Tracing Worldwide Turkey Genetic Diversity Using D-loop Sequence Mitochondrial DNA Analysis

 ,
,  , , , ,
, , , ,  ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Ethics Statement
2.3. DNA Extraction and Amplification
2.4. Genetic Diversity and Differentiation
2.5. Genealogical Relationships between Haplotypes
3. Results
3.1. Sequence Analysis, Genetic Diversity and Differentiation
3.2. Analysis of the Internal Genetic Differentiation of the “Mundi” Group
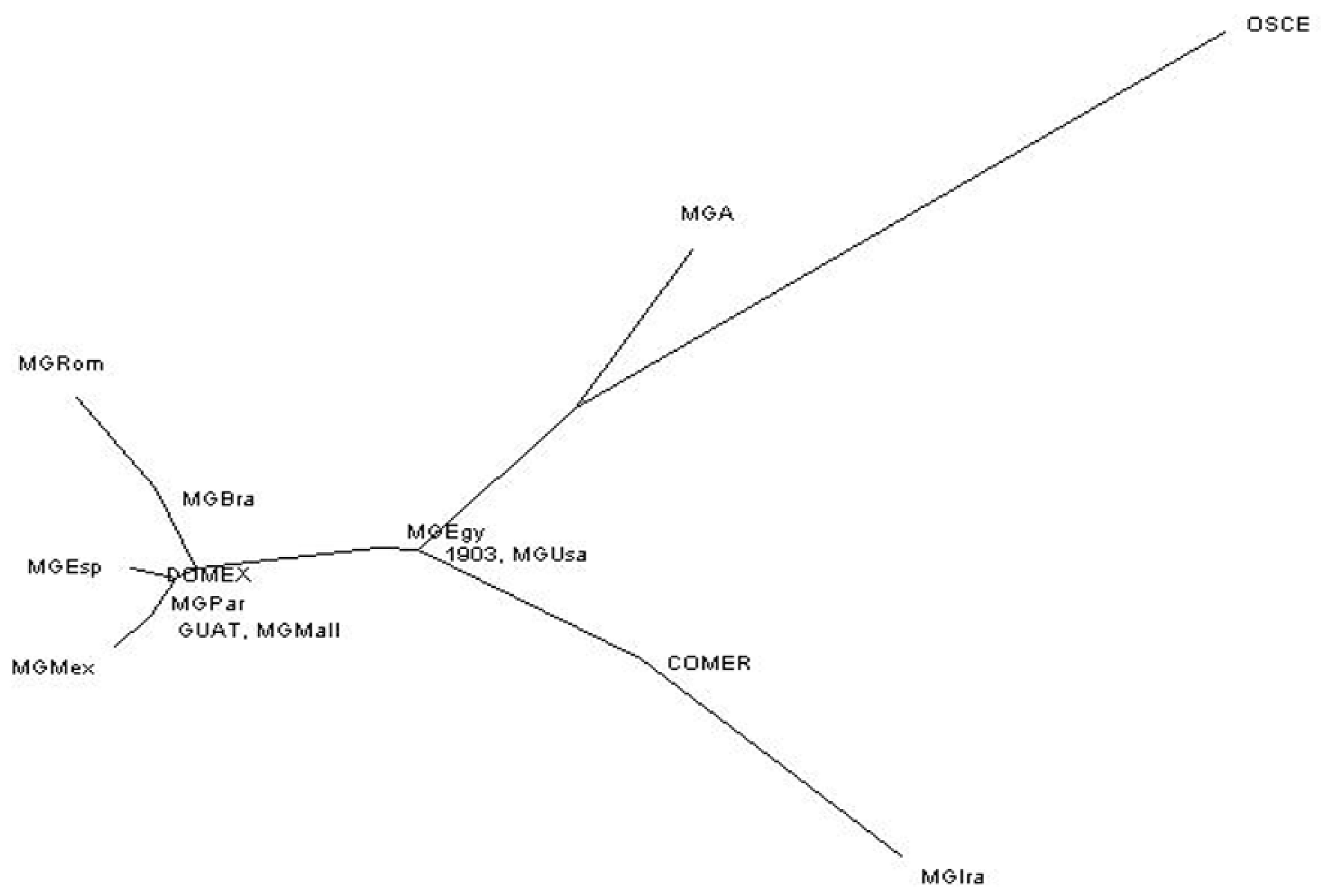
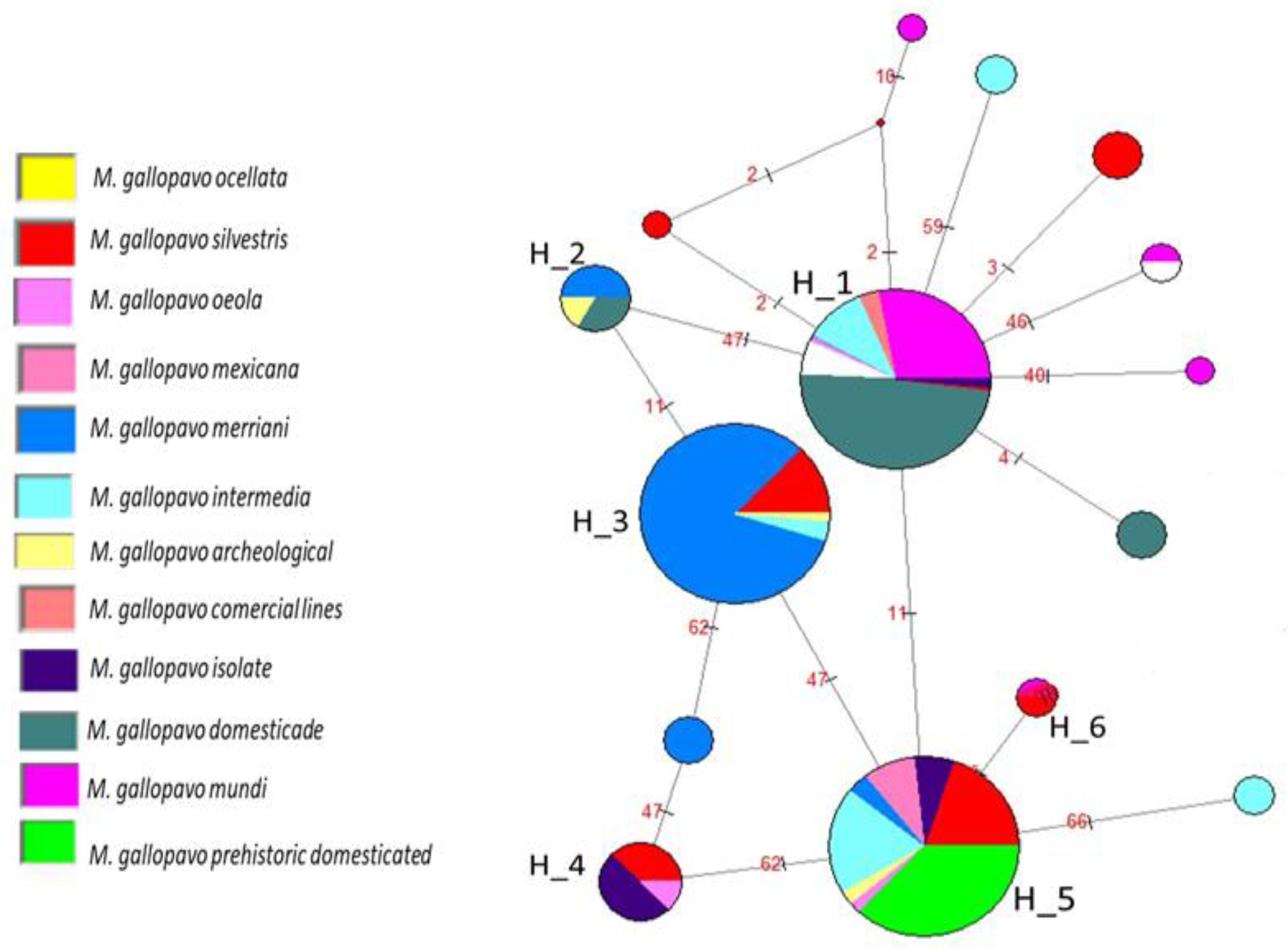
3.3. Haplotype Network
4. Discussion
4.1. Haplotype Network
4.2. Analysis of the Internal Genetic Differentiation of the “Mundi” Group
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Padilla-Jacobo, G.; Cano-Camacho, H.; López-Zavala, R.; Cornejo-Pérez, M.E.; Zavala-Páramo, M.G. Evolutionary history of Mexican domesticated and wild Meleagris gallopavo. Genet. Sel. Evol. 2018, 50, 19. [Google Scholar] [CrossRef] [PubMed]
- Mock, K.E.; Theimer, T.; Rhodes, O., Jr.; Greenberg, D.; Keim, P. Genetic variation across the historical range of the wild turkey (Meleagris gallopavo). Mol. Ecol. 2002, 11, 643–657. [Google Scholar] [CrossRef] [PubMed]
- Breitburg, E. The evolution of turkey domestication in the greater Southwest and Mesoamerica. In Culture and Contact: Charles C. Di Peso’s Gran Chichimeca; University of New Mexico Press: Albuquerque, Mexico, 1993. [Google Scholar]
- Camacho-Escobar, M.A.; Jiménez-Hidalgo, E.; Arroyo-Ledezma, J.; Sánchez-Bernal, E.I.; Pérez-Lara, E. Historia natural, domesticación y distribución del guajolote (Meleagris gallopavo) en México. Univ. y Cienc. 2011, 27, 351–360. [Google Scholar]
- Speller, C.F.; Kemp, B.M.; Wyatt, S.D.; Monroe, C.; Lipe, W.D.; Arndt, U.M.; Yang, D.Y. Ancient mitochondrial DNA analysis reveals complexity of indigenous North American turkey domestication. Proc. Natl. Acad. Sci. USA 2010, 107, 2807–2812. [Google Scholar] [CrossRef]
- Flannery, K. Vertebrate fauna and hunting practices. In Prehistory of the Tehuacan Valley, Environment and Subsistence; Byers, D.S., Ed.; University of Texas Press: Austin, TX, USA, 1967; Volume 1. [Google Scholar]
- Leopold, A.S. Wildlife of Mexico: The Game Birds and Mammals, 2nd ed.; University of California Press: London, UK, 1959; pp. 276–280. [Google Scholar]
- Schorger, A.W. The Wild Turkey: Its History and Domestication; University of Oklahoma: Norman, OK, USA, 1966; Volume 12, p. 41. [Google Scholar]
- Crawford, R. Introduction to Europe and diffusion of domesticated turkeys from the America. Arch. Zootec. 1992, 41, 2. [Google Scholar]
- Alderson, G. Conservation of breeds and maintenance of biodiversity: Justification and methodology for the conservation of Animal Genetic Resources. Arch. Zootec. 2018, 67, 300–309. [Google Scholar] [CrossRef]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Kamara, D.; Gyenai, K.; Geng, T.; Hammade, H.; Smith, E. Microsatellite marker-based genetic analysis of relatedness between commercial and heritage turkeys (Meleagris gallopavo). Poult. Sci. 2007, 86, 46–49. [Google Scholar] [CrossRef]
- Gholizadeh, M.; Mianji, G.R. Use of microsatellite markers in poultry research. Int. J. Poult. Sci. 2007, 6, 145–153. [Google Scholar]
- Vawter, L.; Brown, W.M. Nuclear and mitochondrial DNA comparisons reveal extreme rate variation in the molecular clock. Science 1986, 234, 194–196. [Google Scholar] [CrossRef]
- Adeyemi, M.; Oseni, S. Canonical discriminant analysis applied to biometric data of nigerian indigenous turkeys. Arch. Zootec. 2018, 67, 7–12. [Google Scholar] [CrossRef]
- Casquet, J.; Thebaud, C.; Gillespie, R.G. Chelex without boiling, a rapid and easy technique to obtain stable amplifiable DNA from small amounts of ethanol-stored spiders. Mol. Ecol. Resour. 2012, 12, 136–141. [Google Scholar] [CrossRef]
- Guan, X.; Silva, P.; Gyenai, K.B.; Xu, J.; Geng, T.; Tu, Z.; Samuels, D.C.; Smith, E.J. The mitochondrial genome sequence and molecular phylogeny of the turkey, Meleagris gallopavo. Anim. Genet. 2009, 40, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.-F.; Guindon, S.; Lefort, V.; Lescot, M. Phylogeny. fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Schneider, S.; Roessli, D.; Excoffier, L. Arlequin: A software for population genetics data analysis. User Man. Ver 2000, 2, 2496–2497. [Google Scholar]
- Bandelt, H.-J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies, Molecular Biology and Evolution, User Manual for SplitsTree4 V4. Mol. Biol. Evolut. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Bowen, B.; Grant, W.S. Phylogeography of the sardines (Sardinops spp.): Assessing biogeographic models and population histories in temperate upwelling zones. Evolution 1997, 51, 1601–1610. [Google Scholar] [CrossRef]
- Grant, W.; Bowen, B.W. Shallow population histories in deep evolutionary lineages of marine fishes: Insights from sardines and anchovies and lessons for conservation. J. Hered. 1998, 89, 415–426. [Google Scholar] [CrossRef]
- Guan, X.; Silva, P.; Gyenai, K.; Xu, J.; Geng, T.; Smith, E. Mitochondrial DNA-based analyses of relatedness among Turkeys, Meleagris gallopavo. Biochem. Genet. 2015, 53, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Mock, K.E.; Theimer, T.C.; Wakeling, B.F.; Rhodes, O.E., Jr.; Greenberg, D.L.; Keim, P. Verifying the origins of a reintroduced population of Gould’s wild turkey. J. Wildl. Manag. 2001, 65, 871–879. [Google Scholar] [CrossRef]
- Groeneveld, L.; Lenstra, J.; Eding, H.; Toro, M.; Scherf, B.; Pilling, D.; Negrini, R.; Finlay, E.; Jianlin, H.; Groeneveld, E. Genetic diversity in farm animals—A review. Anim. Genet. 2010, 41, 6–31. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, Y.; Yindee, M.; Li, K.Y.; Kuo, H.Y.; Ju, Y.T.; Ye, S.; Faruque, M.O.; Li, Q.; Wang, Y. Strong and stable geographic differentiation of swamp buffalo maternal and paternal lineages indicates domestication in the China/Indochina border region. Mol. Ecol. 2016, 25, 1530–1550. [Google Scholar] [CrossRef]
- Larson, G.; Fuller, D.Q. The evolution of animal domestication. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 115–136. [Google Scholar] [CrossRef]
- Thornton, E.K.; Emery, K.F.; Steadman, D.W.; Speller, C.; Matheny, R.; Yang, D. Earliest Mexican Turkeys (Meleagris gallopavo) in the Maya region: Implications for pre-hispanic animal trade and the timing of Turkey domestication. PLoS ONE 2012, 7, e42630. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Population | N | Tnm | H | Hd | π | S |
|---|---|---|---|---|---|---|
| Egypt | 10 | 19 | 5 | 0.844 | 0.0070 | 19 |
| Brazil | 11 | 1 | 2 | 0.327 | 0.0005 | 1 |
| Andalusia (Spain) | 18 | 5 | 3 | 0.451 | 0.0013 | 5 |
| Iran | 6 | 11 | 5 | 0.933 | 0.0058 | 11 |
| Majorca (Spain) | 12 | 14 | 3 | 0.318 | 0.0037 | 14 |
| Mexico | 9 | 40 | 6 | 0.833 | 0.0175 | 40 |
| Parma (Italy) | 10 | 4 | 2 | 0.200 | 0.0012 | 4 |
| Romagnolo (Italy) | 12 | 1 | 2 | 0.530 | 0.0008 | 1 |
| USA | 5 | 16 | 3 | 0.700 | 0.0102 | 16 |
| Total | 93 | 79 | 20 | 0.596 | 0.0047 | 76 |
| Population | N | H | S | Hd | π | Tajima’s D Value |
|---|---|---|---|---|---|---|
| Mundi/Domestic | 273 | 21 | 37 | 0.562 | 0.00215 | −2.38448 (NS, p < 0.01) |
| M. g. merriami | 86 | 14 | 13 | 0.724 | 0.00441 | 0.69964 (NS, p < 0.10) |
| M. g. intermedia | 77 | 13 | 13 | 0.887 | 0.00574 | −0.15106 (NS, p < 0.10) |
| M. g. osceola | 8 | 8 | 8 | 1 | 0.00538 | −1.14142 (NS, p > 0.10) |
| M. g. silvestris | 77 | 22 | 23 | 0.923 | 0.00516 | −1.57292 (NS, 0.10 > p > 0.05) |
| M. g. mexicana | 36 | 6 | 9 | 0.352 | 0.00221 | −1.65410 (NS, 0.10 > p > 0.05) |
| Total | 542 | 17 | 56 | 0.643 | 0.02442 | −2.24320 (NS, p < 0.01) |
| MGBra | MGEgy | MGAnd | MGIra | MGMall | MGMex | MGPar | MGRom | MGUsa | OSCE | MGA | 1903 | COMER | GUAT | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MGBra | 0 | |||||||||||||
| MGEgy | 0.041 | 0 | ||||||||||||
| MGAnd | 0.138 | 0.104 | 0 | |||||||||||
| MGIra | 0.755 | 0.148 | 0.623 | 0 | ||||||||||
| MGMall | 0.032 | 0.076 | 0.082 | 0.636 | 0 | |||||||||
| MGMex | 0.057 | 0.074 | 0.098 | 0.791 | −0.013 | 0 | ||||||||
| MGPar | 0.040 | 0.080 | 0.090 | 0.662 | −0.000 | −0.005 | 0 | |||||||
| MGRom | 0.038 | 0.022 | 0.309 | 0.709 | 0.202 | 0.268 | 0.213 | 0 | ||||||
| MGUsa | 0.139 | −0.000 | 0.153 | 0.288 | 0.105 | 0.105 | 0.090 | 0.203 | 0 | |||||
| OSCE | 0.907 | 0.385 | 0.853 | 0.868 | 0.821 | 0.940 | 0.846 | 0.861 | 0.570 | 0 | ||||
| MGA | 0.469 | 0.321 | 0.544 | 0.495 | 0.455 | 0.442 | 0.436 | 0.485 | 0.257 | 0.608 | 0 | |||
| 1903 | 0.302 | −0.258 | −0.093 | 0.225 | 0.097 | 0.352 | 0.120 | 0.311 | −0.275 | 0.893 | 0.218 | 0 | ||
| COMER | 0.443 | 0.149 | 0.343 | 0.020 | 0.404 | 0.427 | 0.399 | 0.466 | 0.185 | 0.511 | 0.489 | −0.163 | 0 | |
| GUAT | −0.040 | 0.043 | −0.005 | 0.825 | −0.128 | −0.116 | −0.121 | 0.200 | −0.052 | 1.000 | 0.320 | 0.384 | 0.337 | 0 |
| DOMEX | 0.103 | 0.250 | −0.023 | 0.458 | 0.095 | 0.067 | 0.092 | 0.253 | 0.219 | 0.711 | 0.702 | −0.180 | 0.376 | −0.018 |
| Source of Variation | Df | Sum of Squares | Variance Components | Percentage of Variation | Fixation Indices |
|---|---|---|---|---|---|
| Among groups | 1 | 32.51 | 0.12069 Va | 7.03 | |
| Among populations/ within groups | 3 | 14.207 | 0.17839 Vb | 10.40 | FSC: 0.11182 FST: 0.17429 FCT: 0.07033 |
| Within populations | 538 | 762.335 | 1.41698 Vc | 82.57 | |
| Total | 542 | 811.794 | 1.71607 |
| Source of Variation | Df | Sum of Squares | Variance Components | Percentage of Variation | Fixation Indices |
|---|---|---|---|---|---|
| Among groups | 2 | 38.284 | 0.05577 Va | 3.26 | |
| Among populations/within groups | 1 | 3.104 | 0.22331 Vb | 13.07 | FSC: 0.13513 FST: 0.16336 FCT: 0.03265 |
| Within Populations | 539 | 770.405 | 1.42932 Vc | 83.66 | |
| Total | 542 | 811.794 | 1.70841 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canales Vergara, A.M.; Landi, V.; Delgado Bermejo, J.V.; Martínez, A.; Cervantes Acosta, P.; Pons Barro, Á.; Bigi, D.; Sponenberg, P.; Helal, M.; Hossein Banabazi, M.; et al. Tracing Worldwide Turkey Genetic Diversity Using D-loop Sequence Mitochondrial DNA Analysis. Animals 2019, 9, 897. https://doi.org/10.3390/ani9110897
Canales Vergara AM, Landi V, Delgado Bermejo JV, Martínez A, Cervantes Acosta P, Pons Barro Á, Bigi D, Sponenberg P, Helal M, Hossein Banabazi M, et al. Tracing Worldwide Turkey Genetic Diversity Using D-loop Sequence Mitochondrial DNA Analysis. Animals. 2019; 9(11):897. https://doi.org/10.3390/ani9110897
Chicago/Turabian StyleCanales Vergara, Amado Manuel, Vincenzo Landi, Juan Vicente Delgado Bermejo, Amparo Martínez, Patricia Cervantes Acosta, Águeda Pons Barro, Daniele Bigi, Phillip Sponenberg, Mostafa Helal, Mohammad Hossein Banabazi, and et al. 2019. "Tracing Worldwide Turkey Genetic Diversity Using D-loop Sequence Mitochondrial DNA Analysis" Animals 9, no. 11: 897. https://doi.org/10.3390/ani9110897
APA StyleCanales Vergara, A. M., Landi, V., Delgado Bermejo, J. V., Martínez, A., Cervantes Acosta, P., Pons Barro, Á., Bigi, D., Sponenberg, P., Helal, M., Hossein Banabazi, M., & Camacho Vallejo, M. E. (2019). Tracing Worldwide Turkey Genetic Diversity Using D-loop Sequence Mitochondrial DNA Analysis. Animals, 9(11), 897. https://doi.org/10.3390/ani9110897