

Rest to Promote Learning: A Brain Default Mode Network Perspective


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
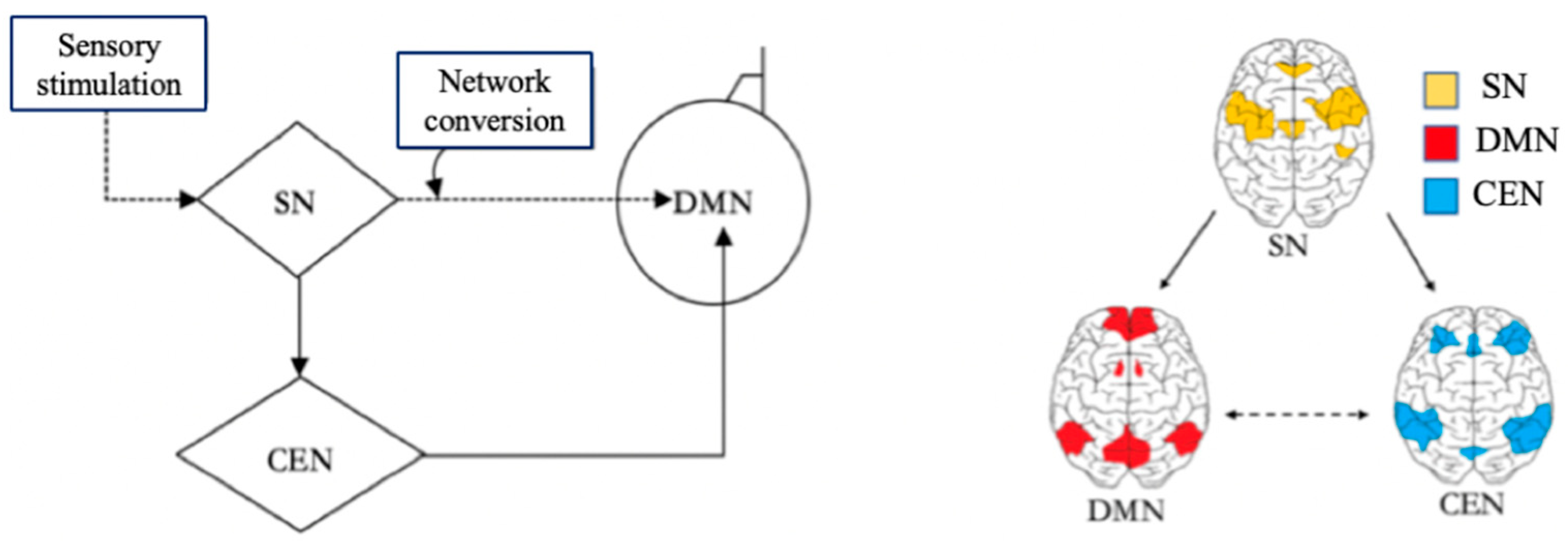
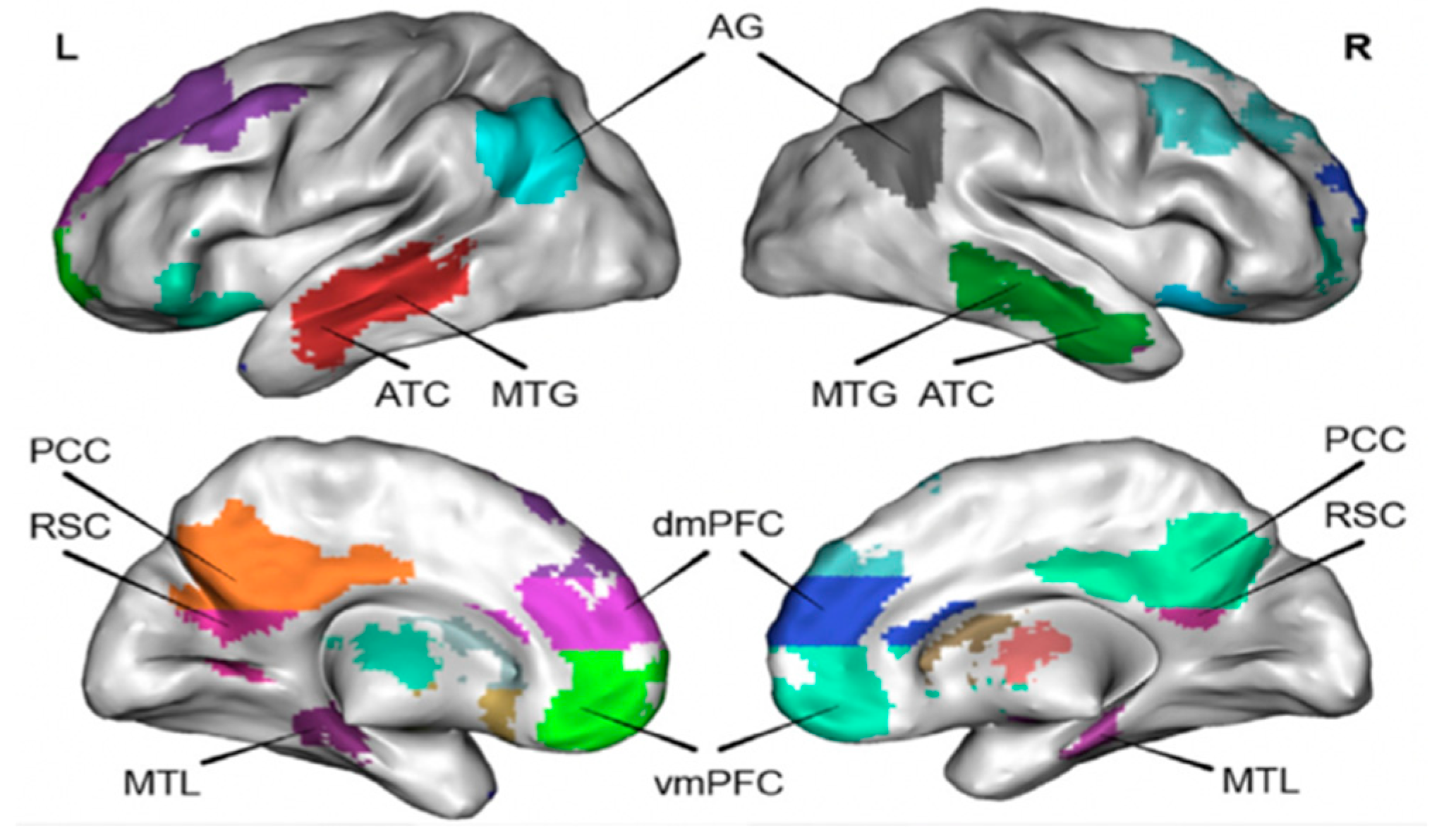
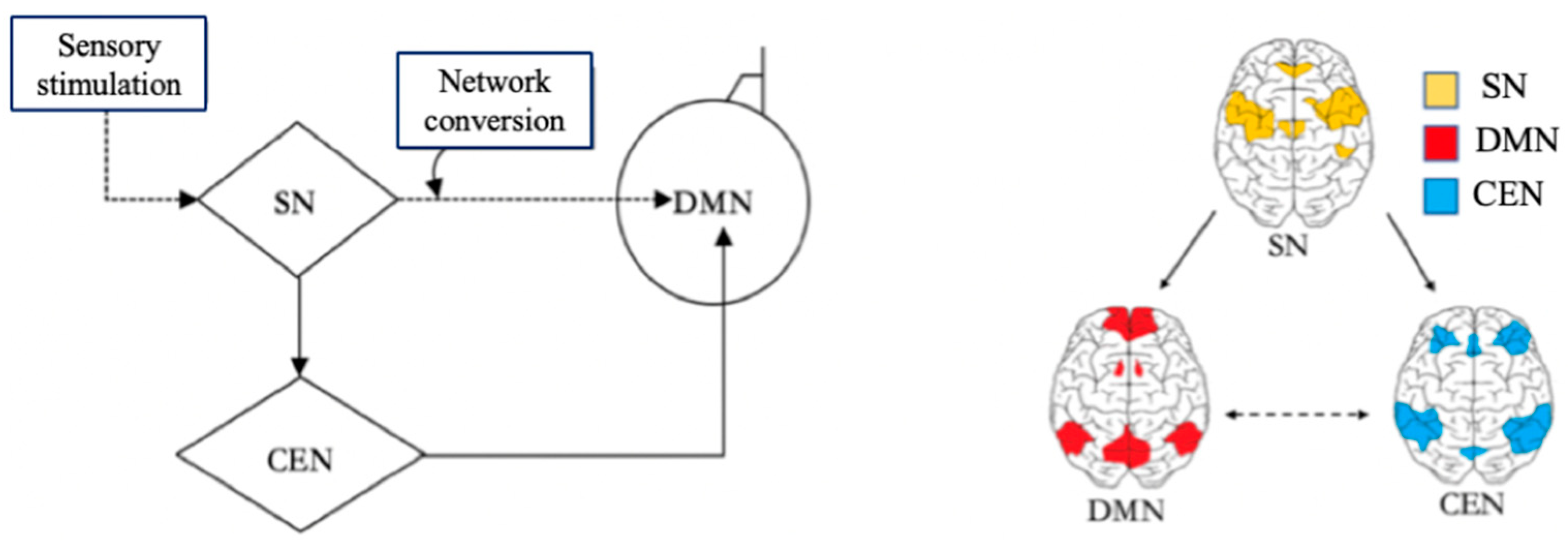
2. The Role of DMN in Learning
3. The DMN’s Functions in Learning
4. Learning-Related DMN Characteristics
4.1. DMN’s High Energy Consumption to Meet the Needs of Learning
4.2. Automatic Adjustment of DMN Activation in Accordance with the Difficulty of Learning Tasks
4.3. DMN’s “Inverted U” Shape with Age: The Close Connection with Learning Activities at Different Ages
4.4. DMN, Mental Illness and Learning: Mental Illness and Alteration in DMN Affect Each Other and Ultimately Affect Learning
5. Neural Activity Occurring in Default Mode Facilitates Learning
5.1. The Brain’s Frequent Use of Beta Waves for Rhythmic Regulation during Rest
5.2. The Brain’s Spontaneous Initiation of the “Subconscious” Divergence Mode during Rest
5.3. The Hippocampus’ Full Engagement in Information Integration during Rest
5.4. Recurrent Neural Replay Occurring in the Brain during Rest
6. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mayer, R.E. How Can Brain Research Inform Academic Learning and Instruction. Educ. Psychol. Rev. 2017, 29, 835–846. [Google Scholar] [CrossRef]
- Buckner, R.L.; Andrews-Hanna, J.R.; Schacter, D.L. The brain’s default network: Anatomy, function and relevance to disease. Ann. N. Y. Acad. Sci. 2008, 1124, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Menon, V. 20 years of the default mode network: A review and synthesis. Neuron 2023, 111, 2469–2487. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.S.; Zheng, Y.; Dong, G.-H.; Glassman, H.; Huang, C.; Xuan, R. Resting state default mode network is associated with wise advising. Sci. Rep. 2023, 13, 14239. [Google Scholar] [CrossRef] [PubMed]
- Ericsson, K.A.; Krampe, R.T.; Tesch-Römer, C. The role of deliberate practice in the acquisition of expert performance. Psychol. Rev. 1993, 100, 363–406. [Google Scholar] [CrossRef]
- Buch, E.R.; Claudino, L.; Quentin, R.; Bönstrup, M.; Cohen, L.G. Consolidation of human skill linked to waking hippocampo-neocortical replay. Cell Rep. 2021, 35, 109193. [Google Scholar] [CrossRef] [PubMed]
- Buckner, R.L.; Carroll, D.C.; Buckner, R.L.; Carroll, D.C. Self-projection and the brain. Trends Cogn. Sci. 2007, 11, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Li, S.; Zhuo, C.; Li, M.; Safron, A.; Genz, A.; Qin, W.; Yu, C.; Walter, M. Altered task specific deactivation in the default mode network depends on valence in patients with major depressive disorder. J. Affect. Disord. 2016, 207, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Jolliffe, M.; Lograsso, Y.S.S.; Bearden, C.E. A Review of Default Mode Network Connectivity and Its Association with Social Cognition in Adolescents with Autism Spectrum Disorder and Early-Onset Psychosis. Front. Psychiatry 2020, 11, 614. [Google Scholar] [CrossRef]
- Sachdev, P.S. The default mode network, depression and Alzheimer’s disease. Int. Psychogeriatr. 2022, 34, 675–678. [Google Scholar] [CrossRef]
- Raichle, M.E. The brain’s default mode network. Annu. Rev. Neurosci. 2015, 38, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Andrews-Hanna, J.R. The Brain’s Default Network and its Adaptive Role in Internal Mentation. Neuroscientist 2012, 18, 251–270. [Google Scholar] [CrossRef] [PubMed]
- Howard-Jones, P.A.; Jay, T.; Mason, A.; Jones, H. Gamification of Learning Deactivates the Default Mode Network. Front. Psychol. 2016, 6, 162828. [Google Scholar] [CrossRef] [PubMed]
- Mangold, R.; Sokoloff, L.; Conner, E.; Kleinerman, J.; Therman, P.-O.G.; Kety, S.S. The effects of sleep and lack of sleep on the cerebral circulation and metabolism of normal young men. J. Clin. Investig. 1955, 34, 1092–1100. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.E.; Snyder, A.Z. A default mode of brain function: A brief history of an evolving idea. NeuroImage 2007, 37, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Zhou, P.; Alcauter, S.; Chen, Y.; Cao, H.; Tian, M.; Ming, D.; Qi, H.; Wang, X.; Zhao, X.; et al. Changes of intranetwork and internetwork functional connectivity in alzheimer’s disease and mild cognitive impairment. J. Neural Eng. 2016, 13, 046008. [Google Scholar] [CrossRef] [PubMed]
- Shulman, G.L.; Fiez, J.A.; Corbetta, M.; Buckner, R.L.; Miezin, F.M.; Raichle, M.E.; Petersen, S.E. Common blood flow changes across visual tasks. II. Decreases in cerebral cortex. J. Cogn. Neurosci. 1997, 9, 648–663. [Google Scholar] [CrossRef]
- Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Corbetta, M.; van Essen, D.C.; Raichle, M.E. The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc. Natl. Acad. Sci. USA 2005, 102, 9673–9678. [Google Scholar] [CrossRef]
- Greicius, M.D.; Krasnow, B.; Reiss, A.L.; Menon, V. Functional Connectivity in the Resting Brain: A Network Analysis of the Default Mode Hypothesis. Proc. Natl. Acad. Sci. USA 2003, 100, 253–258. [Google Scholar] [CrossRef]
- Foster, B.L.; Dastjerdi, M.; Parvizi, J. Neural populations in human posteromedial cortex display opposing responses during memory and numerical processing. Proc. Natl. Acad. Sci. USA 2012, 109, 15514–15519. [Google Scholar] [CrossRef] [PubMed]
- Buckner, R.L.; Snyder, A.Z.; Shannon, B.J.; LaRossa, G.; Sachs, R.; Fotenos, A.F.; Mintun, M.A. Molecular. Structural and functional characterization of Alzheimer’s disease: Evidence for a relationship between default activity, amyloid and memory. J. Neurosci. 2005, 25, 7709–7717. [Google Scholar] [CrossRef] [PubMed]
- Mason, M.F.; Norton, M.I.; Van Horn, J.D.; Wegner, D.M.; Grafton, S.T.; Macrae, C.N. Wandering minds: The default network and stimulus-independent thought. Science 2007, 315, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Thomas Yeo, B.T.; Krienen, F.M.; Sepulcre, J.; Sabuncu, M.R.; Lashkari, D.; Hollinshead, M.; Roffman, J.L.; Smoller, J.W.; Zöllei, L.; Polimeni, J.R.; et al. The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J. Neurophysiol. 2011, 106, 1125–1165. [Google Scholar] [CrossRef] [PubMed]
- Peters, S.K.; Dunlop, K.; Downar, J. Cortico-Striatal-Thalamic Loop Circuits of the Salience Network: A Central Pathway in Psychiatric Disease and Treatment. Front. Syst. Neurosci. 2016, 10, 104. [Google Scholar] [CrossRef] [PubMed]
- Menon, V. Large-scale brain networks and psychopathology: A unifying triple network model. Trends Cogn. Sci. 2011, 15, 483–506. [Google Scholar] [CrossRef] [PubMed]
- Monto, S.; Palva, S.; Voipio, J.; Palva, J.M. Very slow EEG fluctuations predict the dynamics of stimulus detection and oscillation amplitudes in humans. J. Neurosci. 2008, 28, 8268–8272. [Google Scholar] [CrossRef]
- Fox, M.D.; Raichile, M.E. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat. Rev. Neurosci. 2007, 8, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.F.; He, Y.; Zhu, C.Z.; Cao, Q.J.; Sui, M.Q.; Liang, M.; Wang, Y.F. Altered baseline brain activity in children with ADHD revealed by resting-state functional MRI. Brain Dev. 2007, 29, 83–91. [Google Scholar]
- Zang, Y.F.; Jiang, T.Z.; Lu, Y.L.; He, Y.; Tian, L.X. Regional homogeneity approach to fMRI data analysis. NeuroImage 2004, 22, 394–400. [Google Scholar] [CrossRef]
- Long, X.Y.; Zuo, X.N.; Kiviniemi, V.; Yang, Y.H.; Zou, Q.H.; Zhu, C.Z.; Zang, Y.F. Default mode network as revealed with multiple methods for resting-state functional MRI analysis. J. Neurosci. Methods 2008, 171, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q.H.; Zhu, C.Z.; Yang, Y.H.; Zuo, X.N.; Long, X.Y.; Cao, Q.J.; Zang, Y.F. An improved approach to detection of amplitude of low-frequency fluctuation (ALFF) for resting-state fMRI: Fractional ALFF. J. Neurosci. Methods 2008, 172, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Supekar, K.; Uddin, L.Q.; Prater, K.; Amin, H.; Greicius, M.D.; Menon, V. Development of functional and structural connectivity within the default mode network in young children. NeuroImage 2010, 52, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Meunier, D.; Achard, S.; Morcom, A.; Bullmore, E. Age-related changes in modular organization of human brain functional networks. NeuroImage 2009, 44, 715–723. [Google Scholar] [CrossRef]
- Fransson, P.; Marrelec, G. The precuneus/posterior cingulate cortex plays a pivotal role in the default mode network: Evidence from a partial correlation network analysis. NeuroImage 2008, 42, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Fair, D.A.; Cohen, A.L.; Dosenbach, N.U.; Church, J.A.; Miezin, F.M.; Barch, D.M.; Schlaggar, B.L. The maturing architecture of the brain’s default network. Proc. Natl. Acad. Sci. USA 2008, 105, 4028–4032. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Zhu, H.T.; Giovanello, K.S.; Smith, J.K.; Shen, D.G.; Gilmore, J.H.; Lin, W.L. Evidence on the emergence of the brain’s default network from 2-week-old to 2-year-old healthy pediatric subjects. Proc. Natl. Acad. Sci. USA 2009, 106, 6790–6795. [Google Scholar] [CrossRef] [PubMed]
- Boly, M.; Tshibanda, L.; Vanhaudenhuyse, A.; Noirhomme, Q.; Schnakers, C.; Ledoux, D.; Laureys, S. Functional connectivity in the default network during resting state is preserved in a vegetative but not in a brain dead patient. Hum. Brain Mapp. 2009, 30, 2393–2400. [Google Scholar] [CrossRef] [PubMed]
- Thomason, M.E.; Chang, C.E.; Glover, G.H.; Gabrieli, J.D.; Greicius, M.D.; Gotlib, I.H. Default mode function and task induced deactivation have overlapping brain substrates in children. NeuroImage 2008, 41, 1493–1503. [Google Scholar] [CrossRef]
- Marsh, R.; Zhu, H.; Schultz, R.T.; Quackenbush, G.; Royal, J.; Skudlarski, P.; Peterson, B.S.A. Developmental fMRI study of self-regulatory control. Hum. Brain Mapp. 2006, 27, 848–863. [Google Scholar] [CrossRef]
- Damoiseaux, J.; Beckmann, C.; Arigita, E.S.; Barkhof, F.; Scheltens, P.; Stam, C.; Smith, S.; Rombouts, S. Reduced resting-state brain activity in the “default network” in normal aging. Cereb Cortex. 2008, 18, 1856–1864. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, M.C.; Rosen, H.J.; Miller, B.L. Neuroimaging in Dementia; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Wu, J.-T.; Wu, H.-Z.; Yan, C.-G.; Chen, W.-X.; Zhang, H.-Y.; He, Y.; Yang, H.-S. Aging-related changes in the default mode network and its anticorrelated networks: A resting-state fMRI study. Neurosci. Lett. 2011, 504, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Bi, Y.; Chen, W.; Chen, H.; Yang, Z.; Chen, H.; Zhang, Y. Resting default network study on mild cognitive impairment in cerebral white matter osteoporosis. Chin. Rehabil. Theory Pract. 2014, 20, 1133–1139. [Google Scholar]
- Greicius, M.D.; Srivastava, G.; Reiss, A.L.; Menon, V. Default mode network activity distinguishes Alzheimer’s disease from healthy aging: Evidence from functional MRI. Proc. Natl. Acad. Sci. USA 2004, 101, 4637–4642. [Google Scholar] [CrossRef]
- Greicius, M. Resting-state functional connectivity in neuropsychiatric disorders. Curr. Opin. Neurol. 2008, 21, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Broyd, S.J.; Demanuele, C.; Debener, S.; Helps, S.K.; James, C.J.; Sonuga-Barke, E.J. Default-mode brain dysfunction in mental disorders: A systematic review. Neurosci. Biobehav. Rev. 2009, 33, 279–296. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Jiang, T.; Wang, Y.; Zang, Y.; He, Y.; Liang, M.; Sui, M.; Cao, Q.; Hu, S.; Peng, M.; et al. Altered resting-state functional connectivity patterns of anterior cingulate cortex in adolescents with attention deficit hyperactivity disorder. Neurosci. Lett. 2006, 400, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Cao, Q.; Long, X.; Sui, M.; Cao, X.; Zhu, C.; Zuo, X.; An, L.; Song, Y.; Zang, Y.; et al. Abnormal functional connectivity between the anterior cingulate and the default mode network in drugnaive boys with attention deficit hyperactivity disorder. Psychiatry Res. 2012, 201, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Uddin, L.Q.; Kelly, A.C.; Biswal, B.B.; Margulies, D.S.; Shehzad, Z.; Shaw, D.; Ghaffari, M.; Rotrosen, J.; Adler, L.A.; Castellanos, F.X.; et al. A Network homogeneity reveals decreased integrity of default mode network in ADHD. J. Neurosci. Methods 2008, 169, 249–254. [Google Scholar] [CrossRef]
- Greicius, M.D.; Flores, B.H.; Menon, V.; Glover, G.H.; Solvason, H.B.; Kenna, H.; Reiss, A.L.; Schatzberg, A.F. Resting-state functional connectivity in major depression: Abnormally increased contributions from subgenual cingulate cortex and thalamus. Biol. Psychiatry 2007, 62, 429–437. [Google Scholar] [CrossRef]
- Wichers, M.; Geschwind, N.; Van, O.J.; Peeters, F. Scars in depression: Is a conceptual shift necessary to solve the puzzle. Psychol. Med. 2010, 40, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liu, H.; Sun, H.; Zhang, J.; Shi, S. Resting fMRI study of first episode depression patients before and after treatment. Chin. J. Med. Comput. Imaging 2011, 17, 212–216. [Google Scholar]
- Zhu, X.; Wang, X.; Xiao, J. A study on the resting default network of first ever untreated depression. Chin. J. Clin. Psychol. 2011, 34, 146–148. [Google Scholar]
- Kennedy, D.P.; Courchesne, E. The intrinsic functional organisation of the brain is altered in autism. NeuroImage 2008, 39, 1877–1885. [Google Scholar] [CrossRef] [PubMed]
- Washington, S.D.; Gordon, E.M.; Brar, J.; Warburton, S.; Sawyer, A.T.; Wolfe, A.; Vanmeter, J.W. Dysmaturation of the default mode network in autism. Hum. Brain Mapp. 2014, 35, 1284–1296. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi, Y.; Harada, M.; Otsuka, H.; Mori, K.; Ito, H.; Iwanaga, T. Default mode network abnormalities in children with autism spectrum disorder detected by resting state functional magnetic resonance imaging. J. Med. Investig. 2016, 63, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Shukla, D.K.; Keehn, B.; Müller, R.A. Regional homogeneity of fMRI time series in autism spectrum disorders. Neurosci. Lett. 2010, 476, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Buckner, R.L. The brain’s default network: Origins and implications for the study of psychosis. Dialogues Clin. Neurosci. 2013, 15, 351–358. [Google Scholar] [CrossRef]
- Mak, L.E.; Minuzzi, L.; MacQueen, G.; Hall, G.; Kennedy, S.H.; Milev, R. The Default Mode Network in Healthy Individuals: A Systematic Review and Meta-Analysis. Brain Connect. 2017, 7, 25–33. [Google Scholar] [CrossRef]
- Matthew, L.; Jia, Y. Social Nature: Three Driving Forces of Human Socialism; People’s Publishing House: Hangzhou, China, 2016; p. 100. [Google Scholar]
- Available online: https://www.themuse.com/advice/the-rule-of-52-and-17-its-random-but-it-ups-your-productivity (accessed on 19 June 2020).
- Nittono, H.; Fukushima, M.; Yano, A.; Moriya, H. The power of kawaii: Viewing cute images promotes a careful behavior and narrows attentional focus. PLoS ONE 2012, 7, e46362. [Google Scholar] [CrossRef]
- Available online: https://qz.com/267823/the-perfect-recipe-for-productivity-rest-17-minutes-every-52/ (accessed on 18 September 2014).
- Available online: http://news.cornell.edu/stories/1999/09/onscreen-break-reminder-boosts-productivity (accessed on 23 September 1999).
- Bönstrup, M.; Iturrate, I.; Thompson, R.; Cruciani, G.; Censor, N.; Cohen, L.G. A Rapid Form of Offline Consolidation in Skill Learning. Curr. Biol. 2019, 29, 1346–1351. [Google Scholar] [CrossRef] [PubMed]
- Josh, W. The Way to Learning; Su, H.; Xie, J., Translators; China Youth Publishing House: Beijing, China, 2016; p. 138. [Google Scholar]
- Pfeiffer, B.E. The content of hippocampal “replay”. Hippocampus 2020, 30, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mattar, M.G.; Behrens, T.E.J.; Daw, N.D.; Dolan, R.J. Experience replay is associated with efficient nonlocal learning. Science 2021, 372, eabf1357. [Google Scholar] [CrossRef] [PubMed]
- Eichenlaub, J.B.; Jarosiewicz, B.; Saab, J.; Franco, B.; Kelemen, J.; Halgren, E.; Cash, S.S. Replay of Learned Neural Firing Sequences during Rest in Human Motor Cortex. Cell Rep. 2020, 31, 107581. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, W.; Liu, B.; Tang, Y.; Huang, J.; Wu, J. Rest to Promote Learning: A Brain Default Mode Network Perspective. Behav. Sci. 2024, 14, 349. https://doi.org/10.3390/bs14040349
Luo W, Liu B, Tang Y, Huang J, Wu J. Rest to Promote Learning: A Brain Default Mode Network Perspective. Behavioral Sciences. 2024; 14(4):349. https://doi.org/10.3390/bs14040349
Chicago/Turabian StyleLuo, Wei, Biao Liu, Ying Tang, Jingwen Huang, and Ji Wu. 2024. "Rest to Promote Learning: A Brain Default Mode Network Perspective" Behavioral Sciences 14, no. 4: 349. https://doi.org/10.3390/bs14040349
APA StyleLuo, W., Liu, B., Tang, Y., Huang, J., & Wu, J. (2024). Rest to Promote Learning: A Brain Default Mode Network Perspective. Behavioral Sciences, 14(4), 349. https://doi.org/10.3390/bs14040349