
A Description of Echinochasmus pseudobeleocephalus n. sp. (Echinochasmidae) Based on Morphological and Molecular Data
 , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Life Cycle and Morphology of Adult Individuals
2.2. DNA Extraction, Amplification, and Sequencing
3. Results
4. Remark
4.1. Morphological Identification
4.2. Molecular Identification
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skrjabin, K.I.; Bashkirova, E.Y. Family Echinostomatidae Diestz, 1909. Trematodes Animals and Humans. Fundamentals of Trematodology; Publishing House of the Academy of Sciences of the USSR: Moscow, Russia, 1956; Volume 21, pp. 51–915. (In Russian) [Google Scholar]
- Kostadinova, A.; Jones, A.; Bray, R.A.; Gibson, D.I. Family Psilostomidae Looss, 1900. Keys to the Trematoda; Jones, A., Bray, R.A., Gibson, D.I., Eds.; CABI Publishing and the Natural History Museum: Sofia, Bulgaria, 2005; Volume 2, pp. 99–118. [Google Scholar]
- Filimonova, L.V. On the phenomenon of amphixia in trematodes Echinochasmus japonicus (Echinostomatidae). J. Zool. 1979, 58, 1103–1110. (In Russian) [Google Scholar]
- Besprozvannykh, V.V. Life cycles of trematodes Echinochasmus japonicus and E. beleocephalus (Echinostomatidae) in conditions of Primorsky Region. Parazitologia 2009, 43, 248–258. (In Russian) [Google Scholar]
- Tkach, V.V.; Kudlai, O.; Kostadinova, A. Molecular phylogeny and systematics of the Echinostomatoidea Looss, 1899 (Platyhelminthes: Digenea). Int. J. Parasitol. 2016, 46, 171–185. [Google Scholar] [CrossRef]
- Besprozvannykh, V.V.; Ermolenko, A.V. Stephanoprora chasanensis n. sp. (Digenea: Echinochasmidae): Morphology, life cycle, and molecular data. Int. J. Parasitol. 2017, 66, 863–870. [Google Scholar] [CrossRef]
- Tatonova, Y.V.; Izrailskaia, A.V.; Besprozvannykh, V.V. Stephanoprora amurensis sp. nov., Echinochasmus milvi Yamaguti, 1939 and E. suifunensis Besprozvannykh, 1991 from the Russian southern Far East and their phylogenetic relationships within the Echinochasmidae Odhner 1910. Parasitology 2020, 147, 1469–1479. [Google Scholar] [CrossRef]
- Truett, G.E.; Heege, P.; Mynatt, R.L.; Truett, A.A.; Walker, J.A.; Warman, M.L. Preparation of PCR–quality mouse genomic DNA with hot sodium hydroxide and tris (HotSHOT). Biotechniques 2000, 29, 52–54. [Google Scholar] [CrossRef]
- Tkach, V.V.; Timothy, D.; Littlewood, J.; Olson, P.D.; Kinsella, J.M.; Swiderski, Z. Molecular phylogenetic analysis of the Microphalloidea Ward, 1901 (Trematoda: Digenea). Syst. Parasitol. 2003, 56, 1–15. [Google Scholar] [CrossRef]
- Lockyer, A.E.; Olson, P.D.; Ostergaard, P.; Rollinson, D. The phylogeny of the Schistosomatidae based on three genes with emphasis on the interrelationships of Schistosoma Weinland, 1858. Parasitology 2003, 126, 203–224. [Google Scholar] [CrossRef]
- Sawabe, T.; Kita–Tsukamoto, K.; Thompson, F.L. Inferring the evolutionary history of vibrios by means of Multilocus Sequence Analysis. J. Bacteriol. 2007, 21, 7932–7936. [Google Scholar] [CrossRef]
- Krieger, J.; Fuerst, P.A. Evidence of multiple alleles of the nuclear 18S ribosomal RNA gene in sturgeon (Family: Acipenseridae). J. Appl. Ichthyol. 2002, 18, 290–297. [Google Scholar] [CrossRef]
- Bowles, J.; Blair, D.; McManus, D.P. A molecular phylogeny of the human schistosomes. Mol. Phylogenet. Evol. 1995, 4, 103–109. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. J. Mol. Struct. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Besprozvannykh, V.V.; Ermolenko, A.V.; Ngo, H.D.; Ha, N.V.; Hung, N.M.; Rozhkovan, K.V. Descriptions of digenean parasites from three snail species, Bithynia fuchsiana (Morelet), Parafossarulus striatulus Benson and Melanoides tuberculata Müller, in North Vietnam. Helmintologia 2013, 50, 190–204. [Google Scholar] [CrossRef]
- Schwelm, J.; Kudlai, O.; Smit, N.J.; Selbach, C.; Sures, B. High parasite diversity in a neglected host: Larval trematodes of Bithynia tentaculata in Central Europe. J. Helminthol. 2020, 94, e120. [Google Scholar] [CrossRef]
- Staneviciute, S.; Virmantas, S.; Romualda, P. Phylogenetic relationships of some species of the family Echinostomatidae Odner, 1910 (Trematoda), inferred from nuclear rDNA sequences and karyological analysis. Comp. Cytogenet. 2015, 9, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Tkach, V.V.; Pawlowski, J.; Mariaux, J. Phylogenetic analysis of the suborder Plagiorchiata (Platyhelminthes, Digenea) based on partial lsrDNA sequences. Int. J. Parasitol. 2000, 30, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Tkach, V.V.; Mariaux, J.; Swiderski, Z. Molecular phylogeny of the suborder Plagiorchiata and its position in the system of Digenea. In Interrelationships of the Platyhelminthes. In Interrelationships of the Platyhelminthes; CRC Press: Boca Raton, FL, USA, 2001; Volume 17, pp. 186–193. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Yamaguti, S. A Synoptical Review of Life Histories of Digenetic Trematodes of Vertebrates: With Special Reference to the Morphology of Their Larval Forms; Kaigaku Pub Co.: Tokyo, Japan, 1975; pp. 1–590. [Google Scholar]
- Bashkirova, V.Y. Echinostomatids of birds of the USSR and a review of the cycles of their development. Bashkir NIVS 1941, 3, 243–300. (In Russian) [Google Scholar]
- Yamaguti, S. Studies on the helminth fauna of Japan. Part 25. Trematodes of birds IV. Biology 1939, 8, 130–210. [Google Scholar]
- Ito, J. A monograph of cercaria in Japan and adjacent territories. Prog. Med. Parasitol. Jpn. 1964, 1, 395–550. [Google Scholar]
- Karmanova, E.M. Knowledge of the life cycle of the trematodes Echinochasmus coaxatus Dietz, 1909 and E. beleocephalus (Linstow, 1873) (Echinostomatidae). Tr. GELAN 1974, 24, 46–53. (In Russian) [Google Scholar]
- Oshmarin, P.G.; Dotsenko, T.K. About Epizootology of Helminthosises among DOMESTIC Birds in Lowlying Land Near Khanka Lake; Soobchenia DVF AN SSSR: Moscow, Russian, 1951; Volume 3, pp. 8–11. (In Russian) [Google Scholar]
- Braun, M. The so–called “free–swimming sporocyst”. Centralbl. Bakt. Parasit. 1891, 10, 215–219. (In Germany) [Google Scholar]
- Dietz, E. Die Echinostomiden der Vögel. Zoologische Jahrbücher 1910, 12, 265–512. [Google Scholar]
- Odhner, T. Nordostafrikanische Trematoden, grösstenteils vom Weissen Nile. I. Fascioliden. Results of the Swedish Zoological Expedition to Egypt and the White Nile, 1901, under the Direction of LA Jägerskiöld, No. 23A (1910) 1-170. Die Homologien der weiblichen Genitalwege beiden Trematoden und Cestoden. Zool. Anz. 1912, 23, 1–170. [Google Scholar]
- Tanabe, H. A new species Echinochasmus, E. japonicus n. sp. Nippon Byor. Gakk. Shi. 1926, 16, 295–296. [Google Scholar]
- Beaver, P.C. Studies on the life history of Echinochasmus donaldsoni n. sp., a trematode (Echinochasmidae from the Pied—Billed grebe. J. Parasitol. 1941, 27, 347–355. [Google Scholar] [CrossRef]
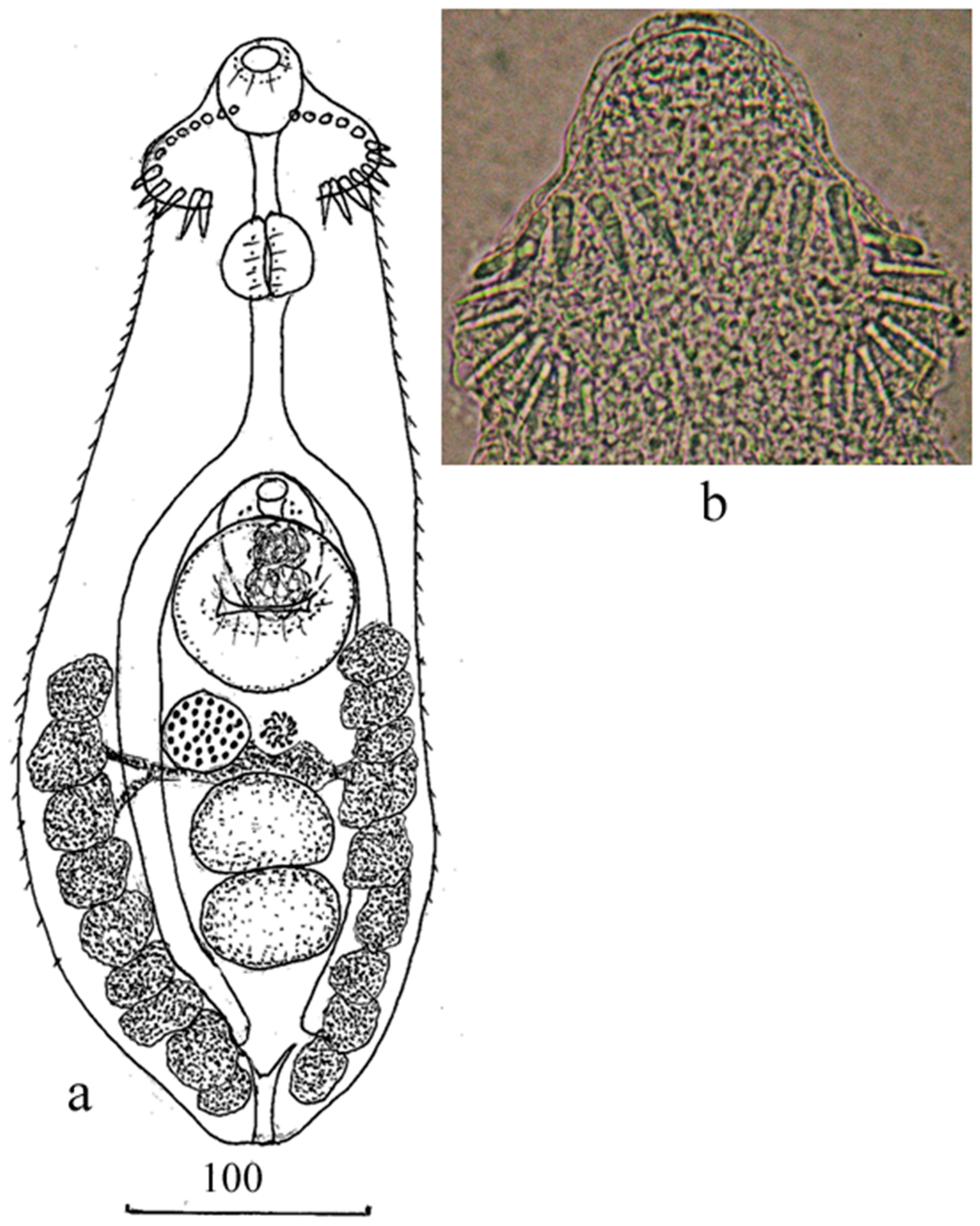


{kind=link}
{kind=link}
| Species | Developmental Stage | Accession Numbers (GenBank) | Reference | Country | Sequence Length, bp |
|---|---|---|---|---|---|
| family Echinochasmidae | |||||
| Echinochasmus pseudobeleocephalus n. sp. | Adult | OR076694, OR076695 | this study | Russia | 701 |
| Cercaria | OR076696 | ||||
| Echinochasmus japonicus | Adult | JQ890579, JQ890580 | [15] | Russia | 1370 |
| Echinochasmus beleocephalus | Adult | KT956929 | [5] | Ukraine | 1178 |
| Echinochasmus sp. 2 | Cercaria | MN726948 | [16] | Germany | 1199 |
| Echinochasmus coaxatus | Adult | KJ542643 | [17] | Ukraine; | 969 |
| Adult | KT956928 | [5] | Ukraine | 1200 | |
| Cercaria | MN726944 | [16] | Germany | 1200 | |
| Echinochasmus bursicola | Adult | KT956938 | [5] | Ukraine | 1173 |
| Microparyphium facetum | Adult | KT956933 | [5] | USA | 1291 |
| Echinochasmus sp. VS-2012 | Cercaria | JQ088098 | [17] | Lithuania | 1258 |
| Echinochasmus mordax | Adult | KT956931 | [5] | Ukraine | 1175 |
| Stephanoprora chasanensis | Adult | KT873320, KT873321 | [6] | Russia | 1369 |
| Echinochasmus sp. | Adult | KT956932 | [5] | USA | 1161 |
| Echinochasmus donaldsoni | Adult | KT956930 | [5] | USA | 1240 |
| Stephanoprora amurensis | Adult | MT447050, MT447051 | [7] | Russia | 1145 |
| Echinochasmus sp. 1 | Cercaria | MN726946, MN726947 | [16] | Germany | 1199 |
| Echinochasmus milvi | Adult | KT873317, KT873318 | [6] | Russia Russia | 1369 |
| MT447054, MT447055 | [7] | 1145 | |||
| Echinochasmus suifunensis | Adult | MT447056, MT447057 | [7] | Russia | 1145 |
| Stephanoprora sp. 1 | Adult | KT956936 | [5] | USA | 1181 |
| Stephanoprora sp. 2 | KT956937 | ||||
| Stephanoprora pseudoechinata | Adult | KT956934, KT956935 | [5] | Ukraine | 1249 |
| Outgroup | |||||
| Psilostomum brevicolle | Adult | KT956950 | [5] | Ukraine | 1292 |
| Psilochasmus oxyurus | Adult | AF151940 | [18] | Ukraine | 1239 |
| Sphaeridiotrema pseudoglobulus | Adult | KT956957 | [5] | USA | 1206 |
| Notocotylus attenuatus | Adult | AF184259 | [19] | Ukraine | 1261 |
| Characters | E. pseudobeleocephalus n. sp. | E. japonicus | E. beleocephalus | ||||||
|---|---|---|---|---|---|---|---|---|---|
| This Study | [22] | [3] | [4] | [15] | [23] | [4] | |||
| Holotype | Range (n = 7) | Mean | |||||||
| Body length (Bl) | 454 | 454–554 | 502 | 600–900 | 540–620 | 780–810 | 520–580 | 715–924 | 550–620 |
| Body width (Bw) | 173 | 139–173 | 152 | 160–180 | 150–180 | 220–250 | 162–235 | 253–330 | 130–170 |
| Bw/Bl (%) * | 38.1 | 27.2–38.1 | 30.3 | ||||||
| Forebody length (Fo) | 204 | 204–258 | 234 | ||||||
| Fo/Bl (%) ** | 44.9 | 44.4–48.6 | 46.6 | ||||||
| Oral sucker length | 39 | 31–39 | 36 | 38–42 | 40–51 | 45–50 | 23–35 | 47–51 | 34–39 |
| Oral sucker width | 35 | 35–39 | 37 | 38–42 | 40–57 | 56 | 27–42 | 47–51 | 39–42 |
| Ventral sucker length | 69 | 62–81 | 70 | 70–96 | 68–91 | 95–110 | 50–65 | 132–143 | 59–70 |
| Ventral sucker width | 73 | 65–85 | 72 | 70–96 | 74–86 | 67–89 | 54–77 | 132–154 | 62–73 |
| Ratio of suckers’ lengths | 1.78 | 1.59–2.10 | 1.94 | ||||||
| Ratio of suckers’ widths | 2.09 | 1.67–2.09 | 1.95 | ||||||
| Head-collar width | 92 | 85–116 | 96 | ||||||
| Prepharynx length | 35 | 31–50 | 40 | 30–60 | 46–68 | 28–34 | 15–42 | 33–38 | 48–50 |
| Pharynx length | 35 | 35–42 | 38 | 35–39 | 34–46 | 67–84 | 19–27 | 51–56 | 28–40 |
| Pharynx width | 39 | 31–39 | 34 | 27–32 | 34–51 | 45–50 | 23–31 | 51–56 | 34–39 |
| Oesophagus length | 58 | 58–100 | 78 | 110–210 | 97–120 | 95–130 | 92–96 | 132–198 | 67–130 |
| Ovary length | 35–39 | 31–39 | 34 | 36–48 | 40–51 | 41–60 | 35–42 | 38–43 | 34–42 |
| Ovary width | 31–35 | 27–35 | 32 | 22–30 | 46–67 | 38–49 | 35–42 | 47 | 31–50 |
| Anterior testis length | 42 | 35–62 | 48 | 60–75 | 40–68 | 83–135 | 58–80 | 43–88 | 50–70 |
| Anterior testis width | 58 | 53–77 | 61 | 54–80 | 34–46 | 100–132 | 65–92 | 34–88 | 59–80 |
| Posterior testis length | 42 | 39–62 | 47 | ||||||
| Posterior testis width | 58 | 46–62 | 59 | ||||||
| Cirrus sac length | 58 | 58–69 | 61 | 75–90 | – | 91–130 | 50–65 | 86–132 | 62–81 |
| Cirrus sac width | 46 | 39–46 | 41 | 36–48 | – | 60–74 | 42–58 | 66–77 | 34–48 |
| Post-testicular field length | 69 | 58–69 | 65 | ||||||
| Egg length | deformed | 77–90 | 63–86 | 84–89 | 80 | 73–81 | 84 | ||
| Egg width | 51–57 | 46–57 | 50 | 53 | 34–43 | 61 | |||
| Number of head-collar spines | 24 | 24 | 24 | 24 | 24 | 22 | 24 | 24 | 24 |
| No | Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cluster 1 * | 1 | Microparyphium facetum | 0.008 | 0.008 | 0.009 | 0.009 | 0.009 | 0.009 | 0.008 | 0.007 | 0.008 | 0.007 | 0.008 | 0.008 | 0.007 | 0.008 | 0.008 | 0.007 | 0.008 | |
| 2 | Echinochasmus bursicola | 0.047 | 0.004 | 0.003 | 0.003 | 0.004 | 0.004 | 0.009 | 0.009 | 0.009 | 0.009 | 0.009 | 0.009 | 0.009 | 0.009 | 0.009 | 0.008 | 0.008 | ||
| 3 | E. coaxatus | 0.059 | 0.011 | 0.004 | 0.004 | 0.005 | 0.005 | 0.010 | 0.009 | 0.009 | 0.010 | 0.010 | 0.010 | 0.010 | 0.010 | 0.010 | 0.009 | 0.009 | ||
| 4 | Echinochasmus. sp. 2 | 0.054 | 0.007 | 0.016 | 0.000 | 0.003 | 0.003 | 0.010 | 0.009 | 0.009 | 0.010 | 0.010 | 0.010 | 0.009 | 0.010 | 0.010 | 0.009 | 0.009 | ||
| 5 | E. beleocephalus | 0.054 | 0.007 | 0.016 | 0.000 | 0.003 | 0.003 | 0.010 | 0.009 | 0.009 | 0.010 | 0.010 | 0.010 | 0.009 | 0.010 | 0.010 | 0.009 | 0.009 | ||
| 6 | E. japonicus | 0.059 | 0.011 | 0.020 | 0.007 | 0.007 | 0.002 | 0.009 | 0.009 | 0.009 | 0.009 | 0.009 | 0.009 | 0.009 | 0.010 | 0.010 | 0.009 | 0.009 | ||
| 7 | E. peudobeleocephalus | 0.059 | 0.011 | 0.020 | 0.007 | 0.007 | 0.003 | 0.009 | 0.009 | 0.009 | 0.009 | 0.009 | 0.009 | 0.010 | 0.010 | 0.010 | 0.009 | 0.009 | ||
| Cluster 2 * | 8 | Echinochasmus sp. VS-2012 | 0.062 | 0.059 | 0.069 | 0.066 | 0.066 | 0.059 | 0.062 | 0.004 | 0.003 | 0.004 | 0.004 | 0.004 | 0.004 | 0.005 | 0.005 | 0.003 | 0.003 | |
| 9 | E. mordax | 0.059 | 0.056 | 0.066 | 0.063 | 0.063 | 0.056 | 0.059 | 0.011 | 0.003 | 0.004 | 0.004 | 0.003 | 0.005 | 0.005 | 0.005 | 0.004 | 0.004 | ||
| 10 | Stephanoprora chasanensis | 0.060 | 0.054 | 0.064 | 0.062 | 0.062 | 0.054 | 0.057 | 0.007 | 0.007 | 0.003 | 0.003 | 0.002 | 0.004 | 0.004 | 0.004 | 0.003 | 0.003 | ||
| 11 | Echinochasmus sp. | 0.067 | 0.062 | 0.072 | 0.069 | 0.069 | 0.062 | 0.064 | 0.011 | 0.011 | 0.007 | 0.003 | 0.003 | 0.005 | 0.005 | 0.005 | 0.004 | 0.004 | ||
| 12 | E. donaldsoni | 0.064 | 0.059 | 0.069 | 0.066 | 0.066 | 0.059 | 0.062 | 0.011 | 0.011 | 0.007 | 0.006 | 0.003 | 0.005 | 0.005 | 0.005 | 0.004 | 0.004 | ||
| 13 | S. amurensis | 0.060 | 0.057 | 0.067 | 0.064 | 0.064 | 0.057 | 0.060 | 0.010 | 0.007 | 0.003 | 0.007 | 0.007 | 0.004 | 0.004 | 0.004 | 0.003 | 0.003 | ||
| 14 | Echinochasmus sp. 1 | 0.059 | 0.056 | 0.066 | 0.063 | 0.063 | 0.059 | 0.062 | 0.014 | 0.014 | 0.010 | 0.017 | 0.017 | 0.013 | 0.005 | 0.005 | 0.004 | 0.004 | ||
| 15 | E. milvi | 0.067 | 0.062 | 0.072 | 0.063 | 0.063 | 0.062 | 0.064 | 0.014 | 0.017 | 0.010 | 0.017 | 0.017 | 0.013 | 0.017 | 0.000 | 0.004 | 0.004 | ||
| 16 | E. suifunensis | 0.067 | 0.062 | 0.072 | 0.063 | 0.063 | 0.062 | 0.064 | 0.014 | 0.017 | 0.010 | 0.017 | 0.017 | 0.013 | 0.017 | 0.000 | 0.004 | 0.004 | ||
| 17 | Stephanoprora sp. | 0.054 | 0.054 | 0.064 | 0.062 | 0.062 | 0.057 | 0.060 | 0.007 | 0.010 | 0.006 | 0.013 | 0.013 | 0.006 | 0.010 | 0.013 | 0.013 | 0.002 | ||
| 18 | S. pseudoechinata | 0.056 | 0.056 | 0.066 | 0.063 | 0.063 | 0.059 | 0.062 | 0.009 | 0.011 | 0.007 | 0.014 | 0.014 | 0.007 | 0.011 | 0.014 | 0.014 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalinina, K.A.; Besprozvannykh, V.V.; Tatonova, Y.V.; Shchelkanov, M.Y. A Description of Echinochasmus pseudobeleocephalus n. sp. (Echinochasmidae) Based on Morphological and Molecular Data. Animals 2023, 13, 3236. https://doi.org/10.3390/ani13203236
Kalinina KA, Besprozvannykh VV, Tatonova YV, Shchelkanov MY. A Description of Echinochasmus pseudobeleocephalus n. sp. (Echinochasmidae) Based on Morphological and Molecular Data. Animals. 2023; 13(20):3236. https://doi.org/10.3390/ani13203236
Chicago/Turabian StyleKalinina, Kristina Andreevna, Vladimir Vladimirovich Besprozvannykh, Yulia Viktorovna Tatonova, and Mikhail Yurievich Shchelkanov. 2023. "A Description of Echinochasmus pseudobeleocephalus n. sp. (Echinochasmidae) Based on Morphological and Molecular Data" Animals 13, no. 20: 3236. https://doi.org/10.3390/ani13203236
APA StyleKalinina, K. A., Besprozvannykh, V. V., Tatonova, Y. V., & Shchelkanov, M. Y. (2023). A Description of Echinochasmus pseudobeleocephalus n. sp. (Echinochasmidae) Based on Morphological and Molecular Data. Animals, 13(20), 3236. https://doi.org/10.3390/ani13203236