

First Description of the Underwater Sounds in the Mediterranean Monk Seal Monachus monachus in Greece: Towards Establishing a Vocal Repertoire
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Acoustic Recording and Analysis
2.3. Statistical Analysis
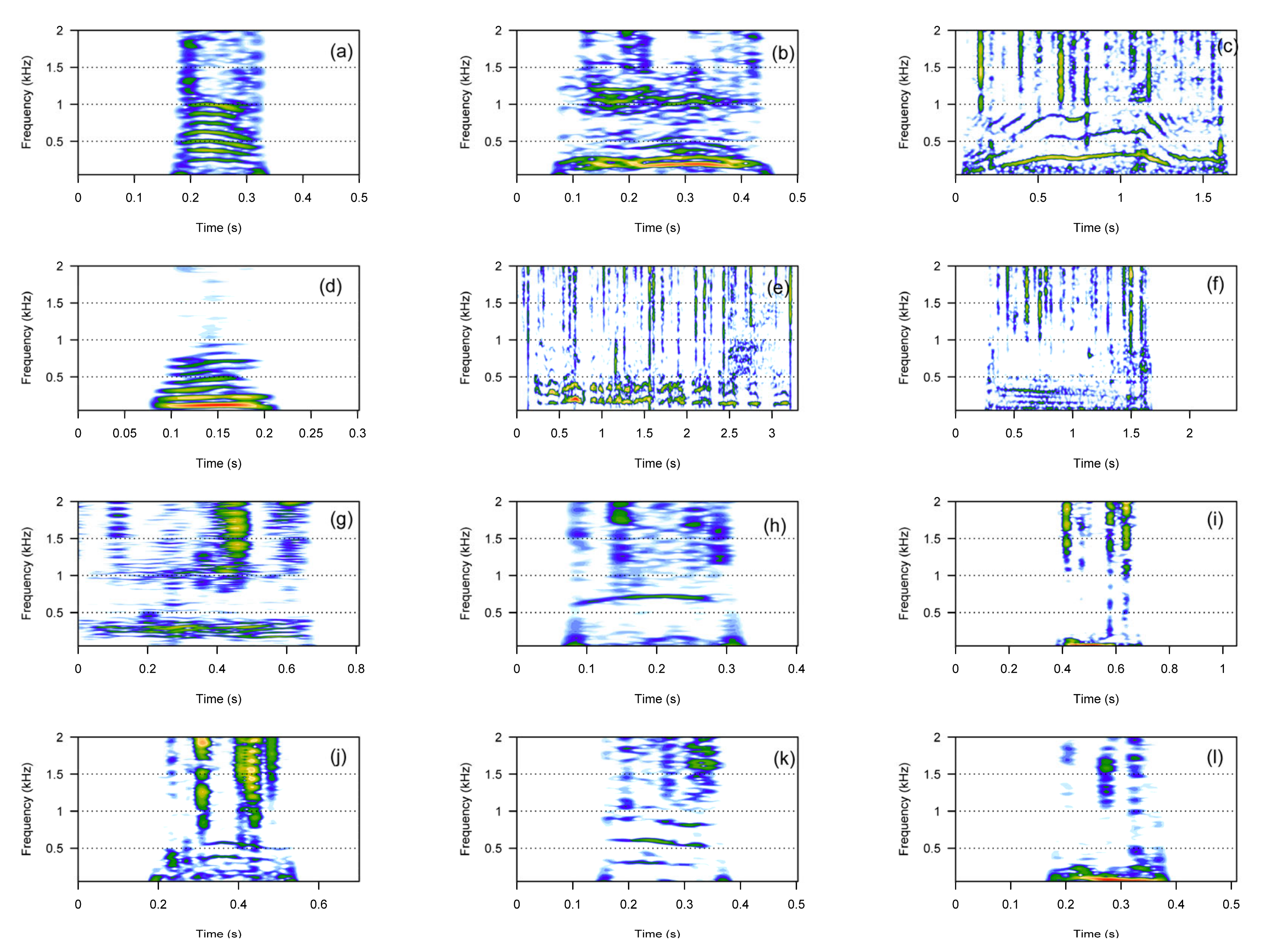
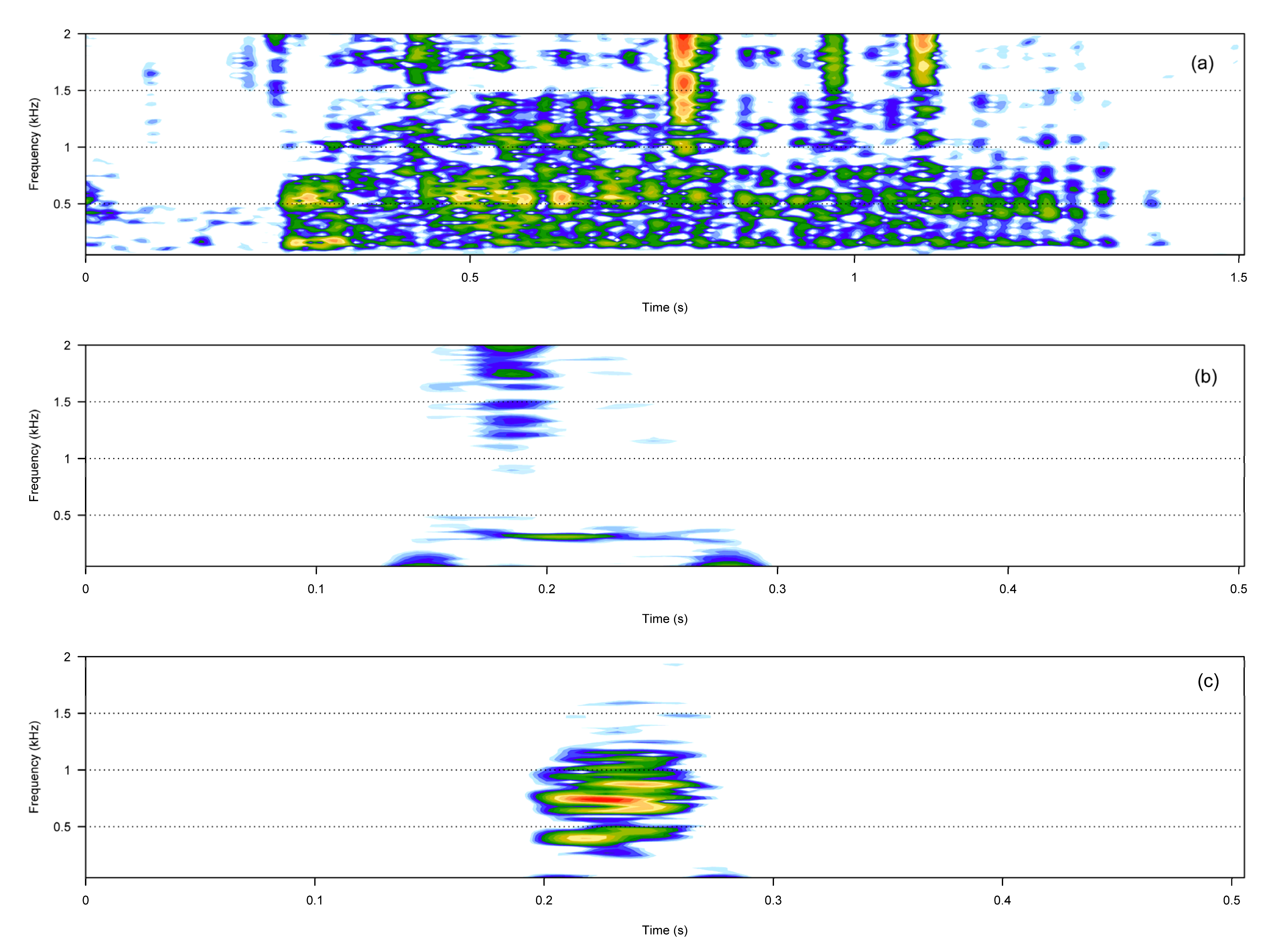
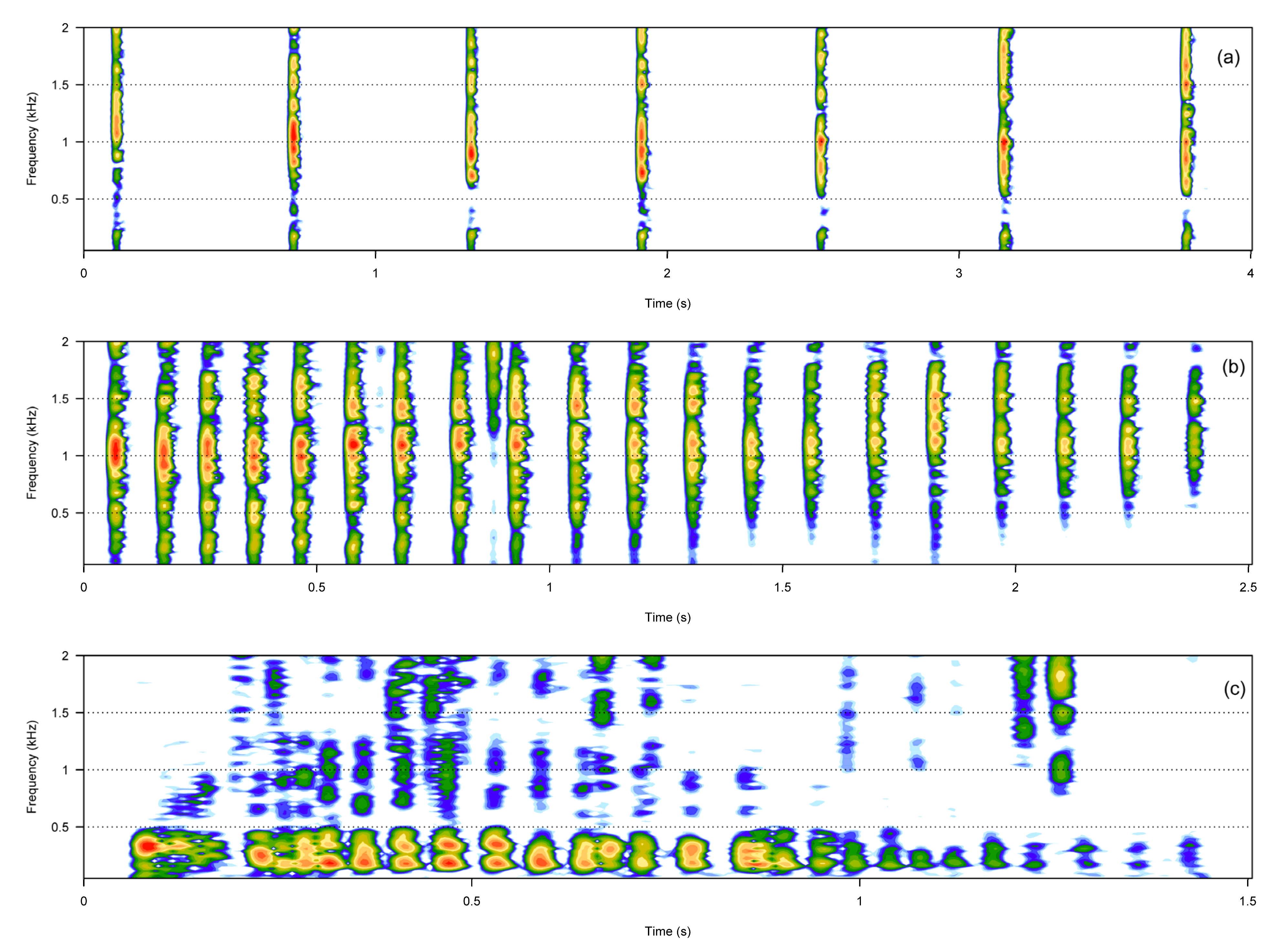
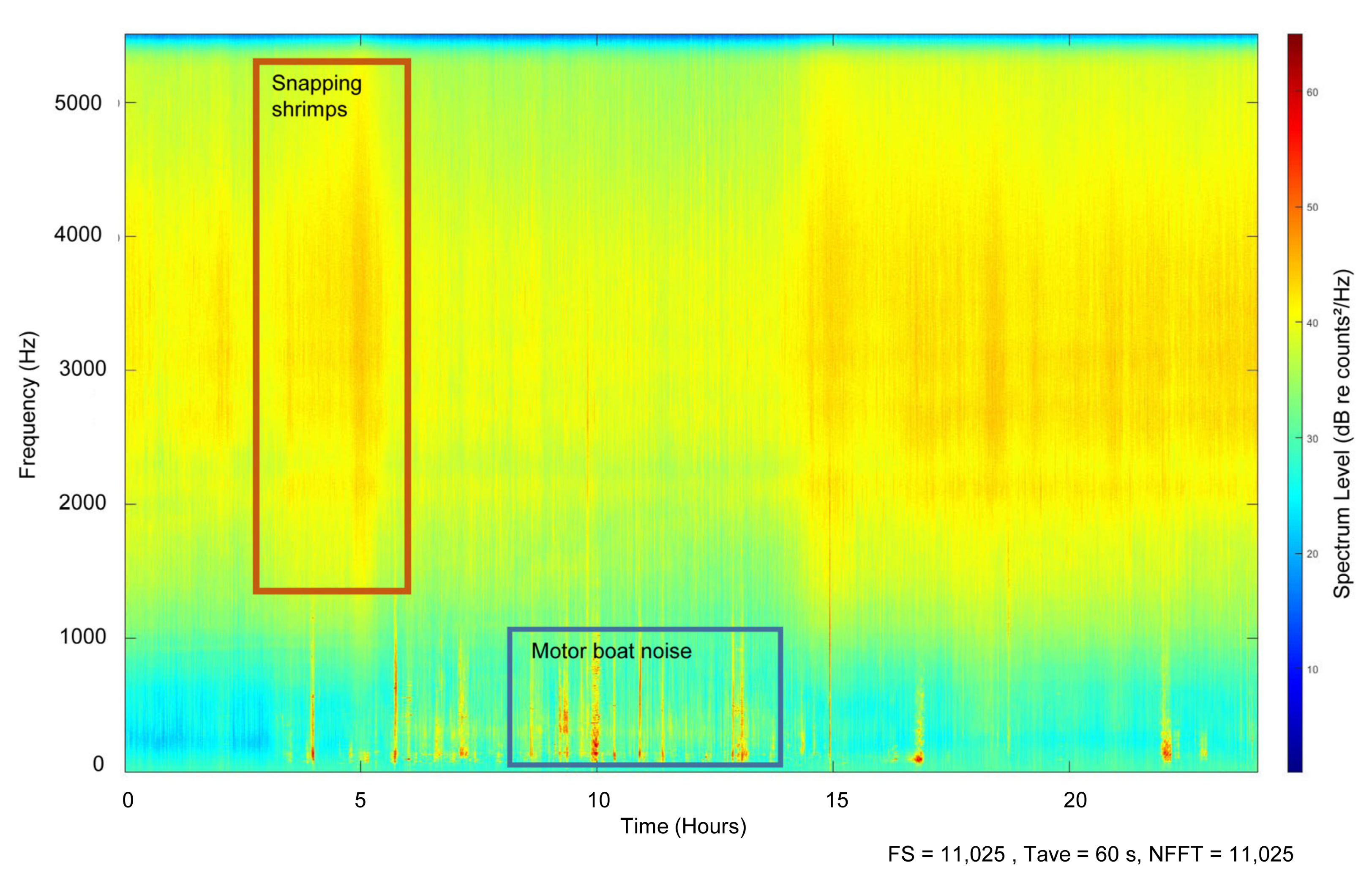
3. Results
3.1. Call Types
3.2. Boat Noise
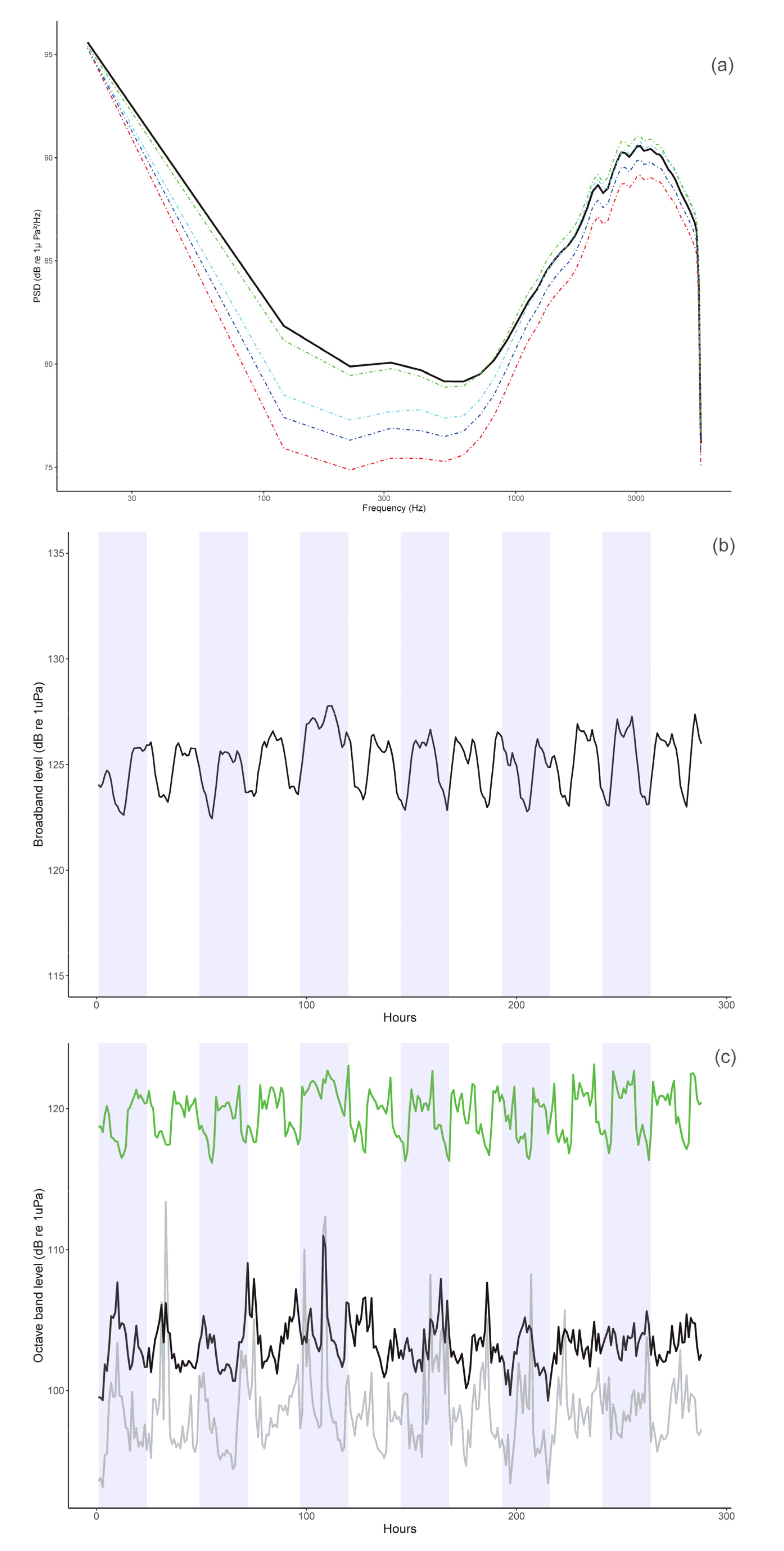
3.3. Soundscape Analysis
4. Discussion
Management Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karamanlidis, A.; Dendrinos, P. Monachus monachus, The IUCN Red List Threat Species. 2015. Available online: https://www.iucnredlist.org/species/13653/117647375#errata (accessed on 22 December 2022).
- Karamanlidis, A.A. MOm/Hellenic Society for the Study and Protection of the Monk Seal, Athens, Greece. 2023; Unpublished work. [Google Scholar]
- Adamantopoulou, S.; Karamanlidis, A.A.; Dendrinos, P.; Gimenez, O. Citizen Science Indicates Significant Range Recovery and Defines New Conservation Priorities for Earth’s Most Endangered Pinniped in Greece. Anim. Conserv. 2022, 26, 115–125. [Google Scholar] [CrossRef]
- Nicolaou, H.; Dendrinos, P.; Marcou, M.; Michaelides, S.; Karamanlidis, A.A. Re-Establishment of the Mediterranean Monk Seal Monachus monachus in Cyprus: Priorities for Conservation. ORYX 2021, 55, 526–528. [Google Scholar] [CrossRef]
- Dendrinos, D.; Karamanlidis, A.A.; Adamantopoulou, S.; Koemtzopoulos, K.; Komninou, A.; Tounta, E. NoLIFE-IP 4 NATURA: Integrated Actions for the Conservation and Management of Natura 2000 Sites, Species, Habitats and Ecosystems in Greece: Deliverable Action A.1: Action Plan for the Mediterranean Monk Seal (Monachus monachus); Report; Hellenic Ministry of Environment and Energy: Athens, Greece, 2020; pp. 1–105. [Google Scholar]
- Charrier, I. Vocal Communication in Otariids and Odobenids. In Ethology and Behavioral Ecology of Otariids and the Odobenid; Campagna, C., Harcourt, R., Eds.; Springer: Cham, Switzerland, 2021; pp. 265–289. [Google Scholar]
- Charrier, I.; Casey, C. Social Communication in Phocids. In Ethology and Behavioral Ecology of Phocids; Costa, D.P., McHuron, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2022; pp. 69–100. ISBN 9783030889234. [Google Scholar]
- Bradbury, J.W.; Vehrencamp, S.L. Principles of Animal Communication; Sinauer Associates, Inc: Sunderland, MA, USA, 2011. [Google Scholar]
- Van Opzeeland, I.; Van Parijs, S.; Bornemann, H.; Frickenhaus, S.; Kindermann, L.; Klinck, H.; Plötz, J.; Boebel, O. Acoustic Ecology of Antarctic Pinnipeds. Mar. Ecol. Prog. Ser. 2010, 414, 267–291. [Google Scholar] [CrossRef]
- Frouin-Mouy, H.; Mouy, X.; Martin, B.; Hannay, D. Underwater Acoustic Behavior of Bearded Seals (Erignathus barbatus) in the Northeastern Chukchi Sea, 2007–2010. Mar. Mammal Sci. 2016, 32, 141–160. [Google Scholar] [CrossRef]
- Mouy, X.; Hannay, D.; Zykov, M.; Martin, B. Tracking of Pacific Walruses in the Chukchi Sea Using a Single Hydrophone. J. Acoust. Soc. Am. 2012, 131, 1349. [Google Scholar] [CrossRef] [PubMed]
- Marcoux, M.; Ferguson, S.H.; Roy, N.; Bedard, J.M.; Simard, Y. Seasonal Marine Mammal Occurrence Detected from Passive Acoustic Monitoring in Scott Inlet, Nunavut, Canada. Polar Biol. 2017, 40, 1127–1138. [Google Scholar] [CrossRef]
- Leroy, E.C.; Royer, J.Y.; Bonnel, J.; Samaran, F. Long-Term and Seasonal Changes of Large Whale Call Frequency in the Southern Indian Ocean. J. Geophys. Res. Ocean. 2018, 123, 8568–8580. [Google Scholar] [CrossRef]
- Moore, S.E.; Stafford, K.M.; Dahlheim, M.E.; Fox, C.G.; Braham, H.W.; Polovina, J.J.; Bain, D.E. Seasonal Variation in Reception of Fin Whale Calls at Five Geographic Areas in the North Pacific. Mar. Mammal Sci. 1998, 14, 617–627. [Google Scholar] [CrossRef]
- Stafford, K.M.; Nieukirk, S.L.; Fox, C.G. Geographic and Seasonal Variation of Blue Whale Calls in the North Pacific. J. Cetacean Res. Manag. 2001, 3, 65–76. [Google Scholar]
- Moore, S.E.; Stafford, K.M.; Mellinger, D.K.; Hildebrand, J.A. Listening for Large Whales in the Offshore Waters of Alaska. Bioscience 2006, 56, 49. [Google Scholar] [CrossRef]
- Zimmer, W.M.X.; Harwood, J.; Tyack, P.L.; Johnson, M.P.; Madsen, P.T. Passive Acoustic Detection of Deep-Diving Beaked Whales. J. Acoust. Soc. Am. 2008, 124, 2823–2832. [Google Scholar] [CrossRef] [PubMed]
- Samaran, F.; Stafford, K.M.; Branch, T.A.; Gedamke, J.; Royer, J.Y.; Dziak, R.P.; Guinet, C. Seasonal and Geographic Variation of Southern Blue Whale Subspecies in the Indian Ocean. PLoS ONE 2013, 8, e71561. [Google Scholar] [CrossRef]
- Stafford, K.M.; Nieukirk, S.L.; Fox, C.G. Low-Frequency Whale Sounds Recorded on Hydrophones Moored in the Eastern Tropical Pacific. J. Acoust. Soc. Am. 1999, 106, 3687–3698. [Google Scholar] [CrossRef]
- Blackwell, S.; Richardson, W. Bowhead Whale (Balaena Mysticetus) Migration and Calling Behaviour in the Alaskan Beaufort Sea, Autumn 2001-04: An Acoustic Localization Study. Arctic 2007, 60, 255–270. [Google Scholar] [CrossRef]
- Duarte, C.M.; Chapuis, L.; Collin, S.P.; Costa, D.P.; Devassy, R.P.; Eguiluz, V.M.; Erbe, C.; Gordon, T.A.C.; Halpern, B.S.; Harding, H.R.; et al. The Soundscape of the Anthropocene Ocean. Science 2021, 371, eaba4658. [Google Scholar] [CrossRef]
- Carroll, A.G.; Przeslawski, R.; Duncan, A.; Gunning, M.; Bruce, B. A Critical Review of the Potential Impacts of Marine Seismic Surveys on Fish & Invertebrates. Mar. Pollut. Bull. 2017, 114, 9–24. [Google Scholar] [PubMed]
- Weilgart, L.S. A Brief Review of Known Effects of Noise on Marine Mammals. Int. J. Comp. Psychol. 2007, 20, 159–168. [Google Scholar] [CrossRef]
- Mikkelsen, L.; Johnson, M.; Wisniewska, D.M.; van Neer, A.; Siebert, U.; Madsen, P.T.; Teilmann, J. Long-Term Sound and Movement Recording Tags to Study Natural Behavior and Reaction to Ship Noise of Seals. Ecol. Evol. 2019, 9, 2588–2601. [Google Scholar] [CrossRef]
- Pichegru, L.; Nyengera, R.; McInnes, A.M.; Pistorius, P. Avoidance of Seismic Survey Activities by Penguins. Sci. Rep. 2017, 7, 16305. [Google Scholar] [CrossRef]
- Pichegru, L.; Vibert, L.; Thiebault, A.; Charrier, I.; Stander, N.; Ludynia, K.; Lewis, M.; Carpenter-Kling, T.; McInnes, A. Maritime Traffic Trends around the Southern Tip of Africa–Did Marine Noise Pollution Contribute to the Local Penguins’ Collapse? Sci. Total Environ. 2022, 849, 157878. [Google Scholar] [CrossRef]
- Shannon, G.; McKenna, M.F.; Angeloni, L.M.; Crooks, K.R.; Fristrup, K.M.; Brown, E.; Warner, K.A.; Nelson, M.D.; White, C.; Briggs, J.; et al. A Synthesis of Two Decades of Research Documenting the Effects of Noise on Wildlife. Biol. Rev. 2016, 91, 982–1005. [Google Scholar] [CrossRef] [PubMed]
- Charrier, I.; Marchesseau, S.; Dendrinos, P.; Tounta, E.; Karamanlidis, A.A. Individual Signatures in the Vocal Repertoire of the Endangered Mediterranean Monk Seal: New Perspectives for Population Monitoring. Endanger. Species Res. 2017, 32, 459–470. [Google Scholar] [CrossRef]
- Muñoz, G.; Karamanlidis, A.A.; Dendrinos, P.; Thomas, J.A. Aerial Vocalizations by Wild and Rehabilitating Mediterranean Monk Seals (Monachus monachus) in Greece. Aquat. Mamm. 2011, 37, 262–279. [Google Scholar] [CrossRef]
- Marchessaux, D.; Muller, N. Le Phoque Moine, Monachus Monachus. Distribution, Statut et Biologie Sur La Côte Saharienne. Sci. Rep. Port Cros Nat. Park 1987, 13, 25–84. [Google Scholar]
- Mpougas, E.; Waggitt, J.J.; Dendrinos, P.; Adamantopoulou, S.; Karamanlidis, A.A. Mediterranean Monk Seal (Monachus monachus) Behavior at Sea and Interactions with Boat Traffic: Implications for the Conservation of the Species in Greece. Aquat. Mamm. 2019, 45, 419–424. [Google Scholar] [CrossRef]
- Sills, J.M.; Parnell, K.; Ruscher, B.; Lew, C.; Kendall, T.L.; Reichmuth, C. Underwater Hearing and Communication in the Endangered Hawaiian Monk Seal Neomonachus Schauinslandi. Endanger. Species Res. 2021, 44, 61–78. [Google Scholar] [CrossRef]
- Dendrinos, P. Contribution to the Study of the Mediterranean Monk Seal’s (Monachus monachus) Ecology and Biology at the Island Complex of Northern Sporades, Greece; National and Kapodistrian University of Athens: Athens, Greece, 2011. [Google Scholar]
- Dendrinos, P.; Adamantopoulou, S.; Koemtzopoulos, K.; Mpatzios, P.; Paxinos, O.; Tounta, E.; Tsiakalos, D.; Karamanlidis, A.A. Anecdotal Observations of Open Beach Use by Female Mediterranean Monk Seals (Monachus monachus) and Their Pups in Greece: Implications for Conservation. Aquat. Mamm. 2022, 48, 602–609. [Google Scholar] [CrossRef]
- Karamanlidis, A.A.; Dendrinos, P.; de Larrinoa, P.F.; Gücü, A.C.; Johnson, W.M.; Kiraç, C.O.; Pires, R. The Mediterranean Monk Seal Monachus monachus: Status, Biology, Threats, and Conservation Priorities. Mamm. Rev. 2016, 46, 92–105. [Google Scholar] [CrossRef]
- Dendrinos, P.; Karamanlidis, A.A.A.; Kotomatas, S.; Paravas, V.; Adamantopoulou, S. Report of a New Mediterranean Monk Seal (Monachus monachus) Breeding Colony in the Aegean Sea, Greece. Aquat. Mamm. 2008, 34, 355–361. [Google Scholar] [CrossRef]
- Sueur, J.; Aubin, T.; Simonis, C. Equipment Review: Seewave, a Free Modular Tool for Sound Analysis and Synthesis. Bioacoustics 2008, 18, 213–226. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing. Available online: https://alem.lu/wp-content/uploads/2019/12/ALEM-Team-2020.pdf (accessed on 4 January 2022).
- Thiebault, A.; Charrier, I.; Pistorius, P.; Aubin, T. At Sea Vocal Repertoire of a Foraging Seabird. J. Avian Biol. 2019, 50, 1–14. [Google Scholar] [CrossRef]
- Indeck, K.L.; Girola, E.; Torterotot, M.; Noad, M.J.; Dunlop, R.A. Adult Female-Calf Acoustic Communication Signals in Migrating East Australian Humpback Whales. Bioacoustics 2021, 30, 341–365. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and Regression by RandomForest. R News 2002, 2, 18–22. [Google Scholar]
- Thomas, J.A.; Kuechle, V.B. Quantitative Analysis of Weddell Seal (Leptonychotes weddelli) Underwater Vocalizations at Mcmurdo Sound, Antarctica. J. Acoust. Soc. Am. 1982, 72, 1730–1738. [Google Scholar] [CrossRef] [PubMed]
- Asselin, S.; Hammill, M.O.; Barrette, C. Underwater Vocalizations of Ice Breeding Grey Seals. Can. J. Zool. 1993, 71, 2211–2219. [Google Scholar] [CrossRef]
- Hocking, D.P.; Evans, A.R.; Burville, B.; Park, T.; Parker, W.M.G.; Marx, F.G. Percussive Underwater Signaling in Wild Gray Seals. Mar. Mammal Sci. 2020, 5, 1–5. [Google Scholar] [CrossRef]
- Sjare, B.; Stirling, I.; Spencer, C. Structural Variation in the Songs of Atlantic Walruses Breeding in the Canadian High Arctic. Aquat. Mamm. 2003, 29, 297–318. [Google Scholar] [CrossRef]
- Oswald, J.N.; Walmsley, S.F.; Casey, C.; Fregosi, S.; Southall, B.; Janik, V.M. Species Information in Whistle Frequency Modulation Patterns of Common Dolphins. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20210046. [Google Scholar] [CrossRef]
- Garcia, H.A.; Zhu, C.; Schinault, M.E.; Kaplan, A.I.; Handegard, N.O.; Godø, O.R.; Ahonen, H.; Makris, N.C.; Wang, D.; Huang, W.; et al. Temporal-Spatial, Spectral, and Source Level Distributions of Fin Whale Vocalizations in the Norwegian Sea Observed with a Coherent Hydrophone Array. ICES J. Mar. Sci. 2019, 76, 268–283. [Google Scholar] [CrossRef]
- Weirathmueller, M.J.; Stafford, K.M.; Wilcock, W.S.D.; Hilmo, R.S.; Dziak, R.P.; Tréhu, A.M. Spatial and Temporal Trends in Fin Whale Vocalizations Recorded in the NE Pacific Ocean between 2003–2013. PLoS ONE 2017, 12, e0186127. [Google Scholar] [CrossRef]
- Jones, B.; Zapetis, M.; Samuelson, M.M.; Ridgway, S. Sounds Produced by Bottlenose Dolphins (Tursiops): A Review of the Defining Characteristics and Acoustic Criteria of the Dolphin Vocal Repertoire. Bioacoustics 2020, 29, 399–440. [Google Scholar] [CrossRef]
- Insley, S.; Phillips, A.; Charrier, I. A Review of Social Recognition in Pinnipeds. Aquat. Mamm. 2003, 29, 181–201. [Google Scholar] [CrossRef]
- Rolland, R.M.; Parks, S.E.; Hunt, K.E.; Castellote, M.; Corkeron, P.J.; Nowacek, D.P.; Wasser, S.K.; Kraus, S.D. Evidence That Ship Noise Increases Stress in Right Whales. Proc. R. Soc. B Biol. Sci. 2012, 279, 2363–2368. [Google Scholar] [CrossRef]
- Atkinson, S.; Crocker, D.; Houser, D.; Mashburn, K. Stress Physiology in Marine Mammals: How Well Do They Fit the Terrestrial Model? J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2015, 185, 463–486. [Google Scholar] [CrossRef]
- Romero, M.L.; Butler, L.K. Endocrinology of Stress. Int. J. Comp. Psychol. 2007, 20, 89–95. [Google Scholar] [CrossRef]
- Nowacek, D.P.; Christiansen, F.; Bejder, L.; Goldbogen, J.A.; Friedlaender, A.S. Studying Cetacean Behaviour: New Technological Approaches and Conservation Applications. Anim. Behav. 2016, 120, 235–244. [Google Scholar] [CrossRef]
- Saloma, A.; Ratsimbazafindranahaka, M.N.; Martin, M.; Andrianarimisa, A.; Huetz, C.; Adam, O.; Charrier, I. Social Calls in Humpback Whale Mother-Calf Groups off Sainte Marie Breeding Ground (Madagascar, Indian Ocean). PeerJ 2022, 10, e13785. [Google Scholar] [CrossRef] [PubMed]
- Saddler, M.R.; Bocconcelli, A.; Hickmott, L.S.; Chiang, G.; Landea-Briones, R.; Bahamonde, P.A.; Howes, G.; Segre, P.S.; Sayigh, L.S. Characterizing Chilean Blue Whale Vocalizations with DTAGs: A Test of Using Tag Accelerometers for Caller Identification. J. Exp. Biol. 2017, 220, 4119–4129. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Harmonic Calls | dur (s) | f0 (Hz) | excF (Hz) | Fmax (Hz) | Q25 (Hz) | Q50 (Hz) | Q75 (Hz) |
|---|---|---|---|---|---|---|---|
| Bark (n = 294) | 0.418 ± 0.262 | 137 ± 37 | 45 ± 27 | 312 ± 190 | 275 ± 83 | 450 ± 134 | 687 ± 148 |
| Croak (n = 32) | 0.460 ± 0.147 | 96 ± 40 | 43 ± 29 | 282 ± 146 | 238 ± 86 | 366 ± 162 | 622 ± 406 |
| Cry (n = 24) | 1.386 ± 1.074 | 290 ± 150 | 95 ± 77 | 360 ± 170 | 272 ± 106 | 433 ± 118 | 699 ± 129 |
| Gloo (n = 76) | 0.350 ± 0.359 | 112 ± 29 | 74 ± 43 | 147 ± 79 | 153 ± 53 | 283 ± 109 | 490 ± 147 |
| Gloogloo (n = 39) | 0.555 ± 0.562 | 116 ± 28 | -- | 193 ± 80 | 171 ± 37 | 288 ± 61 | 503 ± 119 |
| Groan (n = 78) | 0.500 ± 0.247 | 79 ± 36 | -- | 174 ± 87 | 178 ± 58 | 287 ± 82 | 507 ± 289 |
| Scream (n = 19) | 0.408 ± 0.277 | 1207 ± 444 | 150 ± 111 | 1207 ± 444 | -- | -- | -- |
| Whines (n = 47) | 0.663 ± 0.313 | 134 ± 45 | 51 ± 30 | 143 ± 41 | -- | -- | -- |
| Whoo (n = 65) | 0.207 ± 0.068 | 439 ± 154 | 73 ± 48 | 461 ± 190 | -- | -- | -- |
| Wop (n = 23) | 0.229 ± 0.069 | 297 ± 70 | 68 ± 30 | 406 ± 145 | 316 ± 59 | 486 ± 110 | 699 ± 94 |
| Wom (n = 42) | 0.303 ± 0.100 | 95 ± 30 | 54 ± 33 | 100 ± 44 | 117 ± 39 | 288 ± 157 | 632 ± 191 |
| Noisy calls | dur (s) | Fmax (Hz) | Q25 (Hz) | Q50 (Hz) | Q75 (Hz) | ||
| Growl (n = 442) | 1.136 ± 0.616 | 648 ± 412 | 542 ± 249 | 920 ± 388 | 1399 ± 662 | ||
| Hiccup (n = 210) | 0.177 ± 0.042 | 395 ± 109 | 353 ± 82 | 422 ± 99 | 567 ± 120 | ||
| Squeak (n = 43) | 0.155 ± 0.081 | 557 ± 147 | 422 ± 92 | 556 ± 92 | 690 ± 105 | ||
| Pulsative calls | dur (s) | Fmax (Hz) | Q25 (Hz) | Q50 (Hz) | Q75 (Hz) | PR (Hz) | |
| Clap (n = 13) | 5.103 ± 1.619 | -- | -- | -- | -- | 1.70 ± 0.16 | |
| Knock (n = 226) | 1.922 ± 1.828 | 699 ± 441 | -- | -- | -- | 6.81 ± 1.43 | |
| Rumble (n = 23) | 1.062 ± 0.434 | 205 ± 65 | 215 ± 50 | 347 ± 97 | 571 ± 142 | 16.31 ± 3.5 |
| Bark | Croak | Cry | Gloo | Gloogloo | Groan | Scream | Whine | Whoo | Wom | Wop | Classification Error | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| bark | 271 | 2 | 2 | 11 | 1 | 0 | 0 | 0 | 0 | 4 | 3 | 0.08 |
| croak | 15 | 14 | 0 | 2 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0.56 |
| cry | 9 | 0 | 12 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 2 | 0.50 |
| gloo | 17 | 2 | 0 | 44 | 2 | 2 | 0 | 0 | 0 | 9 | 0 | 0.42 |
| gloogloo | 7 | 0 | 0 | 1 | 19 | 11 | 0 | 0 | 0 | 1 | 0 | 0.51 |
| groan | 9 | 0 | 0 | 0 | 8 | 61 | 0 | 0 | 0 | 0 | 0 | 0.22 |
| scream | 0 | 0 | 0 | 0 | 0 | 0 | 15 | 0 | 4 | 0 | 0 | 0.21 |
| whine | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 46 | 1 | 0 | 0 | 0.02 |
| whoo | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 2 | 62 | 0 | 0 | 0.05 |
| wom | 6 | 0 | 0 | 10 | 0 | 2 | 0 | 0 | 0 | 24 | 0 | 0.43 |
| wop | 4 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 18 | 0.22 |
| Growl | Hiccup | Squeak | Classification Error | |
|---|---|---|---|---|
| growl | 441 | 1 | 0 | 0.002 |
| hiccup | 0 | 199 | 11 | 0.052 |
| squeak | 1 | 19 | 23 | 0.465 |
| Clap | Knock | Rumble | Classification Error | |
|---|---|---|---|---|
| clap | 13 | 0 | 0 | 0 |
| knock | 0 | 222 | 2 | 0.009 |
| rumble | 0 | 2 | 21 | 0.086 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charrier, I.; Huetz, C.; Prevost, L.; Dendrinos, P.; Karamanlidis, A.A. First Description of the Underwater Sounds in the Mediterranean Monk Seal Monachus monachus in Greece: Towards Establishing a Vocal Repertoire. Animals 2023, 13, 1048. https://doi.org/10.3390/ani13061048
Charrier I, Huetz C, Prevost L, Dendrinos P, Karamanlidis AA. First Description of the Underwater Sounds in the Mediterranean Monk Seal Monachus monachus in Greece: Towards Establishing a Vocal Repertoire. Animals. 2023; 13(6):1048. https://doi.org/10.3390/ani13061048
Chicago/Turabian StyleCharrier, Isabelle, Chloé Huetz, Léa Prevost, Panagiotis Dendrinos, and Alexandros A. Karamanlidis. 2023. "First Description of the Underwater Sounds in the Mediterranean Monk Seal Monachus monachus in Greece: Towards Establishing a Vocal Repertoire" Animals 13, no. 6: 1048. https://doi.org/10.3390/ani13061048
APA StyleCharrier, I., Huetz, C., Prevost, L., Dendrinos, P., & Karamanlidis, A. A. (2023). First Description of the Underwater Sounds in the Mediterranean Monk Seal Monachus monachus in Greece: Towards Establishing a Vocal Repertoire. Animals, 13(6), 1048. https://doi.org/10.3390/ani13061048