
Bridging the Gap: Using Wild African Elephant Metabolic, Reproductive, and Stress Hormone Profiles to Enhance Management in Human Care

Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Area and Population
2.2. Hormone Analysis
2.3. Statistical Analysis
3. Results
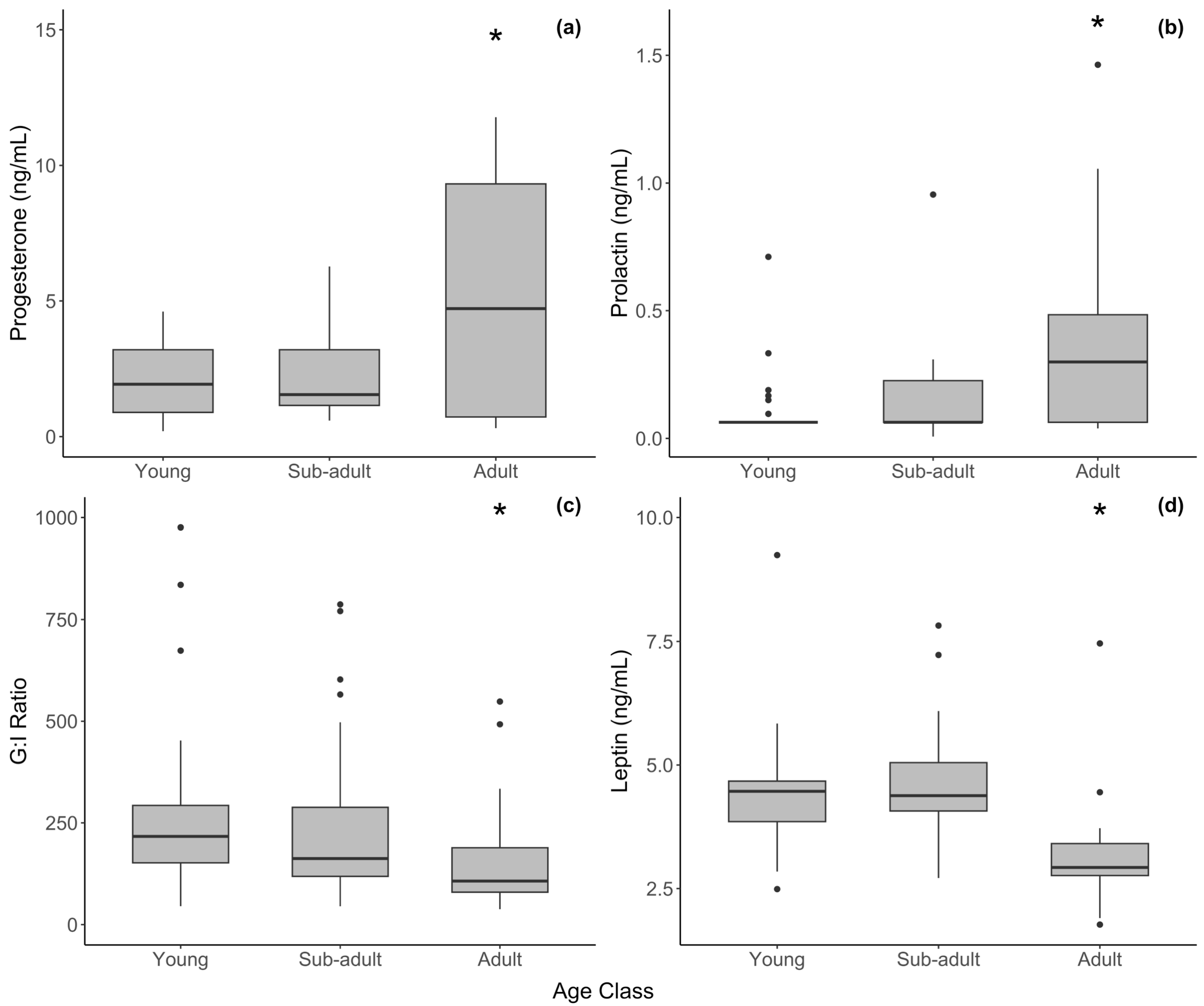
3.1. Females
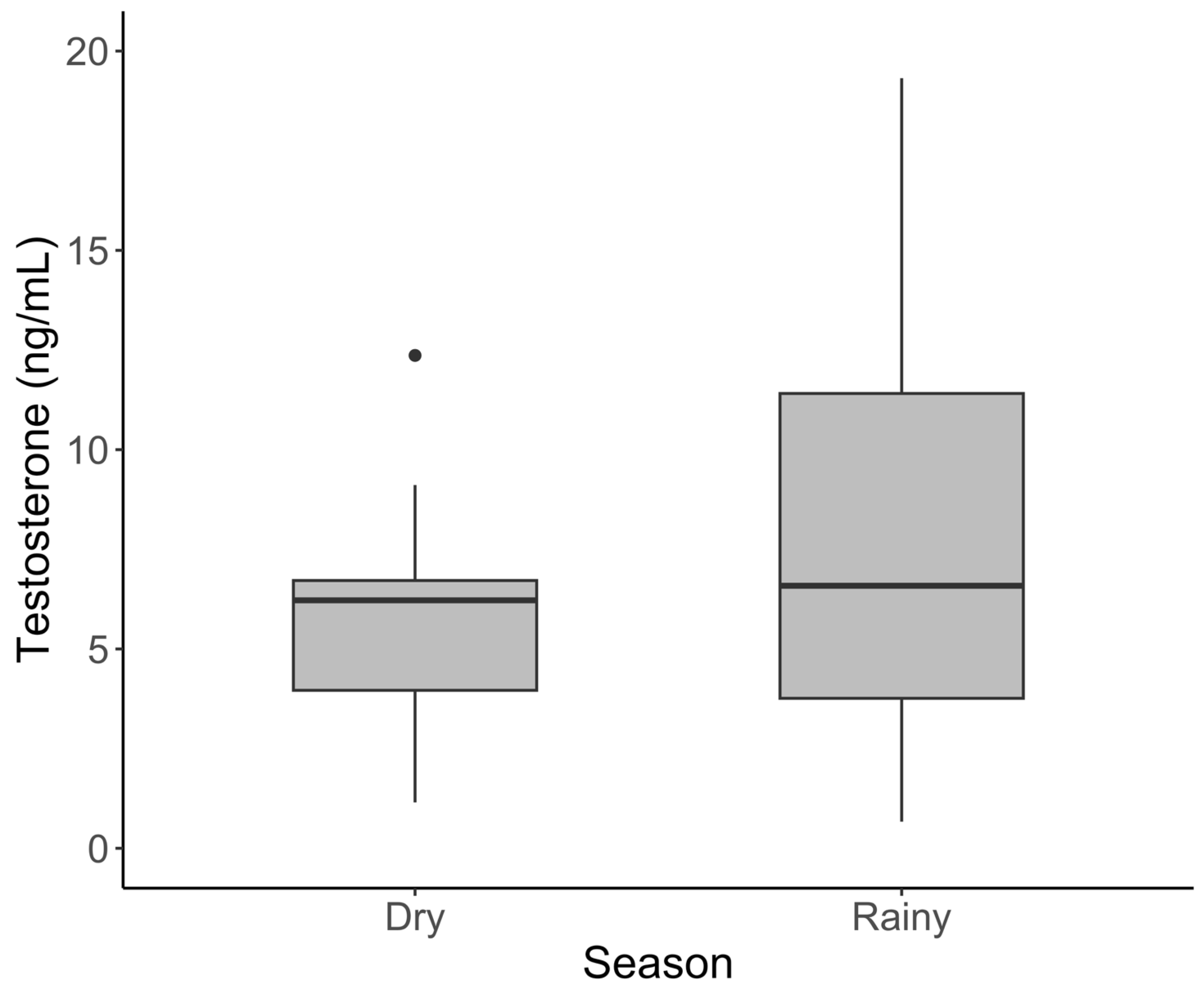
3.2. Males
3.3. Relationships Between Reproductive and Metabolic Markers
4. Discussion
4.1. Limitations
4.2. Females
4.3. Males
4.4. Comparison of Wild and Zoo Baselines
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, J.L.; Olson, D.; Keele, M.; Freeman, E.W. Survey of the reproductive cyclicity status of Asian and African elephants in North America. Zoo Biol. Publ. Affil. Am. Zoo Aquar. Assoc. 2004, 23, 309–321. [Google Scholar] [CrossRef]
- Morfeld, K.A.; Brown, J.L. Ovarian acyclicity in zoo African elephants (Loxodonta africana) is associated with high body condition scores and elevated serum insulin and leptin. Reprod. Fertil. Dev. 2014, 28, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Foley, C.; Papageorge, S.; Wasser, S. Noninvasive stress and reproductive measures of social and ecological pressures in free-ranging African elephants. Conserv. Biol. 2001, 15, 1134–1142. [Google Scholar] [CrossRef]
- Viljoen, J.J.; Ganswindt, A.; Palme, R.; Reynecke, H.C.; Du Toit, J.T.; Langbauer, W.R., Jr. Measurement of concentrations of faecal glucocorticoid metabolites in free-ranging African elephants within the Kruger National Park. Koedoe Afr. Prot. Area Conserv. Sci. 2008, 50, 18–21. [Google Scholar] [CrossRef]
- Ghosal, R.; Ganswindt, A.; Seshagiri, P.B.; Sukumar, R. Endocrine correlates of musth in free-ranging Asian elephants (Elephas maximus) determined by non-invasive faecal steroid hormone metabolite measurements. PLoS ONE 2013, 8, e84787. [Google Scholar] [CrossRef]
- Pokharel, S.S.; Seshagiri, P.B.; Sukumar, R. Assessment of season-dependent body condition scores in relation to faecal glucocorticoid metabolites in free-ranging Asian elephants. Conserv. Physiol. 2017, 5, cox039. [Google Scholar] [CrossRef]
- Allan, C.A.; McLachlan, R.I. Androgens and obesity. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 224–232. [Google Scholar] [CrossRef]
- Gonzalez-Bulnes, A.; Pallares, P.; Ovilo, C. Ovulation, implantation and placentation in females with obesity and metabolic disorders: Life in the balance. Endocr. Metab. Immune Disord. Drug Targets 2011, 11, 285–301. [Google Scholar] [CrossRef]
- Kil, D.Y.; Swanson, K.S. Endocrinology of obesity. Vet. Clin. N. Am. Small Anim. Pract. 2010, 40, 205–219. [Google Scholar] [CrossRef]
- Tsao, C.-W.; Liu, C.-Y.; Chou, Y.-C.; Cha, T.-L.; Chen, S.-C.; Hsu, C.-Y. Exploration of the association between obesity and semen quality in a 7630 male population. PLoS ONE 2015, 10, e0119458. [Google Scholar] [CrossRef]
- Zheng, C.; Liu, Y.; Xu, C.; Zeng, S.; Wang, Q.; Guo, Y.; Li, J.; Li, S.; Dong, M.; Luo, X.; et al. Association between obesity and the prevalence of dyslipidemia in middle-aged and older people: An observational study. Sci. Rep. 2024, 14, 11974. [Google Scholar] [CrossRef] [PubMed]
- Ange, K.; Crissey, S.D.; Doyle, C.; Lance, K.; Hintz, H. A survey of African (Loxodonta africana) and Asian (Elephas maximus) elephant diets and measured body dimensions compared to their estimated nutrient requirements. In Proceedings of the AZA Nutrition Advisory Group, 4th Conference on Zoo and Wildlife Nutrition, Lake Buena Vista, FL, USA, 19–23 September 2001; Edwards, M.S., Lisi, K.J., Schlegel, M.L., Bray, R.E., Eds.; 2001; pp. 5–14. [Google Scholar]
- Hatt, J.M.; Clauss, M. Feeding Asian and African elephants Elephas maximus and Loxodonta africana in captivity. Int. Zoo Yearb. 2006, 40, 88–95. [Google Scholar] [CrossRef]
- Clubb, R.; Rowcliffe, M.; Lee, P.; Mar, K.U.; Moss, C.; Mason, G.J. Fecundity and population viability in female zoo elephants: Problems and possible solutions. Anim. Welf. 2009, 18, 237–247. [Google Scholar] [CrossRef]
- Lewis, K.D.; Shepherdson, D.J.; Owens, T.M.; Keele, M. A survey of elephant husbandry and foot health in North American zoos. Zoo Biol. 2010, 29, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Morfeld, K.A.; Meehan, C.L.; Hogan, J.N.; Brown, J.L. Assessment of Body Condition in African (Loxodonta africana) and Asian (Elephas maximus) Elephants in North American Zoos and Management Practices Associated with High Body Condition Scores. PLoS ONE 2016, 11, e0155146. [Google Scholar] [CrossRef]
- Campbell, K.M.; Wilson, J.A.; Morfeld, K.A. Predictors of testosterone in zoo-managed male African elephants (Loxodonta Africana). Zoo Biol. 2022, 42, 268–282. [Google Scholar] [CrossRef]
- Hartz, A.; Barboriak, P.N.; Wong, A.; Katayama, K.P.; Rimm, A.A. The association of obesity with infertility and related menstural abnormalities in women. Int. J. Obes. 1979, 3, 57–73. [Google Scholar]
- Bray, G.A. Obesity and reproduction. Hum. Reprod. 1997, 12, 26–32. [Google Scholar] [CrossRef]
- Norman, R.; Clark, A. Obesity and reproductive disorders: A review. Reprod. Fertil. Dev. 1998, 10, 55–63. [Google Scholar] [CrossRef]
- Irvine, L.; Shaw, R. The effects of patient obesity in gynaecological practice. Curr. Obstet. Gynaecol. 2003, 13, 179–184. [Google Scholar] [CrossRef]
- Pasquali, R.; Pelusi, C.; Genghini, S.; Cacciari, M.; Gambineri, A. Obesity and reproductive disorders in women. Hum. Reprod. Update 2003, 9, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Plum, L.; Belgardt, B.F.; Brüning, J.C. Central insulin action in energy and glucose homeostasis. J. Clin. Investig. 2006, 116, 1761–1766. [Google Scholar] [CrossRef] [PubMed]
- Sliwowska, J.H.; Fergani, C.; Gawałek, M.; Skowronska, B.; Fichna, P.; Lehman, M.N. Insulin: Its role in the central control of reproduction. Physiol. Behav. 2014, 133, 197–206. [Google Scholar] [CrossRef]
- Cardozo, E.; Pavone, M.E.; Hirshfeld-Cytron, J.E. Metabolic syndrome and oocyte quality. Trends Endocrinol. Metab. 2011, 22, 103–109. [Google Scholar] [CrossRef]
- Oghbaei, H.; Fattahi, A.; Hamidian, G.; Sadigh-Eteghad, S.; Ziaee, M.; Mahmoudi, J. A closer look at the role of insulin for the regulation of male reproductive function. Gen. General. Comp. Endocrinol. 2021, 300, 113643. [Google Scholar] [CrossRef] [PubMed]
- Mantzoros, C.S. Role of leptin in reproduction. Ann. N. Y. Acad. Sci. 2000, 900, 174–183. [Google Scholar] [CrossRef]
- Moschos, S.; Chan, J.L.; Mantzoros, C.S. Leptin and reproduction: A review. Fertil. Steril. 2002, 77, 433–444. [Google Scholar] [CrossRef]
- Qaid, M.M.; Abdelrahman, M.M. Role of insulin and other related hormones in energy metabolism—A review. Cogent Food Agric. 2016, 2, 1267691. [Google Scholar] [CrossRef]
- Morais, J.B.S.; Severo, J.S.; Beserra, J.B.; de Oiveira, A.R.S.; Cruz, K.J.C.; de Sousa Melo, S.R.; do Nascimento, G.V.R.; de Macedo, G.F.S.; do Nascimento Marreiro, D. Association between cortisol, insulin resistance and zinc in obesity: A mini-review. Biol. Trace Elem. Res. 2019, 191, 323–330. [Google Scholar] [CrossRef]
- Fu, J.-H.; Xie, S.-R.; Kong, S.-J.; Wang, Y.; Wei, W.; Shan, Y.; Luo, Y.-M. The combination of a high-fat diet and chronic stress aggravates insulin resistance in Wistar male rats. Exp. Clin. Endocrinol. Diabetes 2009, 117, 354–360. [Google Scholar] [CrossRef]
- Hewagalamulage, S.D.; Lee, T.; Clarke, I.; Henry, B. Stress, cortisol, and obesity: A role for cortisol responsiveness in identifying individuals prone to obesity. Domest. Anim. Endocrinol. 2016, 56, S112–S120. [Google Scholar] [CrossRef] [PubMed]
- Auwerx, J.; Staels, B. Leptin. Lancet 1998, 351, 737–742. [Google Scholar] [CrossRef] [PubMed]
- van Aarde, R.; Ferreira, S.; Jackson, T.; Page, B.; De Beer, Y.; Gough, K.; Guldemond, R.; Junker, J.; Olivier, P.; Ott, T. Elephant population biology and ecology. In Elephant Management: A Scientific Assessment for South Africa; Scholes, R.J., Mennel, K., Eds.; Wits University Press: Johannesburg, South Africa, 2008. [Google Scholar]
- Ferreira, S.; Crowhurst, E.; Greaver, C.; Simms, C. Sample-Based Estimates of Elephants in Kruger National Park, South Africa. Afr. J. Wildl. Res. 2024, 54, 122. [Google Scholar] [CrossRef]
- Holl, K.; Lundin, E.; Kaasila, M.; Grankvist, K.; Afanasyeva, Y.; Hallmans, G.; Lehtinen, M.; Pukkala, E.; Surcel, H.-M.; Toniolo, P. Effect of long-term storage on hormone measurements in samples from pregnant women: The experience of the Finnish Maternity Cohort. Acta Oncol. 2008, 47, 406–412. [Google Scholar] [CrossRef]
- Kleeberger, C.; Shore, D.; Gunter, E.; Sandler, D.P.; Weinberg, C.R. The effects of long-term storage on commonly measured serum analyte levels. Epidemiology 2018, 29, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, C.; Zhang, X.; Bian, Y.; Cao, Y. Assessing the impact of long-term storage on the quality and integrity of biological specimens in a reproductive biobank. Bioeng. Transl. Med. 2024, 9, e10692. [Google Scholar] [CrossRef]
- Gislefoss, R.E.; Grimsrud, T.K.; Mørkrid, L. Stability of selected serum hormones and lipids after long-term storage in the Janus Serum Bank. Clin. Biochem. 2015, 48, 364–369. [Google Scholar] [CrossRef]
- Wagner-Golbs, A.; Neuber, S.; Kamlage, B.; Christiansen, N.; Bethan, B.; Rennefahrt, U.; Schatz, P.; Lind, L. Effects of long-term storage at− 80 C on the human plasma metabolome. Metabolites 2019, 9, 99. [Google Scholar] [CrossRef]
- Giampietro, O.; Navalesi, R.; Buzzigoli, G.; Boni, C.; Benzi, L. Decrease in plasma glucose concentration during storage at-20 degrees C. Clin. Chem. 1980, 26, 1710–1712. [Google Scholar] [CrossRef]
- Bartoń, K. Package ‘MuMIn’. CRAN Repost. 2016. Available online: https://cran.r-project.org/web/packages/MuMIn/MuMIn.pdf (accessed on 4 April 2025).
- Wittemyer, G.; Ganswindt, A.; Hodges, K. The impact of ecological variability on the reproductive endocrinology of wild female African elephants. Horm. Behav. 2007, 51, 346–354. [Google Scholar] [CrossRef]
- Thitaram, C.; Brown, J.L.; Pongsopawijit, P.; Chansitthiwet, S.; Wongkalasin, W.; Daram, P.; Roongsri, R.; Kalmapijit, A.; Mahasawangkul, S.; Rojansthien, S. Seasonal effects on the endocrine pattern of semi-captive female Asian elephants (Elephas maximus): Timing of the anovulatory luteinizing hormone surge determines the length of the estrous cycle. Theriogenology 2008, 69, 237–244. [Google Scholar] [CrossRef]
- Hall-Martin, A. Role of musth in the reproductive strategy of the African elephant (Loxodonta africana). S. Afr. J. Sci. 1987, 83, 616–620. [Google Scholar]
- Glaeser, S.S.; Edwards, K.L.; Wielebnowski, N.; Brown, J.L. Effects of physiological changes and social life events on adrenal glucocorticoid activity in female zoo-housed Asian elephants (Elephas maximus). PLoS ONE 2020, 15, e0241910. [Google Scholar] [CrossRef]
- Ganswindt, A.; Muenscher, S.; Henley, M.; Henley, S.; Heistermann, M.; Palme, R.; Thompson, P.; Bertschinger, H. Endocrine correlates of musth and the impact of ecological and social factors in free-ranging African elephants (Loxodonta africana). Horm. Behav. 2010, 57, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Freeman, E.W.; Guagnano, G.; Olson, D.; Keele, M.; Brown, J.L. Social factors influence ovarian acyclicity in captive African elephants (Loxodonta africana). Zoo Biol. Publ. Affil. Am. Zoo Aquar. Assoc. 2009, 28, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Prado, N.A.; Krcmarik, M.; Paris, S.; Larry, L.R.; Brown, J.L. Abnormal prolactin secretion is associated with changes in metabolic health biomarkers in acyclic female African elephants (Loxodonta africana). Theriogenol. Wild 2023, 3, 100062. [Google Scholar] [CrossRef]
- Woolley, L.-A.; Millspaugh, J.J.; Woods, R.J.; Van Rensburg, S.J.; Page, B.R.; Slotow, R. Intraspecific strategic responses of African elephants to temporal variation in forage quality. J. Wildl. Manag. 2009, 73, 827–835. [Google Scholar] [CrossRef]
- Pokharel, S.; Singh, B.; Seshagiri, P.; Sukumar, R. Lower levels of glucocorticoids in crop-raiders: Diet quality as a potential ‘pacifier’against stress in free-ranging Asian elephants in a human-production habitat. Anim. Conserv. 2019, 22, 177–188. [Google Scholar] [CrossRef]
- Frithioff-Bøjsøe, C.; Lund, M.A.; Kloppenborg, J.T.; Nielsen, T.T.; Fonvig, C.E.; Lausten-Thomsen, U.; Hedley, P.L.; Hansen, T.; Pedersen, O.B.; Christiansen, M. Glucose metabolism in children and adolescents: Population-based reference values and comparisons to children and adolescents enrolled in obesity treatment. Pediatr. Diabetes 2019, 20, 538–548. [Google Scholar] [CrossRef]
- Lecke, S.B.; Morsch, D.M.; Spritzer, P.M. Leptin and adiponectin in the female life course. Braz. J. Med. Biol. Res. 2011, 44, 381–387. [Google Scholar] [CrossRef]
- Lee, P.C.; Fishlock, V.; Webber, C.E.; Moss, C.J. The reproductive advantages of a long life: Longevity and senescence in wild female African elephants. Behav. Ecol. Sociobiol. 2016, 70, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.C.; Bussière, L.F.; Webber, C.E.; Poole, J.H.; Moss, C.J. Enduring consequences of early experiences: 40 year effects on survival and success among African elephants (Loxodonta africana). Biol. Lett. 2013, 9, 20130011. [Google Scholar] [CrossRef] [PubMed]
- Havel, P.J.; Kasim-karakas, S.; Dubuc, G.R.; Muller, W.; Phinney, S.D. Gender differences in plasma leptin concentrations. Nat. Med. 1996, 2, 949–950. [Google Scholar] [CrossRef] [PubMed]
- Morfeld, K.A.; Brown, J.L. Metabolic health assessment of zoo elephants: Management factors predicting leptin levels and the glucose-to-insulin ratio and their associations with health parameters. PLoS ONE 2017, 12, e0188701. [Google Scholar] [CrossRef]
- Chusyd, D.E.; Brown, J.L.; Hambly, C.; Johnson, M.S.; Morfeld, K.; Patki, A.; Speakman, J.R.; Allison, D.B.; Nagy, T.R. Adiposity and reproductive cycling status in zoo African elephants. Obesity 2018, 26, 103–110. [Google Scholar] [CrossRef]
- Holdgate, M.R.; Meehan, C.L.; Hogan, J.N.; Miller, L.J.; Soltis, J.; Andrews, J.; Shepherdson, D.J. Walking behavior of zoo elephants: Associations between GPS-measured daily walking distances and environmental factors, social factors, and welfare indicators. PLoS ONE 2016, 11, e0150331. [Google Scholar] [CrossRef]
- Rothwell, E.S.; Bercovitch, F.B.; Andrews, J.R.; Anderson, M.J. Estimating Daily Walking Distance of Captive African Elephants Using an Accelerometer; Wiley Online Library: Hoboken, NJ, USA, 2011; ISBN 0733-3188. [Google Scholar]
- Rowell, Z.E. Locomotion in captive Asian elephants (Elephas maximus). J. Zoo Aquar. Res. 2014, 2, 130–135. [Google Scholar]
- Hufenus, R.; Schiffmann, C.; Hatt, J.M.; Müller, D.W.; Lackey, L.B.; Clauss, M.; Zerbe, P. Seasonality of reproduction in Asian elephants Elephas maximus and African elephants Loxodonta africana: Underlying photoperiodic cueing? Mammal. Rev. 2018, 48, 261–276. [Google Scholar] [CrossRef]
- Rasmussen, H.B.; Ganswindt, A.; Douglas-Hamilton, I.; Vollrath, F. Endocrine and behavioral changes in male African elephants: Linking hormone changes to sexual state and reproductive tactics. Horm. Behav. 2008, 54, 539–548. [Google Scholar] [CrossRef]
- Ganswindt, A.; Heistermann, M.; Hodges, K. Physical, physiological, and behavioral correlates of musth in captive African elephants (Loxodonta africana). Physiol. Biochem. Zool. 2005, 78, 505–514. [Google Scholar] [CrossRef]
- Rangel-Negrín, A.; Alfaro, J.; Valdez, R.; Romano, M.; Serio-Silva, J. Stress in Yucatan spider monkeys: Effects of environmental conditions on fecal cortisol levels in wild and captive populations. Anim. Conserv. 2009, 12, 496–502. [Google Scholar] [CrossRef]
- Davies, N.; Gramotnev, G.; Seabrook, L.; McAlpine, C.; Baxter, G.; Lunney, D.; Bradley, A. Climate-driven changes in diet composition and physiological stress in an arboreal folivore at the semi-arid edge of its distribution. Biol. Conserv. 2014, 172, 80–88. [Google Scholar] [CrossRef]
- Brown, J.L.; Somerville, M.; Riddle, H.S.; Keele, M.; Duer, C.K.; Freeman, E.W. Comparative endocrinology of testicular, adrenal and thyroid function in captive Asian and African elephant bulls. Gen. General. Comp. Endocrinol. 2007, 151, 153–162. [Google Scholar] [CrossRef]
- Chave, E.; Edwards, K.L.; Paris, S.; Prado, N.; Morfeld, K.A.; Brown, J.L. Variation in metabolic factors and gonadal, pituitary, thyroid, and adrenal hormones in association with musth in African and Asian elephant bulls. Gen. General. Comp. Endocrinol. 2019, 276, 1–13. [Google Scholar] [CrossRef]
- McAuley, K.A.; Williams, S.M.; Mann, J.I.; Walker, R.J.; Lewis-Barned, N.J.; Temple, L.A.; Duncan, A.W. Diagnosing insulin resistance in the general population. Diabetes Care 2001, 24, 460–464. [Google Scholar] [CrossRef] [PubMed]
- Pasquali, R.; Casimirri, F.; De Iasio, R.; Mesini, P.; Boschi, S.; Chierici, R.; Flamia, R.; Biscotti, M.; Vicennati, V. Insulin regulates testosterone and sex hormone-binding globulin concentrations in adult normal weight and obese men. J. Clin. Endocrinol. Metab. 1995, 80, 654–658. [Google Scholar]
- Kahn, B.B.; Flier, J.S. Obesity and insulin resistance. J. Clin. Investig. 2000, 106, 473–481. [Google Scholar] [CrossRef]
- Moriarty-Kelsey, M.; Harwood, J.E.; Travers, S.H.; Zeitler, P.S.; Nadeau, K.J. Testosterone, obesity and insulin resistance in young males: Evidence for an association between gonadal dysfunction and insulin resistance during puberty. J. Pediatr. Endocrinol. Metab. 2010, 23, 1281–1287. [Google Scholar] [CrossRef]
- Poole, J.H. Rutting behavior in African elephants: The phenomenon of musth. Behaviour 1987, 102, 283–316. [Google Scholar] [CrossRef]
- Rasmussen, L.; Perrin, T.E. Physiological correlates of musth: Lipid metabolites and chemical composition of exudates. Physiol. Behav. 1999, 67, 539–549. [Google Scholar] [CrossRef]
- Cahill, G.; Herrera, M.; Morgan, A.m.; Soeldner, J.; Steinke, J.; Levy, P.; Reichard, G.; Kipnis, D. Hormone-fuel interrelationships during fasting. J. Clin. Investig. 1966, 45, 1751–1769. [Google Scholar] [CrossRef] [PubMed]
- Olefsky, J.M. Effects of fasting on insulin binding, glucose transport, and glucose oxidation in isolated rat adipocytes: Relationships between insulin receptors and insulin action. J. Clin. Investig. 1976, 58, 1450–1460. [Google Scholar] [CrossRef]
- Liang, Q.; Zhong, L.; Zhang, J.; Wang, Y.; Bornstein, S.R.; Triggle, C.R.; Ding, H.; Lam, K.S.; Xu, A. FGF21 maintains glucose homeostasis by mediating the cross talk between liver and brain during prolonged fasting. Diabetes 2014, 63, 4064–4075. [Google Scholar] [CrossRef]
- Silva, I.D.; Kuruwita, V.Y. Hematology, plasma, and serum biochemistry values in free-ranging elephants (Elephas maximus ceylonicus) in Sri Lanka. J. Zoo Wildl. Med. 1993, 24, 434–439. [Google Scholar]
- Scott, N.; Riddle, H. Assessment of musth in captivity: A survey of factors affecting the frequency and duration of musth in captive male elephants Elephas maximus—Loxodonta africana. J. Elephant Manag. Assoc. 2003, 14, 11–15. [Google Scholar]
- Ducluzeau, P.-H.; Cousin, P.; Malvoisin, E.; Bornet, H.; Vidal, H.; Laville, M.; Pugeat, M. Glucose-to-insulin ratio rather than sex hormone-binding globulin and adiponectin levels is the best predictor of insulin resistance in nonobese women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2003, 88, 3626–3631. [Google Scholar] [CrossRef]
- Kiso, W.K.; Brown, J.L.; Siewerdt, F.; Schmitt, D.L.; Olson, D.; Crichton, E.G.; Pukazhenthi, B.S. Liquid semen storage in elephants (Elephas maximus and Loxodonta africana): Species differences and storage optimization. J. Androl. 2011, 32, 420–431. [Google Scholar] [CrossRef]
- Brown, J.L. Comparative reproductive biology of elephants. Adv. Exp. Med. Biol. 2014, 753, 135–169. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hormone | Age Class | N | Min. | Max. | Mean ± SE |
|---|---|---|---|---|---|
| Progesterone | Young | 25 | 0.200 | 4.608 | 2.051 ± 0.251 |
| (ng/mL) | Sub-adult | 28 | 0.589 | 6.273 | 2.343 ± 0.302 |
| Adult | 29 | 0.312 | 11.778 | 5.130 ± 0.752 | |
| Prolactin | Young | 28 | 0.063 | 0.711 | 0.108 ± 0.025 |
| (ng/mL) | Sub-adult | 29 | 0.007 | 0.955 | 0.148 ± 0.033 |
| Adult | 29 | 0.039 | 1.463 | 0.383 ± 0.066 | |
| Cortisol | Young | 26 | 74.449 | 1293.850 | 703.151 ± 66.713 |
| (ng/mL) | Sub-adult | 28 | 181.655 | 1203.070 | 696.177 ± 49.513 |
| Adult | 30 | 160.705 | 1075.590 | 503.823 ± 47.076 | |
| Leptin | Young | 25 | 2.489 | 5.838 | 4.264 ± 0.155 |
| (ng/mL) | Sub-adult | 25 | 2.713 | 5.542 | 4.315 ± 0.135 |
| Adult | 28 | 1.771 | 4.447 | 2.971 ± 0.113 | |
| Insulin | Young | 28 | 0.038 | 1.534 | 0.430 ± 0.064 |
| (mg/mL) | Sub-adult | 29 | 0.050 | 0.987 | 0.388 ± 0.048 |
| Adult | 30 | 0.050 | 2.780 | 0.904 ± 0.115 | |
| Glucose | Young | 27 | 20.0 | 161.0 | 87.3 ± 7.0 |
| (mg/dL) | Sub-adult | 28 | 20.0 | 150.5 | 70.5 ± 5.5 |
| Adult | 30 | 36.0 | 153.5 | 98.4 ± 4.4 | |
| G/I Ratio | Young | 27 | 45.306 | 2342.105 | 355.813 ± 87.593 |
| Sub-adult | 28 | 45.058 | 1160.000 | 317.577 ± 59.676 | |
| Adult | 30 | 37.950 | 1450.000 | 202.273 ± 48.506 |
| Hormone | Age Class | N | Min. | Max. | Mean ± SE |
|---|---|---|---|---|---|
| Testosterone | Young | 30 | 0.374 | 2.195 | 0.736 ± 0.083 |
| (ng/mL) | Sub-adult | 30 | 0.144 | 10.084 | 2.253 ± 0.476 |
| Adult | 27 | 0.668 | 41.166 | 9.047 ± 1.708 | |
| Cortisol | Young | 30 | 237.352 | 1496.071 | 750.182 ± 60.024 |
| (ng/mL) | Sub-adult | 29 | 167.340 | 1022.371 | 529.807 ± 44.888 |
| Adult | 28 | 134.413 | 1703.508 | 714.923 ± 74.791 | |
| Leptin | Young | 25 | NA | 4.064 | NA |
| (ng/mL) | Sub-adult | NA | NA | NA | NA |
| Adult | 24 | 1.667 | 6.103 | 3.284 ± 0.231 | |
| Insulin | Young | 30 | 0.050 | 1.238 | 0.372 ± 0.050 |
| (mg/mL) | Sub-adult | 30 | 0.044 | 1.715 | 0.388 ± 0.067 |
| Adult | 29 | 0.031 | 3.000 | 0.697 ± 0.137 | |
| Glucose | Young | 30 | 39.5 | 142.0 | 88.2 ± 5.2 |
| (mg/dL) | Sub-adult | 30 | 20.0 | 125.5 | 107.6 ± 3.4 |
| Adult | 28 | 65.0 | 135.0 | 98.4 ± 4.4 | |
| G/I Ratio | Young | 27 | 96.527 | 848.739 | 290.692 ± 33.664 |
| Sub-adult | 28 | 34.985 | 924.000 | 417.223 ± 51.448 | |
| Adult | 23 | 38.667 | 591.324 | 221.935 ± 32.316 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campbell, K.M.; Marolf, C.J.; Buss, P.E.; Morfeld, K.A. Bridging the Gap: Using Wild African Elephant Metabolic, Reproductive, and Stress Hormone Profiles to Enhance Management in Human Care. Animals 2025, 15, 1863. https://doi.org/10.3390/ani15131863
Campbell KM, Marolf CJ, Buss PE, Morfeld KA. Bridging the Gap: Using Wild African Elephant Metabolic, Reproductive, and Stress Hormone Profiles to Enhance Management in Human Care. Animals. 2025; 15(13):1863. https://doi.org/10.3390/ani15131863
Chicago/Turabian StyleCampbell, Kaitlyn M., Chelsi J. Marolf, Peter E. Buss, and Kari A. Morfeld. 2025. "Bridging the Gap: Using Wild African Elephant Metabolic, Reproductive, and Stress Hormone Profiles to Enhance Management in Human Care" Animals 15, no. 13: 1863. https://doi.org/10.3390/ani15131863
APA StyleCampbell, K. M., Marolf, C. J., Buss, P. E., & Morfeld, K. A. (2025). Bridging the Gap: Using Wild African Elephant Metabolic, Reproductive, and Stress Hormone Profiles to Enhance Management in Human Care. Animals, 15(13), 1863. https://doi.org/10.3390/ani15131863