fluorophore into diatom algae. Adv. Funct. Mater. 2018, 28, 1706214. [Google Scholar] [CrossRef]
- Fuhrmann, T.; Landwehr, S.; El Rharbi-Kucki, M.; Sumper, M. Diatoms as living photonic crystals. Appl. Phys. B 2004, 78, 257–260. [Google Scholar] [CrossRef]
- Yamanaka, S.; Yano, R.; Usami, H.; Hayashida, N.; Ohguchi, M.; Takeda, H.; Yoshino, K. Optical properties of diatom silica frustule with special reference to blue light. J. Appl. Phys. 2008, 103, 074701. [Google Scholar] [CrossRef]
- De Stefano, L.; Maddalena, P.; Moretti, L.; Rea, I.; Rendina, I.; De Tommasi, E.; Mocella, V.; De Stefano, M. Nano-biosilica from marine diatoms: A brand new material for photonic applications. Superlattices Microstruct. 2009, 46, 84–89. [Google Scholar] [CrossRef]
- Kieu, K.; Li, C.; Fang, Y.; Cohoon, G.; Herrera, O.; Hildebrand, M.; Sandhage, K.; Norwood, R.A. Structure-based optical filtering by the silica microshell of the centric marine diatom Coscinodiscus wailesii. Opt. Express 2014, 22, 15992–15999. [Google Scholar] [CrossRef]
- Mcheik, A.; Cassaignon, S.; Livage, J.; Gibaud, A.; Berthier, S.; Lopez, P.J. Optical properties of nanostructured silica structures from marine organisms. Front. Mar. Sci. 2018, 5, 123. [Google Scholar] [CrossRef]
- Goessling, J.W.; Su, Y.; Cartaxana, P.; Maibohm, C.; Rickelt, L.F.; Trampe, E.C.; Walby, S.L.; Wangpraseurt, D.; Wu, X.; Ellegaard, M.; et al. Structure-based optics of centric diatom frustules: Modulation of the in vivo light field for efficient diatom photosynthesis. New Phytol. 2018, 219, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Goessling, J.W.; Gonzalez, A.A.S.; Paul, V.; Ashworth, M.; Manning, S.R.; Lopez-Garcia, M. Biosilica slab photonic crystals as an alternative to cleanroom fabrication. Faraday Discuss. 2020, 223. [Google Scholar] [CrossRef]
- Goessling, J.W.; Wardley, W.P.; Lopez-Garcia, M. Highly Reproducible, Bio-Based Slab Photonic Crystals Grown by Diatoms. Adv. Sci. 2020, 7, 1903726. [Google Scholar] [CrossRef] [PubMed]
- Sakoda, K. Optical Properties of Photonic Crystals; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2004; Volume 80. [Google Scholar]
- Joannopoulos, J.D.; Johnson, S.G.; Winn, J.N.; Meade, R.D. Molding the Flow of Light; Princeton University Press: Princeton, NJ, USA, 2008. [Google Scholar]
- Maier, S.A. Plasmonics: Fundamentals and Applications; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Le Ru, E.; Etchegoin, P. Principles of Surface-Enhanced Raman Spectroscopy: Furthermore, Related Plasmonic Effects; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Lee, P.; Meisel, D. Adsorption and surface-enhanced Raman of dyes on silver and gold sols. J. Phys. Chem. 1982, 86, 3391–3395. [Google Scholar] [CrossRef]
- Liz-Marzán, L.M.; Giersig, M.; Mulvaney, P. Synthesis of nanosized gold-silica core-shell particles. Langmuir 1996, 12, 4329–4335. [Google Scholar] [CrossRef]
- Tao, A.; Kim, F.; Hess, C.; Goldberger, J.; He, R.; Sun, Y.; Xia, Y.; Yang, P. Langmuir-Blodgett silver nanowire monolayers for molecular sensing using surface-enhanced Raman spectroscopy. Nano Lett. 2003, 3, 1229–1233. [Google Scholar] [CrossRef]
- Sun, Y.; Xia, Y. Shape-controlled synthesis of gold and silver nanoparticles. Science 2002, 298, 2176–2179. [Google Scholar] [CrossRef]
- McLellan, J.M.; Li, Z.Y.; Siekkinen, A.R.; Xia, Y. The SERS activity of a supported Ag nanocube strongly depends on its orientation relative to laser polarization. Nano Lett. 2007, 7, 1013–1017. [Google Scholar] [CrossRef]
- Hrelescu, C.; Sau, T.K.; Rogach, A.L.; Jäckel, F.; Feldmann, J. Single gold nanostars enhance Raman scattering. Appl. Phys. Lett. 2009, 94, 153113. [Google Scholar] [CrossRef]
- Matteini, P.; Cottat, M.; Tavanti, F.; Panfilova, E.; Scuderi, M.; Nicotra, G.; Menziani, M.C.; Khlebtsov, N.; de Angelis, M.; Pini, R. Site-selective surface-enhanced Raman detection of proteins. ACS Nano 2017, 11, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Muehlschlegel, P.; Eisler, H.J.; Martin, O.J.; Hecht, B.; Pohl, D. Resonant optical antennas. Science 2005, 308, 1607–1609. [Google Scholar] [CrossRef] [PubMed]
- Crozier, K.B.; Zhu, W.; Wang, D.; Lin, S.; Best, M.D.; Camden, J.P. Plasmonics for surface enhanced raman scattering: Nanoantennas for single molecules. IEEE J. Sel. Top. Quantum Electron. 2013, 20, 152–162. [Google Scholar] [CrossRef]
- Jiao, Y.; Ryckman, J.D.; Ciesielski, P.N.; Escobar, C.A.; Jennings, G.K.; Weiss, S.M. Patterned nanoporous gold as an effective SERS template. Nanotechnology 2011, 22, 295302. [Google Scholar] [CrossRef]
- Kim, S.m.; Zhang, W.; Cunningham, B.T. Coupling discrete metal nanoparticles to photonic crystal surface resonant modes and application to Raman spectroscopy. Opt. Express 2010, 18, 4300–4309. [Google Scholar] [CrossRef]
- Ricciardi, A.; Consales, M.; Quero, G.; Crescitelli, A.; Esposito, E.; Cusano, A. Versatile optical fiber nanoprobes: From plasmonic biosensors to polarization-sensitive devices. ACS Photonics 2014, 1, 69–78. [Google Scholar] [CrossRef]
- De Luca, A.C.; Reader-Harris, P.; Mazilu, M.; Mariggio, S.; Corda, D.; Di Falco, A. Reproducible surface-enhanced Raman quantification of biomarkers in multicomponent mixtures. ACS Nano 2014, 8, 2575–2583. [Google Scholar] [CrossRef]
- De Angelis, F.; Malerba, M.; Patrini, M.; Miele, E.; Das, G.; Toma, A.; Zaccaria, R.P.; Di Fabrizio, E. 3D hollow nanostructures as building blocks for multifunctional plasmonics. Nano Lett. 2013, 13, 3553–3558. [Google Scholar] [CrossRef]
- Payne, E.K.; Rosi, N.L.; Xue, C.; Mirkin, C.A. Sacrificial biological templates for the formation of nanostructured metallic microshells. Angew. Chem. 2005, 117, 5192–5195. [Google Scholar] [CrossRef]
- Ren, F.; Campbell, J.; Rorrer, G.L.; Wang, A.X. Surface-enhanced Raman spectroscopy sensors from nanobiosilica with self-assembled plasmonic nanoparticles. IEEE J. Sel. Top. Quantum Electron. 2014, 20, 127–132. [Google Scholar]
- Kamińska, A.; Sprynskyy, M.; Winkler, K.; Szymborski, T. Ultrasensitive SERS immunoassay based on diatom biosilica for detection of interleukins in blood plasma. Anal. Bioanal. Chem. 2017, 409, 6337–6347. [Google Scholar] [CrossRef] [PubMed]
- Managò, S.; Zito, G.; Rogato, A.; Casalino, M.; Esposito, E.; De Luca, A.C.; De Tommasi, E. Bioderived three-dimensional hierarchical nanostructures as efficient surface-enhanced raman scattering substrates for cell membrane probing. ACS Appl. Mater. Interfaces 2018, 10, 12406–12416. [Google Scholar] [CrossRef] [PubMed]
- Sivashanmugan, K.; Squire, K.; Kraai, J.A.; Tan, A.; Zhao, Y.; Rorrer, G.L.; Wang, A.X. Biological Photonic Crystal-Enhanced Plasmonic Mesocapsules: Approaching Single-Molecule Optofluidic-SERS Sensing. Adv. Opt. Mater. 2019, 7, 1900415. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Chen, V.W.; Cai, Y.; Berrigan, J.D.; Marder, S.R.; Perry, J.W.; Sandhage, K.H. Biologically Enabled Syntheses of Freestanding Metallic Structures Possessing Subwavelength Pore Arrays for Extraordinary (Surface Plasmon-Mediated) Infrared Transmission. Adv. Funct. Mater. 2012, 22, 2550–2559. [Google Scholar] [CrossRef]
- Pacifici, D.; Lezec, H.J.; Sweatlock, L.A.; Walters, R.J.; Atwater, H.A. Universal optical transmission features in periodic and quasiperiodic hole arrays. Opt. Express 2008, 16, 9222–9238. [Google Scholar] [CrossRef]
- Kröger, N.; Dubey, N.; Kumari, E. Immobilization of Proteins on Diatom Biosilica. In Diatom Nanotechnology; Royal Society of Chemistry: London, UK, 2017; pp. 126–149. [Google Scholar]
- Dill, K.A.; Ghosh, K.; Schmit, J.D. Physical limits of cells and proteomes. Proc. Natl. Acad. Sci. USA 2011, 108, 17876–17882. [Google Scholar] [CrossRef]
- Townley, H.E.; Parker, A.R.; White-Cooper, H. Exploitation of diatom frustules for nanotechnology: Tethering active biomolecules. Adv. Funct. Mater. 2008, 18, 369–374. [Google Scholar] [CrossRef]
- De Stefano, L.; Lamberti, A.; Rotiroti, L.; De Stefano, M. Interfacing the nanostructured biosilica microshells of the marine diatom Coscinodiscus wailesii with biological matter. Acta Biomater. 2008, 4, 126–130. [Google Scholar] [CrossRef]
- De Tommasi, E.; Rea, I.; Rendina, I.; Rotiroti, L.; De Stefano, L. Protein conformational changes revealed by optical spectroscopic reflectometry in porous silicon multilayers. J. Phys. Condens. Matter 2008, 21, 035115. [Google Scholar] [CrossRef]
- De Tommasi, E.; De Stefano, L.; Rea, I.; Di Sarno, V.; Rotiroti, L.; Arcari, P.; Lamberti, A.; Sanges, C.; Rendina, I. Porous silicon based resonant mirrors for biochemical sensing. Sensors 2008, 8, 6549–6556. [Google Scholar] [CrossRef]
- Hilkens, J.; Ligtenberg, M.J.; Vos, H.L.; Litvinov, S.V. Cell membrane-associated mucins and their adhesion-modulating property. Trends Biochem. Sci. 1992, 17, 359–363. [Google Scholar] [CrossRef]
- Bayramoglu, G.; Akbulut, A.; Arica, M.Y. Immobilization of tyrosinase on modified diatom biosilica: Enzymatic removal of phenolic compounds from aqueous solution. J. Hazard. Mater. 2013, 244, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Bayramoglu, G.; Akbulut, A.; Ozalp, V.C.; Arica, M.Y. Immobilized lipase on micro-porous biosilica for enzymatic transesterification of algal oil. Chem. Eng. Res. Des. 2015, 95, 12–21. [Google Scholar] [CrossRef]
- De Stefano, L.; Rotiroti, L.; De Stefano, M.; Lamberti, A.; Lettieri, S.; Setaro, A.; Maddalena, P. Marine diatoms as optical biosensors. Biosens. Bioelectron. 2009, 24, 1580–1584. [Google Scholar] [CrossRef]
- Losic, D.; Mitchell, J.G.; Voelcker, N.H. Diatomaceous lessons in nanotechnology and advanced materials. Adv. Mater. 2009, 21, 2947–2958. [Google Scholar] [CrossRef]
- Ragni, R.; Cicco, S.R.; Vona, D.; Farinola, G.M. Multiple routes to smart nanostructured materials from diatom microalgae: A chemical perspective. Adv. Mater. 2018, 30, 1704289. [Google Scholar] [CrossRef]
- Panwar, V.; Dutta, T. Diatom biogenic silica as a felicitous platform for biochemical engineering: Expanding frontiers. ACS Appl. Bio Mater. 2019, 2, 2295–2316. [Google Scholar] [CrossRef]
- De Stefano, L.; Rendina, I.; De Stefano, M.; Bismuto, A.; Maddalena, P. Marine diatoms as optical chemical sensors. Appl. Phys. Lett. 2005, 87, 233902. [Google Scholar] [CrossRef]
- Setaro, A.; Lettieri, S.; Maddalena, P.; De Stefano, L. Highly sensitive optochemical gas detection by luminescent marine diatoms. Appl. Phys. Lett. 2007, 91, 051921. [Google Scholar] [CrossRef]
- Lettieri, S.; Setaro, A.; De Stefano, L.; De Stefano, M.; Maddalena, P. The gas-detection properties of light-emitting diatoms. Adv. Funct. Mater. 2008, 18, 1257–1264. [Google Scholar] [CrossRef]
- Gale, D.K.; Gutu, T.; Jiao, J.; Chang, C.H.; Rorrer, G.L. Photoluminescence detection of biomolecules by antibody-functionalized diatom biosilica. Adv. Funct. Mater. 2009, 19, 926–933. [Google Scholar] [CrossRef]
- Losic, D.; Mitchell, J.G.; Lal, R.; Voelcker, N.H. Rapid fabrication of micro-and nanoscale patterns by replica molding from diatom biosilica. Adv. Funct. Mater. 2007, 17, 2439–2446. [Google Scholar] [CrossRef]
- Bao, Z.; Weatherspoon, M.R.; Shian, S.; Cai, Y.; Graham, P.D.; Allan, S.M.; Ahmad, G.; Dickerson, M.B.; Church, B.C.; Kang, Z.; et al. Chemical reduction of three-dimensional silica micro-assemblies into microporous silicon replicas. Nature 2007, 446, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Lerch, S.J.; Xu, L.; Li, X.; Chuang, Y.J.; Howe, J.Y.; Mahurin, S.M.; Dai, S.; Hildebrand, M. Electronically transparent graphene replicas of diatoms: A new technique for the investigation of frustule morphology. Sci. Rep. 2014, 4, 6117. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Liu, X.; Zheng, T.; Jiang, D.; Zhou, Z.; Liu, C.; Zhang, X.; Zhang, Y.; Losic, D. Tuning MnO2 to FeOOH replicas with bio-template 3D morphology as electrodes for high performance asymmetric supercapacitors. Chem. Eng. J. 2019, 370, 136–147. [Google Scholar] [CrossRef]
- Xu, M.; Gratson, G.M.; Duoss, E.B.; Shepherd, R.F.; Lewis, J.A. Biomimetic silicification of 3D polyamine-rich scaffolds assembled by direct ink writing. Soft Matter 2006, 2, 205–209. [Google Scholar] [CrossRef]
- Gratson, G.M.; Xu, M.; Lewis, J.A. Direct writing of three-dimensional webs. Nature 2004, 428, 386. [Google Scholar] [CrossRef]
- Xie, P.; Chen, Z.; Xu, J.; Xie, D.; Wang, X.; Cui, S.; Zhou, H.; Zhang, D.; Fan, T. Artificial ceramic diatoms with multiscale photonic architectures via nanoimprint lithography for CO2 photoreduction. J. Am. Ceram. Soc. 2019, 102, 4678–4687. [Google Scholar] [CrossRef]
- Kröger, N.; Bergsdorf, C.; Sumper, M. Frustulins: Domain conservation in a protein family associated with diatom cell walls. Eur. J. Biochem. 1996, 239, 259–264. [Google Scholar] [CrossRef]
- Heintze, C.; Formanek, P.; Pohl, D.; Hauptstein, J.; Rellinghaus, B.; Kröger, N. A New View into the Silica Deposition Vesicles of Diatoms. 2020. Available online: https://assets.researchsquare.com/files/rs-22354/v3/a9518908-f148-4d7c-9440-5424fc0fe15e.pdf (accessed on 15 June 2020).
- Kroth, P. Molecular biology and the biotechnological potential of diatoms. In Transgenic Microalgae As Green Cell Factories; Springer: Berlin/Heidelberg, Germany, 2007; pp. 23–33. [Google Scholar]
- Zaslavskaia, L.A.; Lippmeier, J.C.; Kroth, P.G.; Grossman, A.R.; Apt, K.E. Transformation of the diatom Phaeodactylum tricornutum (Bacillariophyceae) with a variety of selectable marker and reporter genes. J. Phycol. 2000, 36, 379–386. [Google Scholar] [CrossRef]
- Coesel, S.; Mangogna, M.; Ishikawa, T.; Heijde, M.; Rogato, A.; Finazzi, G.; Todo, T.; Bowler, C.; Falciatore, A. Diatom PtCPF1 is a new cryptochrome/photolyase family member with DNA repair and transcription regulation activity. EMBO Rep. 2009, 10, 655–661. [Google Scholar] [CrossRef] [PubMed]
- De Riso, V.; Raniello, R.; Maumus, F.; Rogato, A.; Bowler, C.; Falciatore, A. Gene silencing in the marine diatom Phaeodactylum tricornutum. Nucleic Acids Res. 2009, 37, e96. [Google Scholar] [CrossRef] [PubMed]
- Weyman, P.D.; Beeri, K.; Lefebvre, S.C.; Rivera, J.; McCarthy, J.K.; Heuberger, A.L.; Peers, G.; Allen, A.E.; Dupont, C.L. Inactivation of P haeodactylum tricornutum urease gene using transcription activator-like effector nuclease-based targeted mutagenesis. Plant Biotechnol. J. 2015, 13, 460–470. [Google Scholar] [CrossRef]
- Nymark, M.; Sharma, A.K.; Sparstad, T.; Bones, A.M.; Winge, P. A CRISPR/Cas9 system adapted for gene editing in marine algae. Sci. Rep. 2016, 6, 24951. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Daboussi, F. Genetic and metabolic engineering in diatoms. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160411. [Google Scholar] [CrossRef]
- Kroth, P.G.; Bones, A.M.; Daboussi, F.; Ferrante, M.I.; Jaubert, M.; Kolot, M.; Nymark, M.; Bártulos, C.R.; Ritter, A.; Russo, M.T.; et al. Genome editing in diatoms: Achievements and goals. Plant Cell Rep. 2018, 37, 1401–1408. [Google Scholar] [CrossRef]
- Görlich, S.; Pawolski, D.; Zlotnikov, I.; Kröger, N. Control of biosilica morphology and mechanical performance by the conserved diatom gene Silicanin-1. Commun. Biol. 2019, 2, 1–8. [Google Scholar] [CrossRef]
- Rogato, A.; Del Prete, S.; Nocentini, A.; Carginale, V.; Supuran, C.T.; Capasso, C. Phaeodactylum tricornutum as a model organism for testing the membrane penetrability of sulphonamide carbonic anhydrase inhibitors. J. Enzym. Inhib. Med. Chem. 2019, 34, 510–518. [Google Scholar] [CrossRef]
- Tesson, B.; Lerch, S.J.; Hildebrand, M. Characterization of a new protein family associated with the silica deposition vesicle membrane enables genetic manipulation of diatom silica. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Shrestha, R.P.; Hildebrand, M. Evidence for a regulatory role of diatom silicon transporters in cellular silicon responses. Eukaryot. Cell 2015, 14, 29–40. [Google Scholar] [CrossRef]
- Knight, M.J.; Senior, L.; Nancolas, B.; Ratcliffe, S.; Curnow, P. Direct evidence of the molecular basis for biological silicon transport. Nat. Commun. 2016, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Delalat, B.; Sheppard, V.C.; Ghaemi, S.R.; Rao, S.; Prestidge, C.A.; McPhee, G.; Rogers, M.L.; Donoghue, J.F.; Pillay, V.; Johns, T.G.; et al. Targeted drug delivery using genetically engineered diatom biosilica. Nat. Commun. 2015, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kotzsch, A.; Gröger, P.; Pawolski, D.; Bomans, P.H.; Sommerdijk, N.A.; Schlierf, M.; Kröger, N. Silicanin-1 is a conserved diatom membrane protein involved in silica biomineralization. BMC Biol. 2017, 15, 65. [Google Scholar] [CrossRef] [PubMed]
- Wallace, A.K.; Chanut, N.; Voigt, C.A. Silica Nanostructures Produced Using Diatom Peptides with Designed Post-Translational Modifications. Adv. Funct. Mater. 2020, 30, 2000849. [Google Scholar] [CrossRef]
- Shrestha, R.P.; Tesson, B.; Norden-Krichmar, T.; Federowicz, S.; Hildebrand, M.; Allen, A.E. Whole transcriptome analysis of the silicon response of the diatom Thalassiosira pseudonana. BMC Genom. 2012, 13, 499. [Google Scholar] [CrossRef]
- Poulsen, N.; Berne, C.; Spain, J.; Kroeger, N. Silica immobilization of an enzyme through genetic engineering of the diatom Thalassiosira pseudonana. Angew. Chem. Int. Ed. 2007, 46, 1843–1846. [Google Scholar] [CrossRef]
- Sheppard, V.; Scheffel, A.; Poulsen, N.; Kröger, N. Live diatom silica immobilization of multimeric and redox-active enzymes. Appl. Environ. Microbiol. 2012, 78, 211–218. [Google Scholar] [CrossRef]
- Fernandes, F.M.; Coradin, T.; Aimé, C. Self-assembly in biosilicification and biotemplated silica materials. Nanomaterials 2014, 4, 792–812. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Genes or Related Proteins | Method | Reference |
|---|---|---|---|
| T. pseudonana | Silicic acid transporters (SITs 1,2,3). | Gateway technology for GFP fusion proteins. RNAi and antisense knockdown. | [121] |
| Target mutations. | [122] | ||
| Incorporated antibody-binding protein domains. | LiDSI-based method (live diatom silica immobilization). | [123] | |
| SAP1-2-3 (Silicalemma Associated Proteins). | Gateway technology for GFP fusion proteins. RNAi and antisense knockdown. | [120] | |
| Silacin-1 (Sin-1). | Cloning technology for GFP fusion proteins. | [124] | |
| CRISPR/Cas9-based approach. | [118] | ||
| Silaffin-1 (Sil1) and cingulin genes family (CinY1, -2,-3,-4 and CinW1, -2, -3). | LiDSI-based method (live diatom silica immobilization). | [1] | |
| C. fusiformis | Silaffin-1 (Sil1) (R5 peptide post-translational modifications). | E. coli expression system for the recombinant production of R5. | [125] |
| C. cryptica | ID g20669 (Sin1 homologue). | Cloning technology for GFP fusion proteins. | [109] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rogato, A.; De Tommasi, E. Physical, Chemical, and Genetic Techniques for Diatom Frustule Modification: Applications in Nanotechnology. Appl. Sci. 2020, 10, 8738. https://doi.org/10.3390/app10238738
Rogato A, De Tommasi E. Physical, Chemical, and Genetic Techniques for Diatom Frustule Modification: Applications in Nanotechnology. Applied Sciences. 2020; 10(23):8738. https://doi.org/10.3390/app10238738
Chicago/Turabian StyleRogato, Alessandra, and Edoardo De Tommasi. 2020. "Physical, Chemical, and Genetic Techniques for Diatom Frustule Modification: Applications in Nanotechnology" Applied Sciences 10, no. 23: 8738. https://doi.org/10.3390/app10238738
APA StyleRogato, A., & De Tommasi, E. (2020). Physical, Chemical, and Genetic Techniques for Diatom Frustule Modification: Applications in Nanotechnology. Applied Sciences, 10(23), 8738. https://doi.org/10.3390/app10238738