

Strike a Pose: Relationships Between Infants’ Motor Development and Visuospatial Representations of Bodies

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Infants’ Discrimination of Bodies
3. Face and Body Inversion Effects
4. Relationships with Motor Development
5. Current Study
6. Study 1A—7-Month-Old Infants’ Discrimination of Body Postures
6.1. Method
6.1.1. Participants
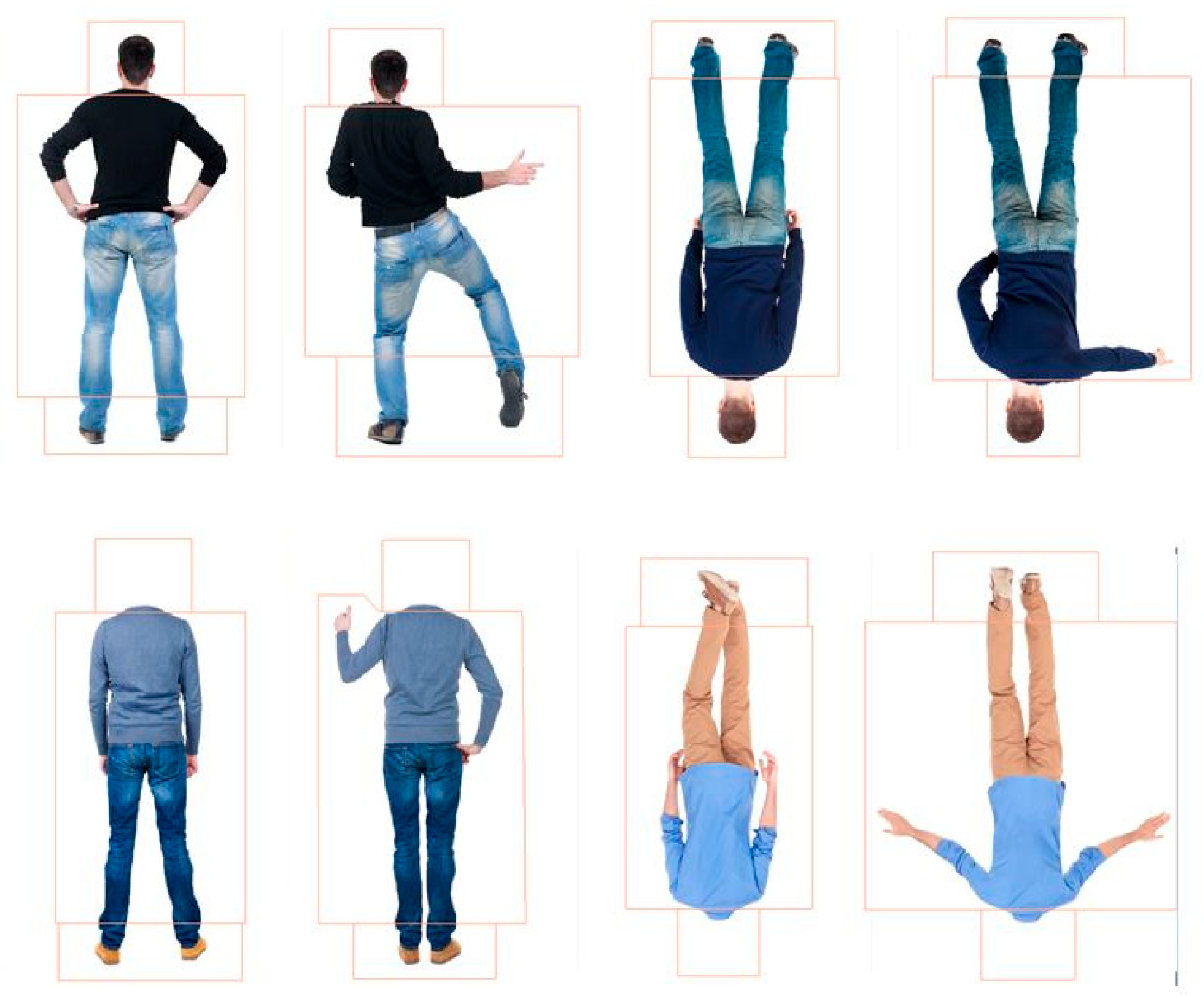
6.1.2. Materials
6.1.3. Procedure
6.2. Design and Analysis
6.3. Results
Novelty Preferences Compared to Chance
6.4. Discussion
7. Study 1B—7-Month-Old Infants’ Discrimination of About-Facing Body Postures
7.1. Method
7.1.1. Participants
7.1.2. Materials and Procedure
7.2. Results
Novelty Preferences Compared to Chance
7.3. Discussion
8. Study 2A—7-Month-Old Infants’ Discrimination of Similarly Varied Body Postures; 5 s Trial Durations
8.1. Method
8.1.1. Participants
8.1.2. Stimuli and Procedure
8.2. Results
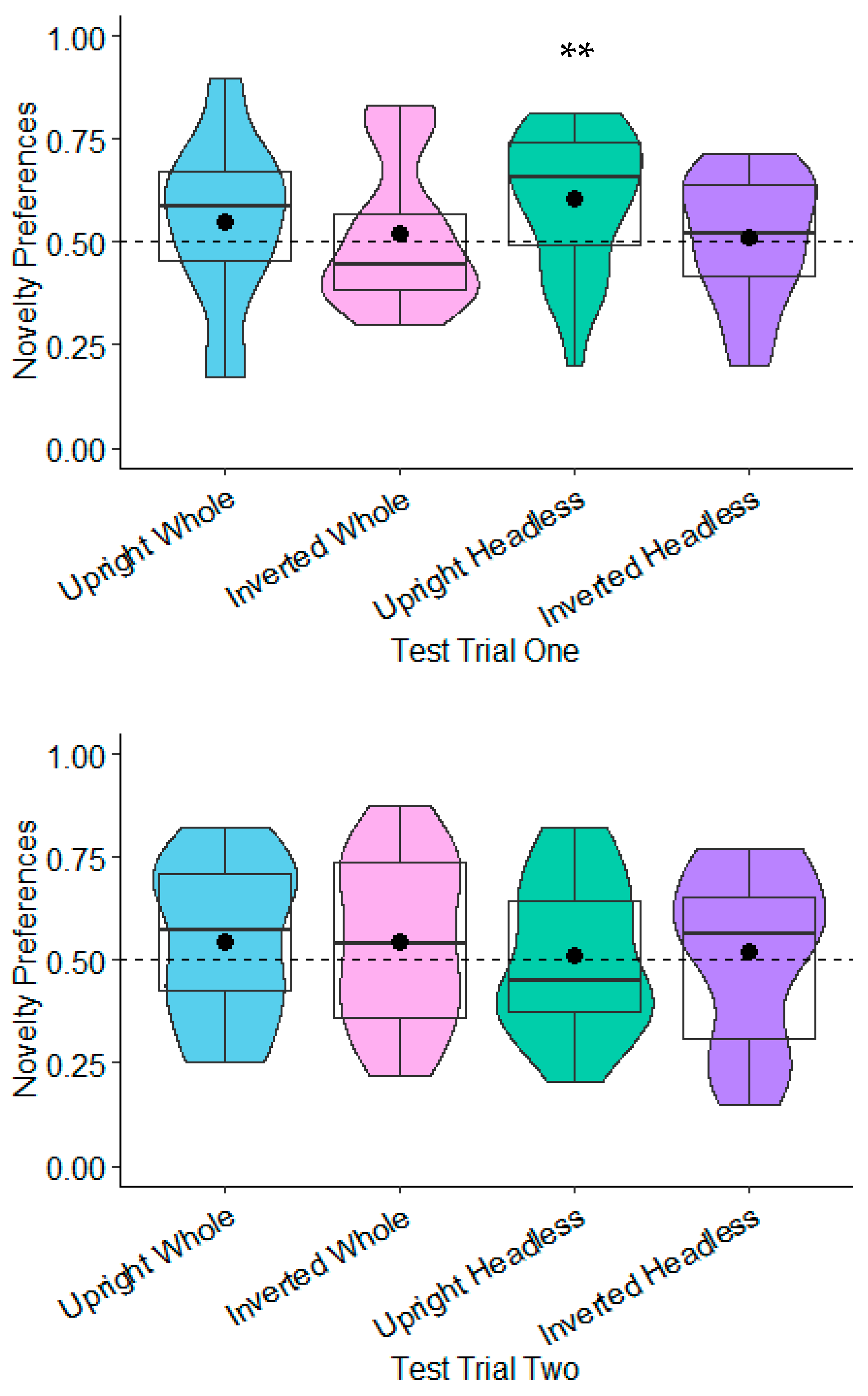
Novelty Preferences Compared to Chance
8.3. Discussion
9. Study 2B—7- and 9-Month-Old Infants’ Discrimination of Similarly Varied Body Postures with 8 s Trials: Associations with Motor Development
9.1. Method
9.1.1. Participants
9.1.2. Materials
9.1.3. Procedure
9.2. Results
9.2.1. Novelty Preferences Compared to Chance
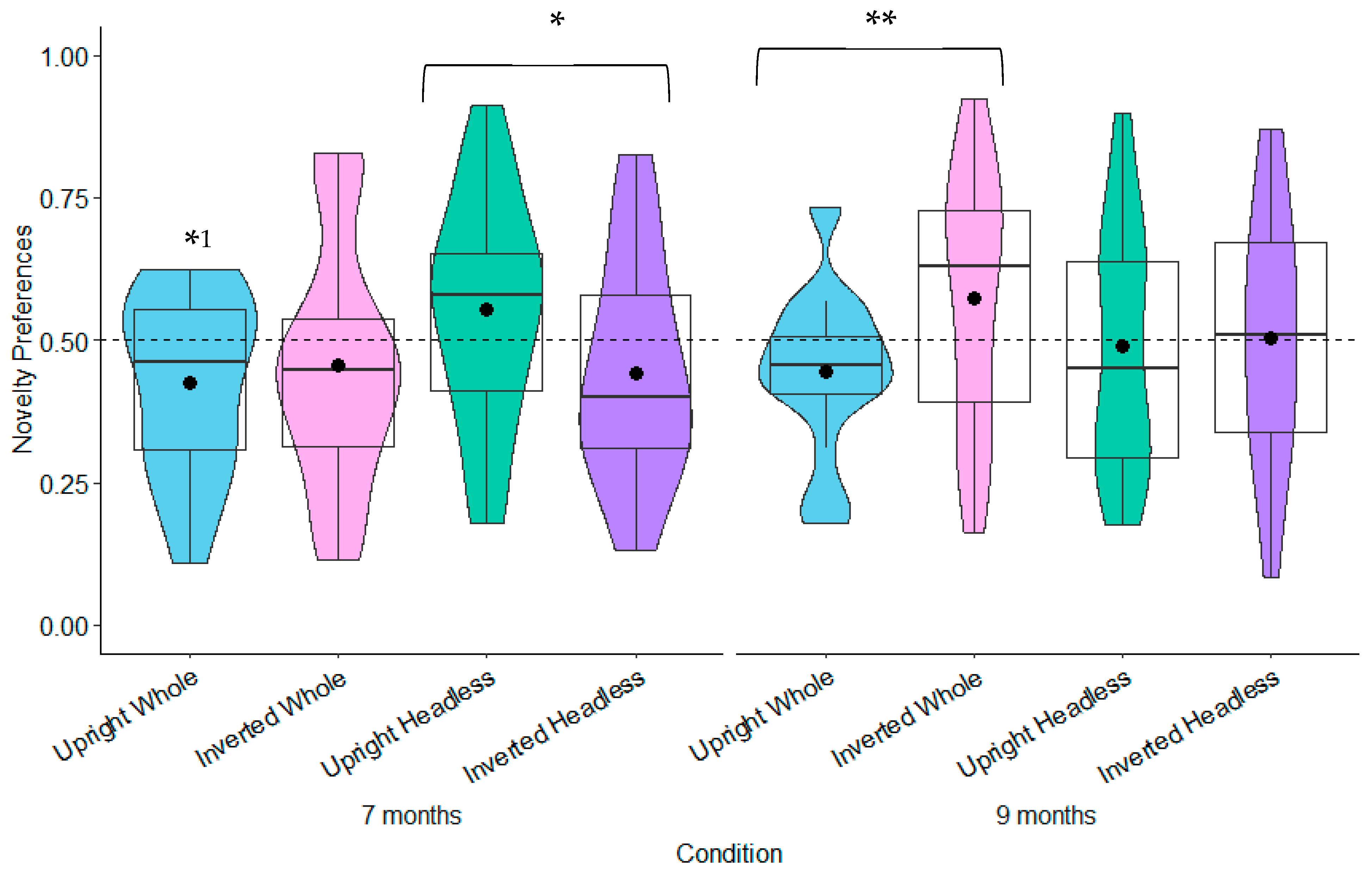
9.2.2. Comparison of Novelty Preferences Across Age, Condition, Test Trial, and Relationships with Gross and Fine Motor Development
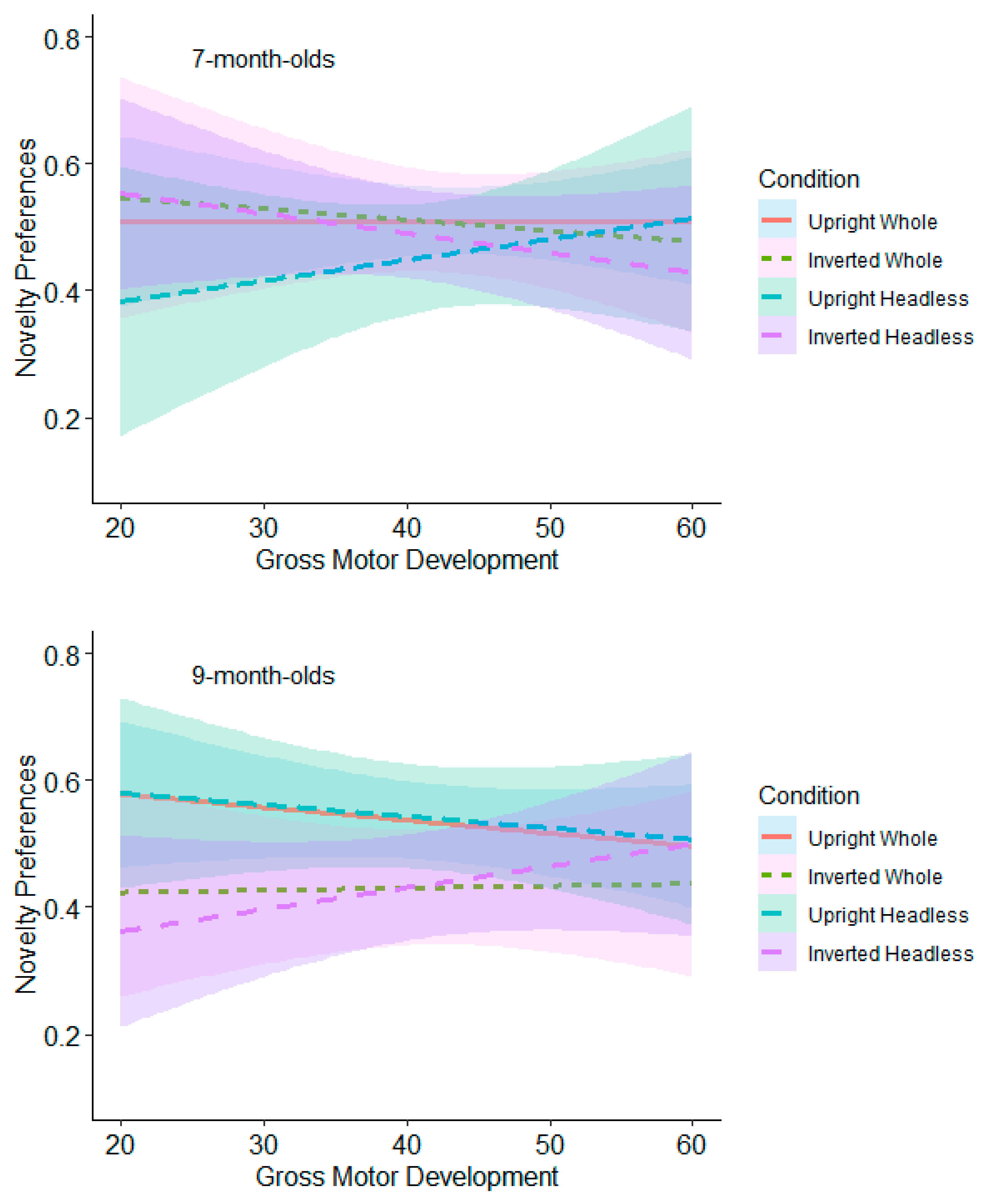
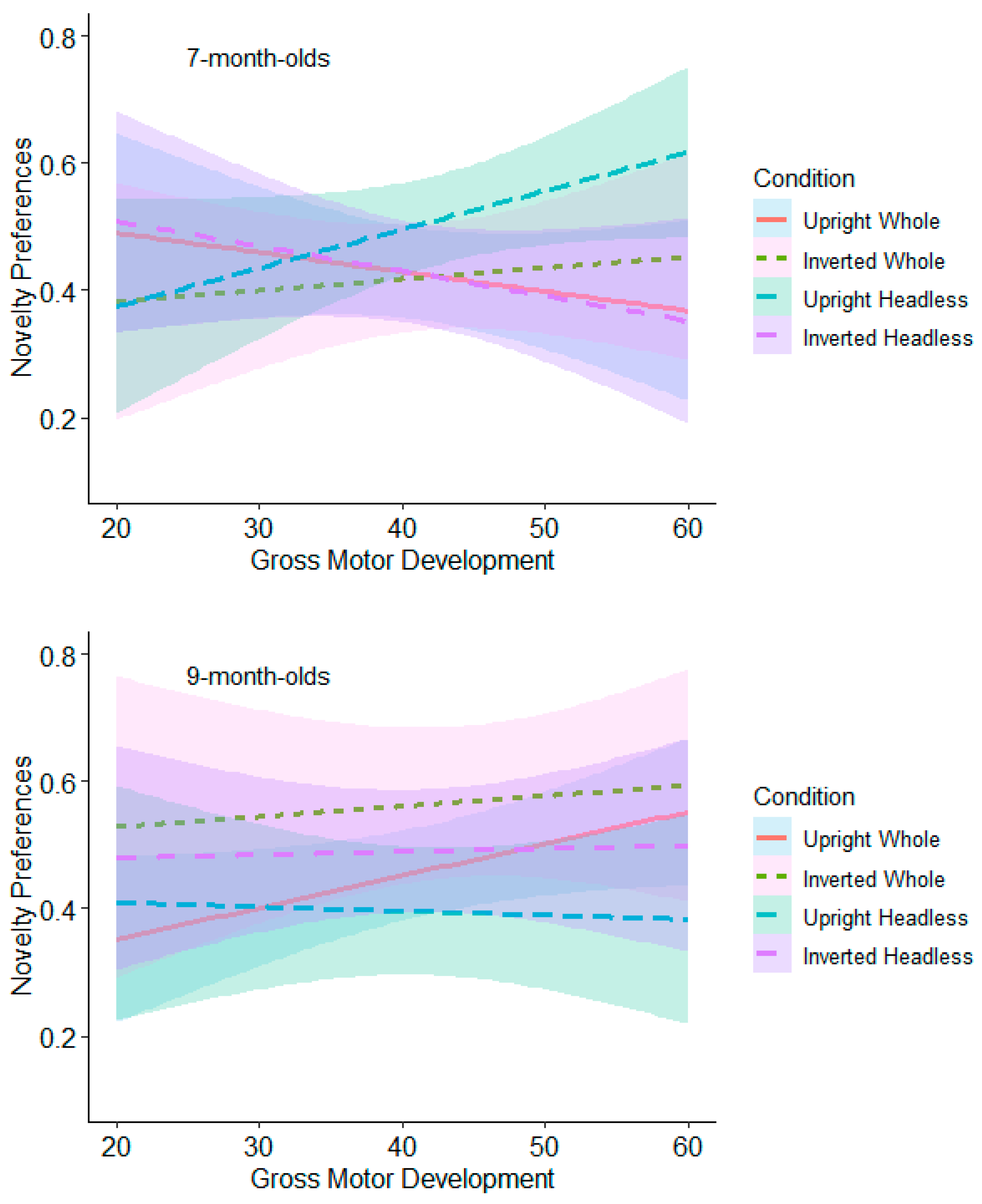
9.2.3. Gross Motor Moderation Analyses
9.2.4. Fine Motor Moderation Analyses
9.3. Discussion
10. General Discussion
10.1. Infants’ Discrimination of Body Postures
10.2. Implications for Infants’ Visuospatial Body Representations
10.3. Relationships with Motor Development
10.4. Implications for Sensorimotor Influences on Infants’ Visuospatial Body Representations
10.5. Limitations and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adolph, K. E., & Hoch, J. E. (2019). Motor development: Embodied, embedded, enculturated, and enabling. Annual Review of Psychology, 70, 141–164. [Google Scholar] [CrossRef]
- Arizpe, J. M., McKean, D. L., Tsao, J. W., & Chan, A. W.-Y. (2017). Where you look matters for body perception: Preferred gaze location contributes to the body inversion effect. PLoS ONE, 12(1), e0169148. [Google Scholar] [CrossRef]
- Axelsson, E. L., Buddhadasa, T., Manca, L., & Robbins, R. A. (2022). Making heads or tails of body inversion effects: Do heads matter? PLoS ONE, 17(2), e0263902. [Google Scholar] [CrossRef]
- Axelsson, E. L., Robbins, R. A., Copeland, H. F., & Covell, H. W. (2019). Body inversion effects with photographic images of body postures: Is it about faces? Frontiers in Psychology, 10, 2686. [Google Scholar] [CrossRef]
- Balas, B. J., Nelson, C. A., Westerlund, A., Vogel-Farley, V., Riggins, T., & Kuefner, D. (2010). Personal familiarity influences the processing of upright and inverted faces in infants [Original Research]. Frontiers in Human Neuroscience, 4, 846. [Google Scholar] [CrossRef]
- Bardi, L., Regolin, L., & Simion, F. (2014). The first time ever I saw your feet: Inversion effect in newborns’ sensitivity to biological motion. Developmental Psychology, 50(4), 986–993. [Google Scholar] [CrossRef]
- Brandman, T., & Yovel, G. (2010). The body inversion effect is mediated by face-selective, not body-selective mechanisms. The Journal of Neuroscience, 30(31), 10534–10540. [Google Scholar] [CrossRef]
- Bruyer, R., & Brysbaert, M. (2011). Combining speed and accuracy in cognitive psychology: Is the inverse efficiency score (IES) a better dependent variable than the mean reaction time (RT) and the percentage of errors (PE)? Psychologica Belgica, 51(1), 5–13. [Google Scholar] [CrossRef]
- Burling, J. M., Kadambi, A., Safari, T., & Lu, H. (2019). The Impact of Autistic Traits on Self-Recognition of Body Movements [Original Research]. Frontiers in Psychology, 9, 2687. [Google Scholar] [CrossRef]
- Burns, Y. R., Ensbey, R. M., & Norrie, M. A. (1989). The neuro-sensory motor developmental assessment part 1: Development and administration of the test. Australian Journal of Physiotherapy, 35(3), 141–149. [Google Scholar] [CrossRef]
- Buxbaum, L. J., & Coslett, H. B. (2001). Specialised structural descriptions for human body parts: Evidence from autotopagnosia. Cognitive Neuropsychology, 18(4), 289–306. [Google Scholar] [CrossRef]
- Caldara, R., & Miellet, S. (2011). iMap: A novel method for statistical fixation mapping of eye movement data. Behavior Research Methods, 43(3), 864–878. [Google Scholar] [CrossRef]
- Cashon, C. H., & Holt, N. A. (2015). Developmental origins of the face inversion effect. Advances in Child Development and Behavior, 48, 117–150. [Google Scholar] [CrossRef]
- Choi, B., Leech, K. A., Tager-Flusberg, H., & Nelson, C. A. (2018). Development of fine motor skills is associated with expressive language outcomes in infants at high and low risk for autism spectrum disorder. Journal of Neurodevelopmental Disorders, 10(1), 14. [Google Scholar] [CrossRef]
- Christie, T., & Slaughter, V. (2009). Exploring links between sensorimotor and visuospatial body representations in infancy. Developmental Neuropsychology, 34(4), 448–460. [Google Scholar] [CrossRef]
- Civile, C., McLaren, R., & McLaren, I. P. (2016). The face inversion effect: Roles of first- and second-order configural information. The American Journal of Psychology, 129(1), 23–35. [Google Scholar] [CrossRef]
- Conte, S., Richards, J. E., Guy, M. W., Xie, W., & Roberts, J. E. (2020). Face-sensitive brain responses in the first year of life. Neuroimage, 211, 116602. [Google Scholar] [CrossRef]
- de Haan, M., Johnson, M. H., & Halit, H. (2003). Development of face-sensitive event-related potentials during infancy: A review. International Journal of Psychophysiology, 51, 45–58. [Google Scholar] [CrossRef]
- de Klerk, C. C. J. M., Filippetti, M. L., & Rigato, S. (2021). The development of body representations: An associative learning account. Proceedings of the Royal Society B: Biological Sciences, 288(1949), 20210070. [Google Scholar] [CrossRef]
- Diamond, R., & Carey, S. (1986). Why faces are not special: An effect of expertise. Journal of Experimental Psychology: General, 115(2), 107–117. [Google Scholar] [CrossRef]
- Fantz, R. L. (1964). Visual experience in infants: Decreased attention to familiar patterns relative to novel ones. Science, 146, 668–670. [Google Scholar] [CrossRef]
- Faul, F., Erdfelder, E., Lang, A., & Buchner, A. (2007). G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods, 39(2), 175–191. [Google Scholar] [CrossRef]
- Fausey, C. M., Jayaraman, S., & Smith, L. B. (2016). From faces to hands: Changing visual input in the first two years. Cognition, 152, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Filippetti, M. L., Johnson, M. H., Lloyd-Fox, S., Dragovic, D., & Farroni, T. (2013). Body perception in newborns. Current Biology, 23(23), 2413–2416. [Google Scholar] [CrossRef]
- Fisher-Thompson, D. (2017). Contributions of look duration and gaze shift patterns to infants’ novelty preferences. Infancy, 22(2), 190–222. [Google Scholar] [CrossRef]
- Fisher-Thompson, D., & Peterson, J. A. (2004). Infant side biases and familiarity-novelty preferences during a serial paired-comparison task. Infancy, 5(3), 309–340. [Google Scholar] [CrossRef]
- Gillmeister, H., Stets, M., Grigorova, M., & Rigato, S. (2019). How do bodies become special? Electrophysiological evidence for the emergence of body-related cortical processing in the first 14 months of life. Developmental Psychology, 55(10), 2025–2038. [Google Scholar] [CrossRef] [PubMed]
- Gliga, T., & Dehaene-Lambertz, G. (2005). Structural encoding of body and face in human infants and adults. Journal of Cognitive Neuroscience, 17(8), 1328–1340. [Google Scholar] [CrossRef]
- Griffin, J. W., & Oswald, F. (2022). A multilevel Bayesian meta-analysis of the body inversion effect: Evaluating controversies over headless and sexualized bodies. Psychonomic Bulletin & Review, 29(5), 1558–1593. [Google Scholar] [CrossRef]
- Heron-Delaney, M., Quinn, P. C., Lee, K., Slater, A. M., & Pascalis, O. (2013). Nine-month-old infants prefer unattractive bodies over attractive bodies. Journal of Experimental Child Psychology, 115, 30–41. [Google Scholar] [CrossRef]
- Hock, A., White, H., Jubran, R., & Bhatt, R. S. (2016). The whole picture: Holistic body posture recognition in infancy. Psychonomic Bulletin and Review, 23(2), 426–431. [Google Scholar] [CrossRef]
- Houston-Price, C., & Nakai, S. (2004). Distinguishing novelty and familiarity effects in infant preference procedures. Infant and Child Development, 13, 341–348. [Google Scholar] [CrossRef]
- Hunter, M. A., & Ames, E. W. (1988). A multifactor model of infant preferences for novel and familiar stimuli. In Advances in infancy research (Vol. 5, pp. 69–95). Ablex Publishing. [Google Scholar]
- Johnson, M. H., Dziurawiec, S., Ellis, H., & Morton, J. (1991). Newborns’ preferential tracking of face-like stimuli and its subsequent decline. Cognition, 40(1–2), 1–19. [Google Scholar] [CrossRef]
- Jones, E. J. H., & Herbert, J. S. (2009). Imitation and the development of infant learning, memory, and categorisation. Revue de Primatologie, 1(4). [Google Scholar] [CrossRef]
- Key, A. P. F., & Stone, W. L. (2012). Processing of novel and familiar faces in infants at average and high risk for autism. Developmental Cognitive Neuroscience, 2(2), 244–255. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M., Kanazawa, S., Yamaguchi, M. K., & O’Toole, A. J. (2021). Cortical processing of dynamic bodies in the superior occipito-temporal regions of the infants’ brain: Difference from dynamic faces and inversion effect. Neuroimage, 244, 118598. [Google Scholar] [CrossRef]
- Kosakowski, H. L., Cohen, M. A., Takahashi, A., Keil, B., Kanwisher, N., & Saxe, R. (2022). Selective responses to faces, scenes, and bodies in the ventral visual pathway of infants. Current Biology, 32(2), 265–274.e265. [Google Scholar] [CrossRef]
- Lao, J., Miellet, S., Pernet, C., Sokhn, N., & Caldara, R. (2017). iMap4: An open source toolbox for the statistical fixation mapping of eye movement data with linear mixed modeling. Behavior Research Methods, 49(2), 559–575. [Google Scholar] [CrossRef]
- Marks, K. P., & LaRosa, A. C. (2012). Understanding developmental-behavioral screening measures. Pediatrics in Review, 33(10), 448–457; quiz 457–448. [Google Scholar] [CrossRef]
- Minnebusch, D. A., Suchan, B., & Daum, I. (2009). Losing your head: Behavioral and electrophysiological effects of body inversion. Journal of Cognitive Neuroscience, 21(5), 865–874. [Google Scholar] [CrossRef]
- Oakes, L. M. (2023a). The cascading development of visual attention in infancy: Learning to look and looking to learn. Current Directions in Psychological Science, 32(5), 410–417. [Google Scholar] [CrossRef]
- Oakes, L. M. (2023b). Understanding developmental cascades and experience: Diversity matters. Infancy, 28(3), 492–506. [Google Scholar] [CrossRef]
- Otsuka, Y., Nakato, E., Kanazawa, S., Yamaguchi, M. K., Watanabe, S., & Kakigi, R. (2007). Neural activation to upright and inverted faces in infants measured by near infrared spectroscopy. Neuroimage, 34(1), 399–406. [Google Scholar] [CrossRef]
- Peelen, M. V., & Downing, P. E. (2007). The neural basis of visual body perception. Nature Reviews Neuroscience, 8, 636–648. [Google Scholar] [CrossRef]
- Piepers, D., & Robbins, R. (2012). A review and clarification of the terms “holistic,” “configural,” and “relational” in the face perception literature [review]. Frontiers in Psychology, 3, 559. [Google Scholar] [CrossRef]
- Piper, M. C., & Darrah, J. (1994). Motor assessment of the developing infant. W.B. Saunders Company. [Google Scholar]
- Pitcher, D., & Ungerleider, L. G. (2021). Evidence for a Third Visual Pathway Specialized for Social Perception. Trends in Cognitive Sciences, 25(2), 100–110. [Google Scholar] [CrossRef]
- Quinn, P. C., & Eimas, P. D. (1998). Evidence for a global categorical representation of humans by young infants. Journal of Experimental Child Psychology, 69(3), 151–174. [Google Scholar] [CrossRef]
- Reed, C. L., Nyberg, A. A., & Grubb, J. D. (2012). Contributions of visual and embodied expertise to body perception. Perception, 41(4), 436–446. [Google Scholar] [CrossRef]
- Reed, C. L., Stone, V. E., Bozova, S., & Tanaka, J. (2003). The body-inversion effect. Psychological Science, 14(4), 302–308. [Google Scholar] [CrossRef]
- Reed, C. L., Stone, V. E., Grubb, J. D., & McGoldrick, J. E. (2006). Turning configural processing upside down: Part and whole body postures. Journal of Experimental Psychology-Human Perception and Performance, 32(1), 73–87. [Google Scholar] [CrossRef]
- Reid, V. M., Belsky, J., & Johnson, M. H. (2005). Infant perception of human action: Toward a developmental cognitive neuroscience of individual differences. Cognition, Brain, Behavior, 9(2), 35–52. [Google Scholar]
- Reid, V. M., Kaduk, K., & Lunn, J. (2019). Links between action perception and action production in 10-week-old infants. Neuropsychologia, 126, 69–74. [Google Scholar] [CrossRef]
- Rigato, S., Filippetti, M. L., & de Klerk, C. (2023). Infants’ representations of the infant body in the first year of life: A preferential looking time study. Scientific Reports, 13(1), 14091. [Google Scholar] [CrossRef]
- Robbins, R. A., & Coltheart, M. (2012). The effects of inversion and familiarity on face versus body cues to person recognition. Journal of Experimental Psychology: Human Perception and Performance, 38(5), 1098–1104. [Google Scholar] [CrossRef]
- Ropar, D., Greenfield, K., Smith, A. D., Carey, M., & Newport, R. (2018). Body representation difficulties in children and adolescents with autism may be due to delayed development of visuo-tactile temporal binding. Developmental Cognitive Neuroscience, 29, 78–85. [Google Scholar] [CrossRef]
- Rose, S. A., Jankowski, J. J., & Feldman, J. F. (2008). The inversion effect in infancy: The role of internal and external features. Infant Behavior & Development, 31(3), 470–480. [Google Scholar] [CrossRef]
- Rubio-Codina, M., Araujo, M. C., Attanasio, O., Muñoz, P., & Grantham-McGregor, S. (2016). Concurrent validity and feasibility of short tests currently used to measure early childhood development in large scale studies. PLoS ONE, 11(8), e0160962. [Google Scholar] [CrossRef]
- Simion, F., & Giorgio, E. D. (2015). Face perception and processing in early infancy: Inborn predispositions and developmental changes. Frontiers in Psychology, 6, 969. [Google Scholar] [CrossRef]
- Sirigu, A., Grafman, J., Bressler, K., & Sunderland, T. (1991). Multiple representations contribute to body knowledge processing. Brain, 114, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Slater, A., Bremner, G., Johnson, S. P., Sherwood, P., Hayes, R., & Brown, E. (2000). Newborn infants’ preference for attractive faces: The role of internal and external facial features. Infancy, 1(2), 265–274. [Google Scholar] [CrossRef] [PubMed]
- Slaughter, V., Heron-Delaney, M., & Christie, T. (2012). Developing expertise in human body perception. In V. Slaughter, & C. A. Brownell (Eds.), Early development of body representations (pp. 81–100). Cambridge University Press. [Google Scholar] [CrossRef]
- Slaughter, V., Stone, V. E., & Reed, C. (2004). Perception of faces and bodies. Similar or different? Current Directions in Psychological Science, 13(6), 219–223. [Google Scholar] [CrossRef]
- Sommerville, J. A., Woodward, A. L., & Needham, A. (2005). Action experience alters 3-month-old infants’ perception of others’ actions. Cognition, 96(1), B1–B11. [Google Scholar] [CrossRef] [PubMed]
- Squires, J., Bricker, D. D., Twombly, E., & Potter, L. (2009). Ages & stages questionnaires: A parent-completed, child-monitoring system (3rd ed.). Brookes Pub. [Google Scholar]
- Thompson, L. A., Madrid, V., Westbrook, S., & Johnston, V. (2001). Infants attend to second-order relational properties of faces. Psychonomic Bulletin & Review, 8(4), 769–777. [Google Scholar] [CrossRef] [PubMed]
- Thurman, S. L., & Corbetta, D. (2019). Changes in posture and interactive behaviors as infants progress from sitting to walking: A longitudinal study [original research]. Frontiers in Psychology, 10, 822. [Google Scholar] [CrossRef]
- Turati, C., Sangrigoli, S., Ruel, J., & de Schonen, S. (2004). Evidence of the face inversion effect in 4-month-old infants. Infancy, 6(2), 275–297. [Google Scholar] [CrossRef]
- Woodward, A. L., & Guajardo, J. J. (2002). Infants’ understanding of the point gesture as an object-directed action. Cognitive Development, 17(1), 1061–1084. [Google Scholar] [CrossRef]
- Xie, S., Hoehl, S., Moeskops, M., Kayhan, E., Kliesch, C., Turtleton, B., Köster, M., & Cichy, R. M. (2022). Visual category representations in the infant brain. Current Biology, 32(24), 5422–5432.e5426. [Google Scholar] [CrossRef]
- Yin, R. K. (1969). Looking at upside-down faces. Journal of Experimental Psychology, 81, 141–145. [Google Scholar] [CrossRef]
- Yovel, G., Pelc, T., & Lubetzky, I. (2010). It’s all in your head: Why is the body inversion effect abolished for headless bodies? Journal of Experimental Psychology: Human Perception and Performance, 36(3), 759–767. [Google Scholar] [CrossRef]
- Yu, C., & Smith, L. B. (2012). Embodied attention and word learning by toddlers. Cognition, 125(2), 244–262. [Google Scholar] [CrossRef]
- Zieber, N., Bhatt, R. S., Hayden, A., Kangas, A., Collins, R., & Bada, H. (2010). Body representation in the first year of life. Infancy, 15(5), 534–544. [Google Scholar] [CrossRef] [PubMed]
- Zieber, N., Kangas, A., Hock, A., & Bhatt, R. S. (2014). Infants’ perception of emotion from body movements. Child Development, 85(2), 675–684. [Google Scholar] [CrossRef] [PubMed]
- Zieber, N., Kangas, A., Hock, A., & Bhatt, R. S. (2015). Body structure perception in infancy. Infancy, 20(1), 1–17. [Google Scholar] [CrossRef]
- Zimmermann, M., Toni, I., & de Lange, F. P. (2013). Body posture modulates action perception. Journal of Neuroscience, 33(14), 5930–5938. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Axelsson, E.L.; Britton, T.; Gulhati, G.K.; Kelly, C.; Copeland, H.; McNamara, L.; Covell, H.; Quinn, A.A. Strike a Pose: Relationships Between Infants’ Motor Development and Visuospatial Representations of Bodies. Behav. Sci. 2025, 15, 1021. https://doi.org/10.3390/bs15081021
Axelsson EL, Britton T, Gulhati GK, Kelly C, Copeland H, McNamara L, Covell H, Quinn AA. Strike a Pose: Relationships Between Infants’ Motor Development and Visuospatial Representations of Bodies. Behavioral Sciences. 2025; 15(8):1021. https://doi.org/10.3390/bs15081021
Chicago/Turabian StyleAxelsson, Emma L., Tayla Britton, Gurmeher K. Gulhati, Chloe Kelly, Helen Copeland, Luca McNamara, Hester Covell, and Alyssa A. Quinn. 2025. "Strike a Pose: Relationships Between Infants’ Motor Development and Visuospatial Representations of Bodies" Behavioral Sciences 15, no. 8: 1021. https://doi.org/10.3390/bs15081021
APA StyleAxelsson, E. L., Britton, T., Gulhati, G. K., Kelly, C., Copeland, H., McNamara, L., Covell, H., & Quinn, A. A. (2025). Strike a Pose: Relationships Between Infants’ Motor Development and Visuospatial Representations of Bodies. Behavioral Sciences, 15(8), 1021. https://doi.org/10.3390/bs15081021