
Alterations in Gene Expression and the Fatty Acid Profile Impact but Do Not Compromise the In Vitro Maturation of Zebrafish (Danio rerio) Stage III Ovarian Follicles after Cryopreservation
 , and
, and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Treatment and Ovarian Follicle Collection
2.2. Vitrification of Ovarian Follicles
2.3. Total RNA Extraction and Quantitative RT-PCR
2.4. Fatty Acid Profile
2.5. In Vitro Culture of Fresh and Vitrified Stage III Ovarian Follicles
2.6. Ovarian Follicle Viability Assay
2.7. Statistical Analysis
3. Results
3.1. Reproductive Pathway Genes
3.2. Fatty Acid Profile
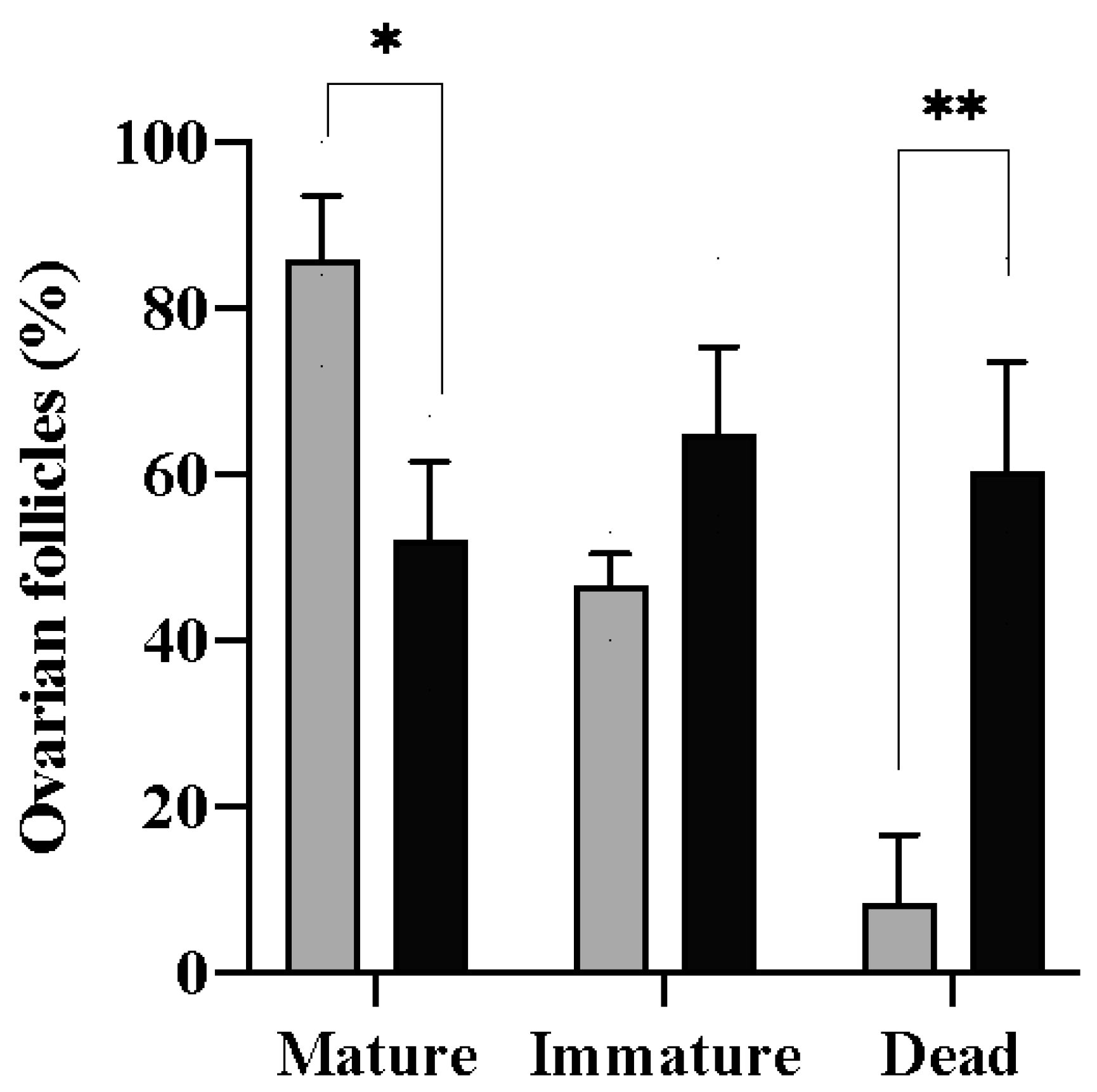
3.3. In Vitro Maturation of Fresh and Vitrified Follicles
4. Discussion
4.1. Impact of Follicle Vitrification on the Expression of Reproductive Pathway Genes
4.2. Impact of Vitrification on the Fatty Acid (FA) Profile
4.3. Impact of Vitrification on In Vitro Development Capacity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Labbe, C.; France, V.; Robles, M.; Herraez, P. Cryopreservation of gametes for aquaculture and alternative cell sources for genome preservation. In Advances in Aquaculture Hatchery Technology; Woodhead Publishing: Sawston, UK, 2012; ISBN 978-0-85709-119-2. [Google Scholar]
- Anil, S.; Zampolla, T.; Zhang, T. Development of in vitro culture method for zebrafish ovarian tissue fragment. Cryobiology 2011, 63, 311–312. [Google Scholar] [CrossRef]
- Asturiano, J.F.; Cabrita, E.; Horváth, Á. Progress, Challenges and perspectives on fish gamete cryopreservation: A mini-review. Gen. Comp. Endocrinol. 2017, 245, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Marques, L.S.; Fossati, A.A.N.; Rodrigues, R.B.; Da Rosa, H.T.; Izaguirry, A.P.; Ramalho, J.B.; Moreira, J.C.F.; Santos, F.W.; Zhang, T.; Streit, D.P. Slow freezing versus vitrification for the cryopreservation of zebrafish (Danio rerio) ovarian tissue. Sci. Rep. 2019, 9, 15353. [Google Scholar] [CrossRef]
- Martínez-Páramo, S.; Horváth, Á.; Labbé, C.; Zhang, T.; Robles, V.; Herráez, P.; Suquet, M.; Adams, S.; Viveiros, A.; Tiersch, T.R.; et al. Cryobanking of aquatic species. Aquaculture 2017, 472, 156–177. [Google Scholar] [CrossRef] [PubMed]
- Lujić, J.; Franěk, R.; Marinović, Z.; Kašpar, V.; Xie, X.; Horváth, Á.; Pšenička, M.; Urbányi, B. Vitrification of the ovarian tissue in sturgeons. Theriogenology 2023, 196, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Sturmey, R.; Reis, A.; Leese, H.; McEvoy, T. Role of fatty acids in energy provision during oocyte maturation and early embryo development. Reprod. Domest. Anim. 2009, 44, 50–58. [Google Scholar] [CrossRef]
- Zhou, C.-J.; Wang, D.-H.; Niu, X.-X.; Kong, X.-W.; Li, Y.-J.; Ren, J.; Zhou, H.-X.; Lu, A.; Zhao, Y.-F.; Liang, C.-G. High survival of mouse oocytes using an optimized vitrification protocol. Sci. Rep. 2016, 6, 19465. [Google Scholar] [CrossRef] [PubMed]
- Momozawa, K.; Matsuzawa, A.; Tokunaga, Y.; Abe, S.; Koyanagi, Y.; Kurita, M.; Nakano, M.; Miyake, T. Efficient vitrification of mouse embryos using the kitasato vitrification system as a novel vitrification device. Reprod. Biol. Endocrinol. 2017, 15, 29. [Google Scholar] [CrossRef]
- Gibbons, A.; Bruno-Galarraga, M.; Fernandez, J.; Gonzalez-Bulnes, A.; Cueto, M.; Gibbons, A.; Bruno-Galarraga, M.; Fernandez, J.; Gonzalez-Bulnes, A.; Cueto, M. Vitrified embryo transfer in Merino sheep under extensive conditions. Anim. Reprod. 2019, 16, 297–301. [Google Scholar] [CrossRef]
- Ledda, S.; Kelly, J.M.; Nieddu, S.; Bebbere, D.; Ariu, F.; Bogliolo, L.; Natan, D.; Arav, A. High in vitro survival rate of sheep in vitro produced blastocysts vitrified with a new method and device. J. Anim. Sci. Biotechnol. 2019, 10, 90. [Google Scholar] [CrossRef]
- Elnahas, A.; Alcolak, E.; Marar, E.A.; Elnahas, T.; Elnahas, K.; Palapelas, V.; Diedrich, K.; Al-Hasani, S. Vitrification of human oocytes and different development stages of embryos: An overview. Middle East Fertil. Soc. J. 2010, 15, 2–9. [Google Scholar] [CrossRef]
- Gallardo, M.; Saenz, J.; Risco, R. Human oocytes and zygotes are ready for ultra-fast vitrification after 2 minutes of exposure to standard CPA solutions. Sci. Rep. 2019, 9, 15986. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.; Rawson, D.M.; Zhang, T. Development of in vitro culture method for early stage zebrafish (Danio rerio) ovarian follicles for use in cryopreservation studies. Theriogenology 2010, 74, 290–303. [Google Scholar] [CrossRef] [PubMed]
- Godoy, L.; Streit Jr, D.; Zampolla, T.; Bos-Mikich, A.; Zhang, T. A study on the vitrification of stage III zebrafish (Danio rerio) ovarian follicles. Cryobiology 2013, 67, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Streit Júnior, D.P.; de Godoy, L.C.; Ribeiro, R.P.; Fornari, D.C.; Digmayer, M.; Zhang, T. Cryopreservation of Embryos and oocytes of South American Fish Species; InTech: Singapore, 2014. [Google Scholar]
- Marques, L.S.; Bos-Mikich, A.; Godoy, L.C.; Silva, L.A.; Maschio, D.; Zhang, T.; Streit, D.P. Viability of zebrafish (Danio rerio) ovarian follicles after vitrification in a metal container. Cryobiology 2015, 71, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Biran, J.; Sivan, B. Endocrine Control of Reproduction, Fish. In Encyclopedia of Reproduction, 2nd ed.; Skinner, M.K., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 6, pp. 362–368. [Google Scholar]
- Vissio, P.G.; Di Yorio, M.P.; Pérez-Sirkin, D.I.; Somoza, G.M.; Tsutsui, K.; Sallemi, J.E. Developmental aspects of the hypothalamic-pituitary network related to reproduction in teleost fish. Front. Neuroendocrinol. 2021, 63, 100948. [Google Scholar] [CrossRef]
- Li, L.; Shi, X.; Shi, Y.; Wang, Z. The signaling pathways involved in ovarian follicle development. Front. Physiol. 2021, 12, 730196. [Google Scholar] [CrossRef] [PubMed]
- Lubzens, E.; Young, G.; Bobe, J.; Cerdà, J. Oogenesis in teleosts: How eggs are formed. Gen. Comp. Endocrinol. 2010, 165, 367–389. [Google Scholar] [CrossRef]
- Mylonas, C.; Zohar, Y.; Pankhurst, N.; Kagawa, H. Reproduction and broodstock management. In Sparidae; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 95–131. ISBN 978-1-4443-9221-0. [Google Scholar]
- Zhang, Z.; Zhu, B.; Ge, W. Genetic analysis of zebrafish gonadotropin (fsh and lh) functions by TALEN-mediated gene disruption. Mol. Endocrinol. 2015, 29, 76–98. [Google Scholar] [CrossRef]
- Tocher, D.R. Metabolism and functions of lipids and fatty acids in teleost fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Mazorra, C.; Bruce, M.; Bell, J.G.; Davie, A.; Alorend, E.; Jordan, N.; Rees, J.; Papanikos, N.; Porter, M.; Bromage, N. Dietary lipid enhancement of broodstock reproductive performance and egg and larval quality in Atlantic halibut (Hippoglossus hippoglossus). Aquaculture 2003, 227, 21–33. [Google Scholar] [CrossRef]
- Bowden, L.A.; Restall, C.J.; Rowley, A.F. The Influence of environmental temperature on membrane fluidity, fatty acid composition and lipoxygenase product generation in head kidney leucocytes of the rainbow trout, Oncorhynchus mykiss. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1996, 115, 375–382. [Google Scholar] [CrossRef]
- Anido, R.V.; Zaniboni-Filho, E.; Garcia, A.S.; Baggio, S.R.; Fracalossi, D.M. Characterization of the ovary fatty acids composition of Rhamdia quelen (Quoy & Gaimard) (Teleostei: Siluriformes), throughout their reproductive cycle. Neotrop. Ichthyol. 2015, 13, 453–460. [Google Scholar] [CrossRef]
- Selman, K.; Wallace, R.A.; Sarka, A.; Qi, X. Stages of oocyte development in the zebrafish, Brachy Danio rerio. J. Morphol. 1993, 218, 203–224. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. Development of a gas chromatographic method for profiling neutral lipids in marine samples. Master’s Thesis, Memorial University of Newfoundland, St. John’s, Canada, 1995. [Google Scholar]
- Christie, W.W.; Han, X. Lipid Analysis, 4th ed.; Oily Press Lipid Library Series; Woodhead Publishing: Sawston, UK, 2012; p. 5. ISBN 978-0-9552512-4-5. [Google Scholar]
- Seki, S.; Kouya, T.; Tsuchiya, R.; Valdez, D.M.; Jin, B.; Hara, T.; Saida, N.; Kasai, M.; Edashige, K. Development of a reliable in vitro maturation system for zebrafish oocytes. Reproduction 2008, 135, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Horiguchi, R.; Miura, S.; Nakamura, M. Sex- and tissue-specific expression of P450 aromatase (cyp19a1a) in the yellowtail clownfish, Amphiprion clarkii. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 155, 237–244. [Google Scholar] [CrossRef]
- Ogiwara, K.; Fujimori, C.; Rajapakse, S.; Takahashi, T. Characterization of luteinizing hormone and luteinizing hormone Receptor and their indispensable role in the ovulatory process of the medaka. PLoS ONE 2013, 8, e54482. [Google Scholar] [CrossRef] [PubMed]
- Riccetti, L.; Yvinec, R.; Klett, D.; Gallay, N.; Combarnous, Y.; Reiter, E.; Simoni, M.; Casarini, L.; Ayoub, M.A. Human luteinizing hormone and chorionic gonadotropin display biased agonism at the LH and LH/CG receptors. Sci. Rep. 2017, 7, 940. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, Y.; Yamashita, M. Regulation of oocyte maturation in fish. Dev. Growth Differ. 2008, 50, S195–S219. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Ogiwara, K. cAMP signaling in ovarian physiology in teleosts: A review. Cell. Signal. 2023, 101, 110499. [Google Scholar] [CrossRef]
- Fitzpatrick, S.L.; Carlone, D.L.; Robker, R.L.; Richards, J.S. Expression of aromatase in the ovary: Down-regulation of mRNA by the ovulatory luteinizing hormone surge. Steroids 1997, 62, 197–206. [Google Scholar] [CrossRef]
- Watanabe, M.; Tanaka, M.; Kobayashi, D.; Yoshiura, Y.; Oba, Y.; Nagahama, Y. Medaka (Oryzias latipes) FTZ-F1 potentially regulates the transcription of P-450 aromatase in ovarian follicles: cDNA cloning and functional characterization. Mol. Cell. Endocrinol. 1999, 149, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Yoshiura, Y.; Senthilkumaran, B.; Watanabe, M.; Oba, Y.; Kobayashi, T.; Nagahama, Y. Synergistic expression of Ad4BP/SF-1 and cytochrome P-450 aromatase (ovarian type) in the ovary of Nile tilapia, Oreochromis niloticus, during vitellogenesis suggests transcriptional interaction. Biol. Reprod. 2003, 68, 1545–1553. [Google Scholar] [CrossRef]
- Ascoli, M.; Fanelli, F.; Segaloff, D.L. The lutropin/choriogonadotropin receptor, a 2002 perspective. Endocr. Rev. 2002, 23, 141–174. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Dong, Y.; Matzuk, M.M.; Kumar, T.R. Targeted disruption of luteinizing hormone beta-subunit leads to hypogonadism, defects in gonadal steroidogenesis, and infertility. Proc. Natl. Acad. Sci. USA 2004, 101, 17294–17299. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.R.; Habibi, H.R. Estrogen receptor function and regulation in fish and other vertebrates. Gen. Comp. Endocrinol. 2013, 192, 15–24. [Google Scholar] [CrossRef]
- Li, J.; Chu, L.; Sun, X.; Liu, Y.; Cheng, C.H.K. IGFs mediate the action of LH on oocyte maturation in zebrafish. Mol. Endocrinol. 2015, 29, 373–383. [Google Scholar] [CrossRef]
- Fuentes, N.; Silveyra, P. Estrogen receptor signaling mechanisms. Adv. Protein. Chem. Struct. Biol. 2019, 116, 135–170. [Google Scholar] [CrossRef]
- Nelson, S.; Kraak, G. The role of the insulin-like growth factor (IGF) system in zebrafish (Danio rerio) ovarian development. Gen. Comp. Endocrinol. 2010, 168, 103–110. [Google Scholar] [CrossRef]
- Liu, T.; Qu, J.; Tian, M.; Yang, R.; Song, X.; Li, R.; Yan, J.; Qiao, J. Lipid metabolic Process involved in oocyte maturation during folliculogenesis. Front. Cell Dev. Biol. 2022, 10, 806890. [Google Scholar] [CrossRef] [PubMed]
- Nzioka, A.; Valencia, A.; Atxaerandio-Landa, A.; Diaz de Cerio, O.; Hossain, M.A.; Korta, M.; Ortiz-Zarragoitia, M.; Cancio, I. Apoptosis and autophagy-related gene transcription during ovarian follicular atresia in European hake (Merluccius merluccius). Mar. Environ. Res. 2023, 183, 105846. [Google Scholar] [CrossRef] [PubMed]
- Sargent, J.R.; Tocher, D.R.; Bell, J. Fish Nutrition, 3rd ed.; Elsevier Science: San Diego, CA, USA, 2002. [Google Scholar]
- da Silva Ribeiro, C.; Schreiner, M.; Iannini, C.A.N.; Dal’Olio Gomes, A.; Tolussi, C.E.; Moreira, R.G. Acute and chronic effects of temperature on membrane adjustments in the gills of a neotropical catfish. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2021, 256, 110625. [Google Scholar] [CrossRef] [PubMed]
- Zeron, Y.; Ocheretny, A.; Kedar, O.; Borochov, A.; Sklan, D.; Arav, A. Seasonal changes in bovine fertility: Relation to developmental competence of oocytes, membrane properties and fatty acid composition of follicles. Reproduction 2001, 121, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Pati, D.; Habibi, H.R. Involvement of protein kinase C and arachidonic acid pathways in the gonadotropin-releasing hormone regulation of oocyte meiosis and follicular steroidogenesis in the goldfish ovary1. Biol. Reprod. 2002, 66, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.; Heisenberg, C.-P. The yolk syncytial layer in early zebrafish development. Trends Cell Biol. 2010, 20, 586–592. [Google Scholar] [CrossRef]
- Riddle, M.R.; Hu, C.-K. Fish models for investigating nutritional regulation of embryonic development. Dev. Biol. 2021, 476, 101–111. [Google Scholar] [CrossRef]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- de Mello, F.; Marques, V.; Pires Vieira Morais de Faria, N.; Godoy, L.; Moreira, R. Vitrification changes the fatty acids profile of zebrafish ovarian follicles at different developmental stages. Cryobiology 2021, 103, 189. [Google Scholar] [CrossRef]
- Laale, H.W. The perivitelline space and egg envelopes of bony fishes: A review. Copeia 1980, 1980, 210–226. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Acc Number | Product Length (pb) |
|---|---|---|---|---|
| Luteinizing hormone/choriogonadotropin receptor (lhcgr) | AAGGACGAGTCGCTGAAAC | CACAGCAGAAAGATGTCAGC | AY424302 | 189 |
| Follicle-stimulating hormone receptor (fshr) | GATCCACTCGCTCTTTCGTC | CCGTTCTCGGACACCACTAT | AY424301 | 169 |
| Cytochrome P450 Family 19 Subfamily A Member 1 (Cyp19a1) | TGAGCACGATCTGCTTCAGA | TGCAACTCCTGAGCATCTCT | AF183906 | 129 |
| Insulin-like growth factor I (IGF-I) | CGAAGAAGGTCAAGGTGCTC | GACTGCATCCATCTCTGCAA | AH010825 | 211 |
| Steroidogenic acute regulatory protein (Star) | CCAAGTGCAGATGACCCCAA | GGAAGGTGTGTGCCCTTGTT | NM_131663.1 | 213 |
| Estrogen receptor 1 (esr1) | GGTCCAGTGTGGTGTCCTCT | AGAAAGCTTTGCATCCCTCA | NM_152959.1 | 204 |
| Estrogen receptor 2a (esr2a) | TAGTGGGACTTGGACCGAAC | TTCACACGACCACACTCCAT | NM_180966.2 | 187 |
| Estrogen receptor 2b (esr2b) | TTGTGTTCTCCAGCATGAGC | CCACATATGGGGAAGGAATG | NM_174862.3 | 156 |
| Caspase3 (casp3) | GTGCCAGTCAACAAACAAAG | CATCTCCAACCGCTTAACG | NM_131877.3 | 172 |
| Tumor protein p53 (tp53) | GGCTCTTGCTGGGACATCAT | TGGATGGCTGAGGCTGTTCT | AF365873.1 | 159 |
| Elongation factor alpha (ef1-alfa) | GGTACTACTCTTCTTGATGC | GACTTGACCTCAGTGGTTAC | AY422992 | 200 |
| Beta-actin | GCCAACACTGTATTGTCTGG | GTACTCCTGCTTGCTAATCC | BC165823 | 203 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Mello, F.; Alonso, D.J.; Faria, N.P.V.M.d.; Marques, V.H.; Oliveira, E.F.d.; de Mello, P.H.; Godoy, L.C.d.; Moreira, R.G. Alterations in Gene Expression and the Fatty Acid Profile Impact but Do Not Compromise the In Vitro Maturation of Zebrafish (Danio rerio) Stage III Ovarian Follicles after Cryopreservation. Animals 2023, 13, 3563. https://doi.org/10.3390/ani13223563
de Mello F, Alonso DJ, Faria NPVMd, Marques VH, Oliveira EFd, de Mello PH, Godoy LCd, Moreira RG. Alterations in Gene Expression and the Fatty Acid Profile Impact but Do Not Compromise the In Vitro Maturation of Zebrafish (Danio rerio) Stage III Ovarian Follicles after Cryopreservation. Animals. 2023; 13(22):3563. https://doi.org/10.3390/ani13223563
Chicago/Turabian Stylede Mello, Fernanda, Daniel Jaen Alonso, Natália Pires Vieira Morais de Faria, Victor Hugo Marques, Ethiene Fernandes de Oliveira, Paulo Henrique de Mello, Leandro César de Godoy, and Renata Guimaraes Moreira. 2023. "Alterations in Gene Expression and the Fatty Acid Profile Impact but Do Not Compromise the In Vitro Maturation of Zebrafish (Danio rerio) Stage III Ovarian Follicles after Cryopreservation" Animals 13, no. 22: 3563. https://doi.org/10.3390/ani13223563
APA Stylede Mello, F., Alonso, D. J., Faria, N. P. V. M. d., Marques, V. H., Oliveira, E. F. d., de Mello, P. H., Godoy, L. C. d., & Moreira, R. G. (2023). Alterations in Gene Expression and the Fatty Acid Profile Impact but Do Not Compromise the In Vitro Maturation of Zebrafish (Danio rerio) Stage III Ovarian Follicles after Cryopreservation. Animals, 13(22), 3563. https://doi.org/10.3390/ani13223563