Simple Summary
Cold stress is considered an environmental stress and is an important managemental factor in regions where winter temperatures drop below 18 °C. Although a low temperature lowered the productive performance of laying hens, there are still unanswered questions as to the physiological changes that occur when they are exposed to low temperature, especially changes in antioxidants and stress indicators, which prompted us to set up the current study. Laying hens reared at low temperature (12 ± 4.5 °C) showed impaired laying performance compared with laying hens reared at normal temperature (24 ± 3 °C). However, eggshell color was intensified in laying hens exposed to low temperature. During the early stage of exposure to low-temperature stress, malondialdehyde levels in the egg yolk increased in laying hens raised at low temperature vs. normal temperature. However, yolk corticosterone, an indicator of stress responses, remained unchanged. Low temperature had an impact on total cholesterol and triglyceride levels in serum increases in laying hens reared at low temperature. In essence, low temperature in laying hens altered antioxidant systems and lipid metabolism without inducing stress responses.
Abstract
The present study investigated the effect of low temperature on laying performance, egg quality, body temperature, yolk malondialdehyde, yolk corticosterone, and serum biochemistry in laying hens. A total of 40 laying hens (Hy-Line Brown) aged 36 weeks were housed in one of two environmental chambers kept at 12 ± 4.5 °C (low temperature) or 24 ± 3 °C (normal temperature) for 4 weeks. Low vs. normal temperature significantly increased (p < 0.05) live body weight, feed intake, and feed conversion ratio in laying hens. Skin surface temperature, but not rectal temperature, was decreased in laying hens exposed to low vs. normal temperature. Hens exposed to low temperature laid an intense eggshell color compared with those raised in a normal temperature. Malondialdehyde concentrations in yolk were increased in low-temperature-exposed laying hens compared with those at normal temperature conditions, but this effect was only noted on day 7, post the low-temperature exposure (p = 0.04). Finally, low vs. normal temperature increased the concentrations of total cholesterol and triglyceride in serum. Collectively, this study indicates that exposure to low temperature in laying hens initially disrupted antioxidant system and altered lipid metabolism in laying hens without inducing stress responses.
1. Introduction
The future of poultry husbandry will be focused on improved animal welfare, with less reliance on preventative medical interventions. In the poultry husbandry environment, animals are likely to encounter various kinds of simultaneous environmental stressors. In emphasizing the latter, authors have tried to understand the physiological and stress responses of laying hens exposed to various stressors, including stocking density, transport, or ambient temperature [1]. It is generally understood that the thermal neutral zone for the optimal metabolic and productive activity of poultry ranges from around 18 to 23.9 °C [2] with a relative humidity between 50% and 70% during the laying period [3]. Heat stress is well-explored and acknowledged when laying hens are raised beyond the thermal neutral zone [1]. In contrast to heat stress, our understanding of the impact of low temperature (i.e., cold stress) on laying hens is still limited.
It has been reported that the ambient environmental temperature below 16 °C results in negative effects on poultry production performance, such as egg production, egg mass, and egg quality [4,5,6]. Cold stress is considered an environmental stress and can be considered an important managemental factor in regions where environmental temperatures during the winter periods drop below 18 °C. Earlier studies reported that cold exposure could influence the antioxidant and immune system functions of the host [7,8,9]. During low-temperature exposure, the hypothalamus, which acts as the control center for temperature regulation, triggers homeostatic mechanisms to maintain body temperature by generating heat. Also, upon experiencing stress, the activation of the hypothalamic–pituitary–adrenal (HPA) axis leads to the elevated secretion of corticosterone (CORT) from the adrenal glands. This process is intended to stimulate gluconeogenesis within various tissues and lipolysis in adipose tissue, thereby augmenting both glucose accessibility and the metabolic rate of energy [10].
In general, low temperature, as a physical environmental stressor, is known to in-crease feed intake to meet energy requirements [11]. Low ambient temperatures also result in decreased insulin and increased corticosterone (CORT) concentrations in the serum of laying hens [12,13]. It is well-known that insulin and CORT are influenced by temperature and these hormones play an important role in controlling nutrient metabolism [14,15]. In addition, a low temperature can disrupt the balance in an antioxidant system, leading to oxidative damage of several tissues by altering the enzymatic and non-enzymatic antioxidant status, lipid peroxidation, antioxidant vitamins, and reactive oxygen species production [16]. It has been reported that malondialdehyde (MDA) was increased in chickens exposed to low temperature, when compared to normal-temperature-reared counterparts [17,18]. Nonetheless, studies of the effects of low temperature on the egg quality, body temperature (rectal and surface), MDA in yolk, and CORT in yolk of laying hens that engage on a weekly basis are scarce, which prompted us to set up the current experiment. Thus, this present study aimed to address the biological and physiological responses of laying hens exposed to low ambient temperature.
2. Materials and Methods
2.1. Birds, Diets, and Experimental Design
A total of forty 36-week-old Hy-Line Brown laying hens with the average body weight of 1871 ± 123 g were used in this study. The birds were housed (one bird/cage) in one of two experimental chambers, which had one tier of 20 cages, 1 m high from the floor. Each cage, measuring 41 × 37 × 40 cm (length × depth × height), had nipples and a trough-type feeder. Two adjacent cages sharing one feeder were considered a replicate (n = 2/replicate, 10 replicates/treatment). The sample size (n = 10 per treatment) was calculated based on a Type I error of 5%, 80% statistical power, an effect size of 7.6%, and a coefficient of variation of 6%. In addition, to reduce variation within replicates, 2 laying hens per replicate were used. Each hen was provided with a 1517 cm2 floor space. All birds were initially adapted to the chambers for 7 days at an ambient temperature of 24 °C with a relative humidity of 40 ± 4% and a lighting program of 16L/8D. After adaptation periods, hens were exposed to one of two temperature regimes (see Section 2.2 for the temperature regimes) for 28 days. Corn, soybean meal, and dried distillers grains with a solubles-based commercial layer diet were provided (Table 1), with feed and water supplied ad libitum.

Table 1.
Ingredients and chemical composition of the basal diet (%, as-fed basis).
2.2. Temperature Treatments and Temperature Monitor
Two environmental chambers were continuously set at 12 ± 4.5 °C (i.e., low temperature) and 24 ± 3.5 °C (i.e., normal temperature) and the average relative humidity inside the chambers was maintained at 40 ± 4%. Each chamber was equipped with a heater (MCP-300; MAXCON Co., Bucheon, Republic of Korea), air-conditioner (AR07J5174HA, SAMSUNG, Suwon, Republic of Korea), humidifier (MH-601A; mtechwin Co., Gimhae, Republic of Korea), dehumidifier (NED-062P; NAWOOEL Co., Gimpo, Republic of Korea), and the main controller panel. Temperature and humidity loggers (MHT-381SD; Lutron Electronic Enterprise Co., Taipei, Taiwan) were also installed to monitor the temperature and relative humidity at 10 min intervals throughout the experiment.
2.3. Measurements of Rectal Temperature and Skin Surface Temperature
On days 7, 14, 21, and 28 following temperature treatment, the rectal and skin surface temperature of laying hens was measured. One hen per replicate was randomly selected and measured by inserting a rectal thermometer to a depth of 3 cm into the rectum. Skin surface temperature was measured at three different sites (i.e., head, chest, and leg) using a thermal imaging camera (Cat® S60: equipped with an FLIR™ Lepton, FLIR Systems Inc., Wilsonville, OR, USA), as previously described [19]. The hens were handled by wearing latex gloves, to avoid the influences of heat and moisture of the hands on the temperature of the feathers.
2.4. Egg Production and Egg Quality
Body weight and feed intake were measured weekly. Eggs were collected daily and weighed per replicate to calculate hen-day egg production. Feed conversion ratio was expressed as kg of feed consumed per kg of eggs produced. Eggs (3 eggs per replicate) collected on the preceding 3 consecutive days, at 7, 14, 21, and 28 days following the temperature treatment, were used to measure egg quality with a digital egg tester (DET-6000, Navel, Kyoto, Japan). Yolk color intensity was evaluated on a scale between 1 and 16, with 1 being a very pale yellow and 16 being a dark orange. Eggshell color was measured using an egg multi-tester made by TSS (QCR, Technical Services and Supplies Ltd., Yolk, UK). Eggshell color intensity was evaluated on a scale between 0 and 100, with 0 being a darkness and 100 being a lightness.
2.5. Malondialdehyde in Yolk Samples
Eggs were collected to measure yolk MDA concentrations at days 7, 14, 21, and 28. The eggs were cracked open, and the yolks were separated from the albumin by gently rolling the yolk on filter paper. Three yolks were pooled and homogenized. The yolk MDA was measured using the OxiSelect TBARS Assay kit (Cell Biolabs Inc., San Diego, CA, USA).
2.6. Corticosterone in Yolk Samples
Yolk CORT concentrations were measured at 7, 14, 21, and 28 days, and 4 g of pooled yolk were vortexed with an equal volume of PBS. Then, 1 mL of the yolk suspension was mixed with an equal volume of ethanol, incubated at 37 °C for 1 h, and subsequently centrifuged. The 50 μL of supernatants were mixed with 50 μL of ethanol and 50 μL of PBS solution, and these mixtures were analyzed with a CORT ELISA kit (Enzo life science Inc., ADI-901-097, Farmingdale, NY, USA), as previously described [20,21].
2.7. Serum Parameters
At 28 days, one bird per replicate was selected to collect blood from a wing vein. Serum was separated by centrifugation at 200 g for 15 min and stored at −20 °C until the analysis. Serum samples were analyzed by using an automatic blood chemical analyzer (Film DRI CHEM 7000i, Fuji film, Tokyo, Japan) to measure for total cholesterol, triglyceride, high-density lipoprotein cholesterol, glutamic oxaloacetic transaminase, glutamic pyruvic transaminase, and uric acid.
2.8. Statistical Analysis
Two adjacent cages were considered an experimental unit. All data were analyzed using Student’s t-test procedure of SAS (SAS Institute Inc., Cary, NC, USA). Results were presented as least square means and standard deviation. Differences were considered significant at p < 0.05.
3. Results
Ambient temperature ranged from 21 to 27 °C (normal temperature) and from 7.5 to 16.5 °C (low temperature) in the normal- and low-temperature chambers (Figure 1).
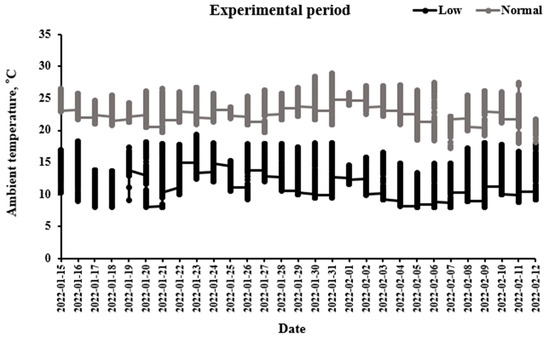
Figure 1.
Ambient temperatures in chambers were recorded at an interval of 10 min with a thermometer.
Low temperature increased (p < 0.05) the final body weight, feed intake, and FCR of laying hens compared with those raised at a normal temperature (Table 2).
Table 2.
The effect of cold stress on production performance 1.
However, egg weight, egg production and egg mass were not affected (p > 0.05) by temperature regimes. Rectal temperature during the experimental period was maintained from 41.27 to 41.58 °C and was not different between temperature treatment groups (Table 3).
Table 3.
The effect of cold stress on rectal temperature (°C) 1.
Table 4 presents the skin surface temperature at 7, 14, 21, and 28 days. The laying hens exposed to low temperature at all ages had lower skin (i.e., head, breast, and leg regions) surface temperature compared with those raised at normal temperature (p < 0.05). Skin surface temperature between low- vs. normal-temperature treatments was 5.7 to 12.2 °C lower in the breast surface area, followed by the legs and the head areas.
Table 4.
The effect of cold stress on skin surface temperature (°C) 1.
Low temperature significantly increased (p < 0.05) eggshell color at days 7 and 28, compared with normal-temperature-raised laying hens (Table 5). However, yolk color, the Haugh unit, shell strength, and shell thickness were not affected by temperature treatments on all days (p > 0.05).
Table 5.
The effect of cold stress on egg quality 1.
Yolk MDA concentrations were higher in laying hens exposed to low temperature, compared with those in normal temperature at 7 days (Table 6). However, low-temperature-induced decreases in yolk MDA concentrations were not noted (p > 0.05) at 14, 21, and 28 days.
Table 6.
The effect of cold stress on malondialdehyde in egg yolk (µM) 1.
The CORT concentrations were not different in laying hens exposed to low and normal temperature (Table 7). Low vs. normal temperature increased (p < 0.05) the concentrations of total cholesterol and triglycerides, but did not affect the concentrations of high-density lipoprotein cholesterol, glutamic oxaloacetic transaminase, glutamic pyruvic transaminase, and uric acid in serum samples (Table 8; p > 0.05).
Table 7.
The effect of cold stress on corticosterone in egg yolk (ng/g) 1.
Table 8.
The effect of cold stress on serum biological parameters in serum 1.
4. Discussion
The present study showed that low vs. normal ambient temperature increased final body weight, feed intake, and FCR in laying hens. This finding is well-established because chickens, as a homeothermic animal, are unable to warm themselves and so overfeed to compensate for heat loss to the environment when they are exposed to low temperature (e.g., 18 °C) [22]. As ambient temperature shifted from 30 to 18 °C, the abdominal fat weight, abdominal fat rate, and subcutaneous fat thickness linearly increased, favoring body fat deposition [23]. For these reasons, the present study indicates that heavier body weight is due to increases in feed intake and concomitant body fat deposition, leading to a higher FCR in laying hens exposed to low vs. normal temperature. It is well-documented that low temperatures stimulate feed intakes in laying hens [6,24], broiler chickens [17,25], quails [4,26] and turkeys [27].
In the present study, rectal temperature at all ages was not altered in birds exposed to a low temperature or those at normal temperature, which agrees with previous studies with turkeys [28] and laying hens [3]. These results suggest that body heat exchange (e.g., production and loss) does not change above or below the normal rectal temperature range when the homeostasis of warm-blooded animals is maintained [29]. On the other hand, skin surface temperature, an important evaluable parameter, quickly shifts in response to environmental changes and serves as an indicator of alterations in peripheral blood flow and heat exchange [30]. Skin surface (e.g., head, chest, and leg) temperatures were significantly decreased in laying hens raised at low temperature, compared with those at normal temperature, during the whole experiment period. In line with our finding, lower skin surface temperatures in broiler chickens [31] and laying hens [29] have been observed when they were exposed to low vs. normal ambient temperature. It is understood as a way of preventing the loss of heat by cold-induced peripheral vasoconstriction, which reduces blood circulation to the body surface [31]. Additionally, the temperatures of the head and legs were kept higher than that of the chest. It is speculated that this is because the head and legs are featherless body surfaces, compared with the breast body surface, which is covered by feathers [30].
We found that yolk color and eggshell strength were not altered in laying hens raised at a low temperature compared with normal-temperature-raised laying hens, although the former ate more. In contrast to our belief [24,32], our finding indicates that hens exposed to low temperatures might not use excess feed-origin carotenoids and essential nutrients (e.g., amino acids, Ca, Mg) to intensify yolk and to increase eggshell strength. It is of interest that low-temperature-raised hens laid more intensified eggshell colors than hens raised in normal ambient temperature. It is known that the endogenously synthesized protoporphyrin IX is the major pigment in brown eggshell, and that stress factors, including stocking density, fear, or molting, often deteriorate eggshell color pigmentation in brown laying hens [33]. As the pigment is known to be synthesized in the shell gland of the oviduct, it seems that low temperature per se may stimulate the synthetic process of pigment or effectively deposit it on the shell layers. Further studies are warranted to reveal the underlying mechanisms that will show how low temperature affects shell color deposition in laying hens.
It is reported that cold stress causes tissue damage [22] via increased metabolic rate, which demands tissue requirement for oxygen in birds [34]. We found that yolk MDA levels were elevated in low- vs. normal-temperature treatments, but this effect was only noted at 7 days post temperature exposure. Rahmani et al. [17] reported that broiler chickens exposed to 15 °C vs. 22 °C had increased plasma MDA concentrations. In addition, MDA levels in plasma and liver samples were significantly higher in broiler chickens in the low temperature group (from 10 to 15 °C) at 21 and 42 days [18].
In contrast to yolk MDA alterations caused by low temperature, no differences in yolk CORT were noted between low-temperature and normal-condition groups. Exposure of chickens to stress increases the secretion of CORT, a major stress hormone in chickens, via the activation of the hypothalamic pituitary adrenal axis [35]. In earlier studies, the levels of CORT in the albumen and yolk of chicken eggs were used as a non-invasive method to measure stress [20,36,37]. The substances or hormone concentrations in the blood during the egg’s formation phase are deposited in the yolk composition. Hence, if there is an increase in plasma CORT, it would consequently be transferred into the eggs [36]. Also, catching animals for blood sampling can be stressful and it also causes an increase in CORT [38]–therefore, egg yolks were used for an analysis of CORT concentrations in this study. Conflicted results have been reported regarding plasma CORT in response to low ambient temperature. With various treatments (durations of cold exposure, temperature, and age), increased plasma levels of CORT were reported in young broiler chickens (19 ± 1 °C, 6 h/day, from the first to the seventh day of life) [39], male turkeys [40], chicks (1.2 °C for 3 h) [41], and laying hens (0 °C for 72 h) [42]. On the other hand, Hangalapura et al. [43] reported that low temperature (10.4 ± 0.5 °C) decreased plasma CORT in laying hens and Hu and Cheng [44] reported no effect of low temperature on the CORT level in laying hens. It is not clear at this stage why low temperature failed to affect yolk CORT levels in this study, although the information on the effect of low temperature in yolk CORT is scarce. The unaltered CORT could be due to the negative feedback of CORT on the hypothalamus axis with chronic low-temperature exposure, resulting in an inhibition of adrenocorticotropin secretion [45]. Similarly, previous studies have reported that consistently increased CORT is not commonly seen during chronic stress, possibly due to the negative feedback of CORT on the hypothalamus axis [43,44,46].
The finding that low temperature increased the concentrations of total cholesterol and triglyceride is in agreement with the results of [17,47]. This finding indicates that a low-temperature-induced increase in feed intake, coupled with cold stress response in chickens, could stimulate hepatic lipogenic and hypercholesterolemic metabolic pathways [48,49]. On the other hand, glutamic oxaloacetic transaminase and glutamic pyruvic transaminase were not altered by low temperatures, indicating the maintenance of the hepatic function.
5. Conclusions
In conclusion, low vs. normal ambient temperature stimulated feed intake and in-creased body weight and FCR. Eggshell color was intensified in laying hens exposed to low temperature. Low temperature elevated skin surface temperature without affecting rectal temperature. Finally, low temperature did not influence stress responses, as manifested by constant yolk CORT concentrations, but altered MDA, total cholesterol, and triglyceride levels. Taken together, our study indicates that the exposure of laying hens to low temperature disrupted the antioxidant system, especially at an early stage of exposure, and altered lipid metabolism (i.e., total cholesterol and triglyceride) without inducing stress responses. The low-temperature-mediated increase in eggshell color seen in this study warrants further study.
Author Contributions
Conceptualization, D.-H.K. and K.-W.L.; methodology, D.-H.K.; formal analysis, J.P., B.-Y.K. and J.-Y.S.; investigation, D.-H.K., J.P., B.-Y.K. and J.-Y.S.; resources, K.-W.L.; writing—original draft preparation, D.-H.K.; writing—review and editing, D.-H.K. and K.-W.L.; project administration, K.-W.L.; funding acquisition, K.-W.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee of Konkuk University (KU22031).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated or analyzed during the current study are not publicly available but are available from the corresponding author at reasonable request.
Acknowledgments
This paper was supported by Konkuk University in 2023.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tilbrook, A.J.; Fisher, A.D. Stress, health and the welfare of laying hens. Anim. Prod. Sci. 2020, 61, 931–943. [Google Scholar] [CrossRef]
- Durmuş, I.; Kamanli, S. Effects of cold and heat stress on egg quality traits of a newly developed native hybrid layer. Turk. Tarim. Gida. Bilim. Teknol. Derg. 2015, 3, 444–447. [Google Scholar] [CrossRef]
- Kamanli, S.; Durmuş, I.; Yalçın, S.; Yıldırım, U.; Meral, Ö. Effect of prenatal temperature conditioning of laying hen embryos: Hatching, live performance and response to heat and cold stress during laying period. J. Therm. Biol. 2015, 51, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Sahin, N.; Sahin, K.; Onderci, M. Vitamin E and selenium supplementation to alleviate cold-stress-associated deterioration in egg quality and egg yolk mineral concentrations of Japanese quails. Biol. Trace. Elem. Res. 2003, 96, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Torki, M.; Akbari, M.; Kaviani, K. Single and combined effects of zinc and cinnamon essential oil in diet on productive performance, egg quality traits, and blood parameters of laying hens reared under cold stress condition. Int. J. Biometeorol. 2015, 59, 1169–1177. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Tong, Q.; Shi, Z.; Zheng, W.; Wang, Y.; Li, B.; Yan, G. Effects of cold stress and ammonia concentration on productive performance and egg quality traits of laying hens. Animals 2020, 10, 2252. [Google Scholar] [CrossRef] [PubMed]
- Hangalapura, B.N.; Nieuwland, M.G.; de Vries, R.G.; Heetkamp, M.J.; van den Brand, H.; Kemp, B.; Parmentier, H.K. Effects of cold stress on immune responses and body weight of chicken lines divergently selected for antibody responses to sheep red blood cells. Poult. Sci. 2003, 82, 1692–1700. [Google Scholar] [CrossRef]
- Hangalapura, B.N.; Nieuwland, M.G.B.; de Vries Reilingh, G.; Van Den Brand, H.; Kemp, B.; Parmentier, H.K. Durations of cold stress modulates overall immunity of chicken lines divergently selected for antibody responses. Poult. Sci. 2004, 83, 765–775. [Google Scholar] [CrossRef]
- Hangalapura, B.N.; Nieuwland, M.G.B.; Buyse, J.; Kemp, B.; Parmentier, H.K. Effect of duration of cold stress on plasma adrenal and thyroid hormone levels and immune responses in chicken lines divergently selected for antibody responses. Poult. Sci. 2004, 83, 1644–1649. [Google Scholar] [CrossRef]
- Schweiger, M.; Schreiber, R.; Haemmerle, G.; Lass, A.; Fledelius, C.; Jacobsen, P.; Tornqvist, R.; Zechner, R.; Zimmermann, R. Adipose triglyceride lipase and hormone-sensitive lipase are the major enzymes in adipose tissue triacylglycerol catabolism. J. Biol. Chem. 2006, 281, 40236–40241. [Google Scholar] [CrossRef]
- Blahová, J.; Dobšíková, R.; Straková, E.; Suchý, P. Effect of low environmental temperature on performance and blood system in broiler chickens (Gallus domesticus). Acta. Vet. Brno. 2007, 76, 17–23. [Google Scholar] [CrossRef]
- Şahin, K.; Önderci, M. Optimal dietary concentrations of vitamin C and chromium for alleviating the effect of low ambient temperature on serum insulin, corticosterone, and some blood metabolites in laying hens. J. Trace. Elem. Exp. Med. 2002, 15, 153–161. [Google Scholar] [CrossRef]
- Siegel, H.S. Stress, strains and resistance. Br. Poult. Sci. 1995, 36, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Sahin, N.; Kucuk, O. Effects of chromium, and ascorbic acid supplementation on growth, carcass traits, serum metabolites, and antioxidant status of broiler chickens reared at a high ambient temperature (32 °C). Nutr. Res. 2003, 23, 225–238. [Google Scholar] [CrossRef]
- Şahin, K.; Küçuk, O.; Şahin, N. Effects of dietary chromium picolinate supplementation on performance and plasma concentrations of insulin and corticosterone in laying hens under low ambient temperature. J. Anim. Physiol. Anim. Nutr. 2001, 85, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Singh, S.N.; Saha, M.; Kain, T.C.; Tyagi, A.K.; Ray, U.S. Antioxidant and oxidative stress responses of sojourners at high altitude in different climatic temperatures. Int. J. Biometeorol. 2010, 54, 85–92. [Google Scholar] [CrossRef]
- Rahmani, M.; Golian, A.; Kermanshahi, H.; Bassami, M.R. Effects of curcumin or nanocurcumin on blood biochemical parameters, intestinal morphology and microbial population of broiler chickens reared under normal and cold stress conditions. J. Appl. Anim. Res. 2018, 46, 200–209. [Google Scholar] [CrossRef]
- Fathi, M.; Nazer, A.K.; Nezhad, Y.E.; Shahryar, H.A.; Daneshyar, M.; Tanha, T. The role of oxidative stress in development of congestive heart failure (CHF) in broiler with pulmonary hypertension syndrome (PHS). J. Anim. Vet. Adv. 2011, 10, 2724–2729. [Google Scholar]
- Cangar, Ö.; Aerts, J.M.; Buyse, J.; Berckmans, D. Quantification of the spatial distribution of surface temperatures of broilers. Poult. Sci. 2008, 87, 2493–2499. [Google Scholar] [CrossRef]
- Kim, D.H.; Lee, Y.K.; Lee, S.D.; Kim, S.H.; Lee, K.W. Physiological and behavioral responses of laying hens exposed to long-term high temperature. J. Therm. Biol. 2021, 99, 103017. [Google Scholar] [CrossRef]
- Kozlowski, C.P.; Bauman, J.E.; Caldwell Hahn, D. A simplified method for extracting androgens from avian egg yolks. Zoo Biol. 2009, 28, 137–143. [Google Scholar] [CrossRef]
- Dhanalakshmi, S.; Devi, R.S.; Srikumar, R.; Manikandan, S.; Thangaraj, R. Protective effect of triphala on cold stress-induced behavioral and biochemical abnormalities in rats. Yakugaku Zasshi 2007, 127, 1863–1867. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.L.; Chen, Y.; Xue, J.J.; Huang, X.F.; Chen, Z.P.; Wang, Q.G.; Wang, C. Effects of ambient temperature on the growth performance, fat deposition, and intestinal morphology of geese from 28 to 49 days of age. Poult. Sci. 2022, 101, 101814. [Google Scholar] [CrossRef]
- Zurak, D.; Slovenec, P.; Janječić, Z.; Bedeković, X.D.; Pintar, J.; Kljak, K. Overview on recent findings of nutritional and non-nutritional factors affecting egg yolk pigmentation. Worlds Poult. Sci. J. 2022, 78, 531–560. [Google Scholar] [CrossRef]
- Ipek, A.Y.D.I.N.; Sahan, U. Effects of cold stress on broiler performance and ascites susceptibility. Asian-Australas. J. Anim. Sci. 2006, 19, 734–738. [Google Scholar] [CrossRef]
- Sahin, N.; Sahin, K.; Onderci, M.; Ozcelik, M.; Smith, M.O. In vivo antioxidant properties of vitamin E and chromium in cold-stressed Japanese quails. Arch. Anim. Nutr. 2003, 57, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Yahav, S. Limitations in energy intake affect the ability of young turkeys to cope with low ambient temperatures. J. Therm. Biol. 2002, 27, 103–108. [Google Scholar] [CrossRef]
- Yahav, S. The effect of constant and diurnal cyclic temperatures on performance and blood system of young turkeys. J. Therm. Biol. 1999, 24, 71–78. [Google Scholar] [CrossRef]
- Kim, N.Y.; Kim, S.J.; Oh, M.; Jang, S.Y.; Moon, S.H. Changes in facial surface temperature of laying hens under different thermal conditions. Anim. Biosci. 2021, 34, 1235–1242. [Google Scholar] [CrossRef]
- Nascimento, S.T.; da Silva, I.J.O.; Maia, A.S.C.; de Castro, A.C.; Vieira, F.M.C. Mean surface temperature prediction models for broiler chickens—A study of sensible heat flow. Int. J. Biometeorol. 2014, 58, 195–201. [Google Scholar] [CrossRef]
- Andrade, R.R.; Tinôco, I.F.F.; Baêta, F.C.; Barbari, M.; Conti, L.; Cecon, P.R.; Cândido, M.G.L.; Martins, I.T.A.; Junior Teles, C.G.S. Evaluation of the surface temperature of laying hens in different thermal environments during the initial stage of age based on thermographic images. Agron. Res. 2017, 15, 629–638. [Google Scholar]
- Al-Saffar, A.A.; Rose, S.P. Ambient temperature and the egg laying characteristics of laying fowl. Worlds Poult. Sci. J. 2002, 58, 317–331. [Google Scholar] [CrossRef]
- Samiullah, S.; Roberts, J.R.; Chousalkar, K. Eggshell color in brown-egg laying hens—A review. Poult. Sci. 2015, 94, 2566–2575. [Google Scholar] [CrossRef]
- Guo, J.L.; Zheng, Q.H.; Yin, Q.Q.; Cheng, W.; Jiang, Y.B. Study on mechanism of ascites syndrome of broilers. Am. J. Anim. Vet. Sci. 2007, 2, 62–65. [Google Scholar] [CrossRef]
- Quinteiro-Filho, W.M.; Rodrigues, M.V.; Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.L.; Sá, L.R.M.; Ferreira, A.J.P.; Palermo-Neto, J. Acute heat stress impairs performance parameters and induces mild intestinal enteritis in broiler chickens: Role of acute hypothalamic-pituitary-adrenal axis activation. J. Anim. Sci. 2012, 90, 1986–1994. [Google Scholar] [CrossRef] [PubMed]
- Downing, J.A.; Bryden, W.L. Determination of corticosterone concentrations in egg albumen: A non-invasive indicator of stress in laying hens. Physiol. Behav. 2008, 95, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Alm, M.; Tauson, R.; Holm, L.; Wichman, A.; Kalliokoski, O.; Wall, H. Welfare indicators in laying hens in relation to nest exclusion. Poult. Sci. 2016, 95, 1238–1247. [Google Scholar] [CrossRef] [PubMed]
- Caulfield, M.P.; Padula, M.P. HPLC MS-MS analysis shows measurement of corticosterone in egg albumen is not a valid indicator of chicken welfare. Animals 2020, 10, 821. [Google Scholar] [CrossRef]
- Borsoi, A.; Quinteiro-Filho, W.M.; Calefi, A.S.; Piantino Ferreira, A.J.; Astolfi-Ferreira, C.S.; Florio, J.C.; Palermo-Neto, J. Effects of cold stress and Salmonella Heidelberg infection on bacterial load and immunity of chickens. Avian Pathol. 2015, 44, 490–497. [Google Scholar] [CrossRef] [PubMed]
- El-halawani, M.E.; Waibel, P.E.; Appel, J.R.; Good, A.L. Effects of temperature stress on catecholamines and corticosterone of male turkeys. Am. J. Physiol. 1973, 224, 384–388. [Google Scholar] [CrossRef]
- Buckland, R.B.; Blagrave, K.; Lague, P.C. Competitive protein-binding assay for corticoids in the peripheral plasma of the immature chicken. Poult. Sci. 1974, 53, 241–245. [Google Scholar] [CrossRef]
- Hester, P.Y.; Muir, W.M.; Craig, J.V.; Albright, J.L. Group selection for adaptation to multiple-hen cages: Hematology and adrenal function. Poult. Sci. 1996, 75, 1295–1307. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialogues Clin. Neurosci. 2006, 8, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.Y.; Cheng, H.W. Warm perches: A novel approach for reducing cold stress effect on production, plasma hormones, and immunity in laying hens. Poult. Sci. 2021, 100, 101294. [Google Scholar] [CrossRef]
- Vandenborne, K.; de Groef, B.; Geelissen, S.M.E.; Kühn, E.R.; Darras, V.M.; van der Geyten, S. Corticosterone-induced negative feedback mechanisms within the hypothalamo-pituitary-adrenal axis of the chicken. J. Endocr. 2005, 185, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Angelier, F.; Wingfield, J.C. Importance of the glucocorticoid stress response in a changing world: Theory, hypotheses and perspectives. Gen. Comp. Endocrinol. 2012, 190, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Houshmand, M.; Azhar, K.; Zulkifli, I.; Bejo, M.H.; Kamyab, A. Effects of prebiotic, protein level, and stocking density on performance, immunity, and stress indicators of broilers. Poult. Sci. 2012, 91, 393–401. [Google Scholar] [CrossRef]
- Zhou, H.J.; Kong, L.L.; Zhu, L.X.; Hu, X.Y.; Busye, J.; Song, Z.G. Effects of cold stress on growth performance, serum biochemistry, intestinal barrier molecules, and adenosine monophosphate-activated protein kinase in broilers. Animal 2021, 15, 100138. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Bi, M.Y.; Yao, H.D.; Fu, J.; Li, S.; Xu, S.W. Effect of cold stress on expression of AMPKα-PPARα pathway and inflammation genes. Avian Dis. 2014, 58, 415–426. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).