
Effects of Dietary Bacillus subtilis HC6 on Growth Performance, Antioxidant Capacity, Immunity, and Intestinal Health in Broilers

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Preparation
2.2. Birds, Experimental Design, and Diets
2.3. Sample Collection
2.4. Measured Serum Biomarkers
2.5. Serum and Liver Antioxidant Markers
2.6. Intestinal Tissue Morphology
2.7. RNA Extraction, cDNA Synthesis and Quantitative PCR
2.8. 16S Sequencing and Cecal Microbiota Analysis
2.9. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Intestinal Morphology
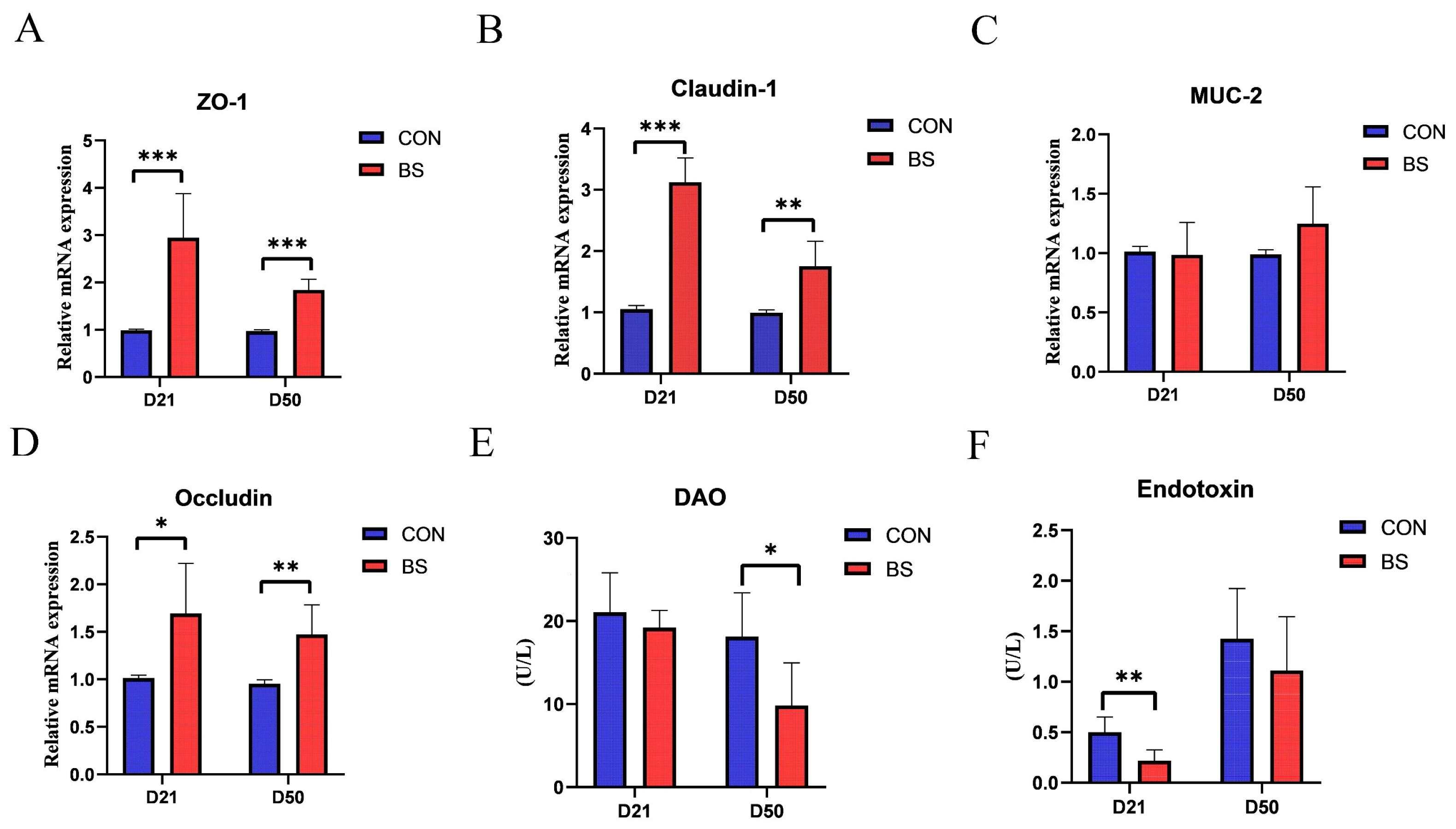
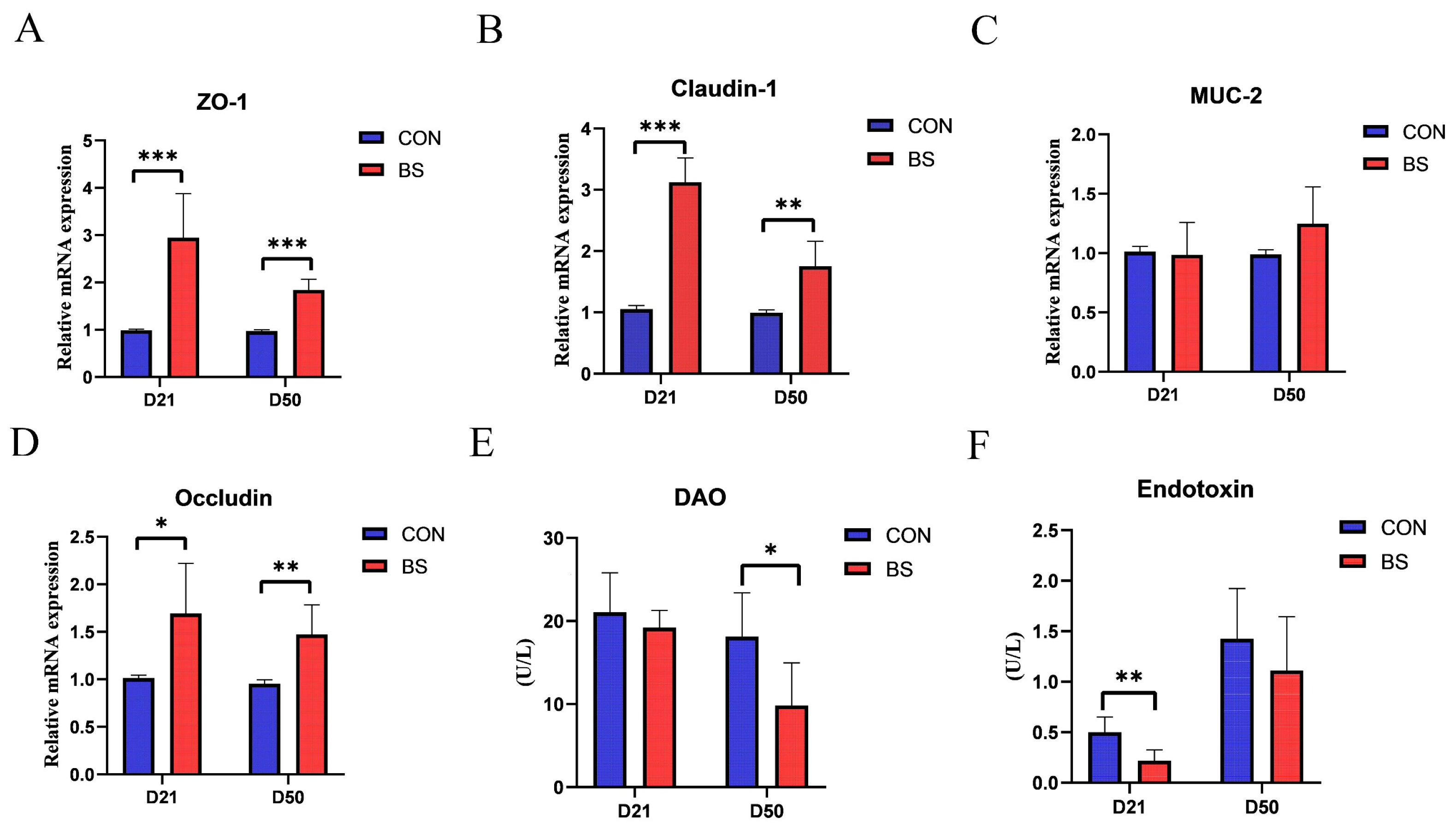
3.3. Jejunal Mucosal Barrier Function
3.4. Immune Factor
3.5. Serum and Liver Antioxidant Index
3.6. Serum Biochemical Index
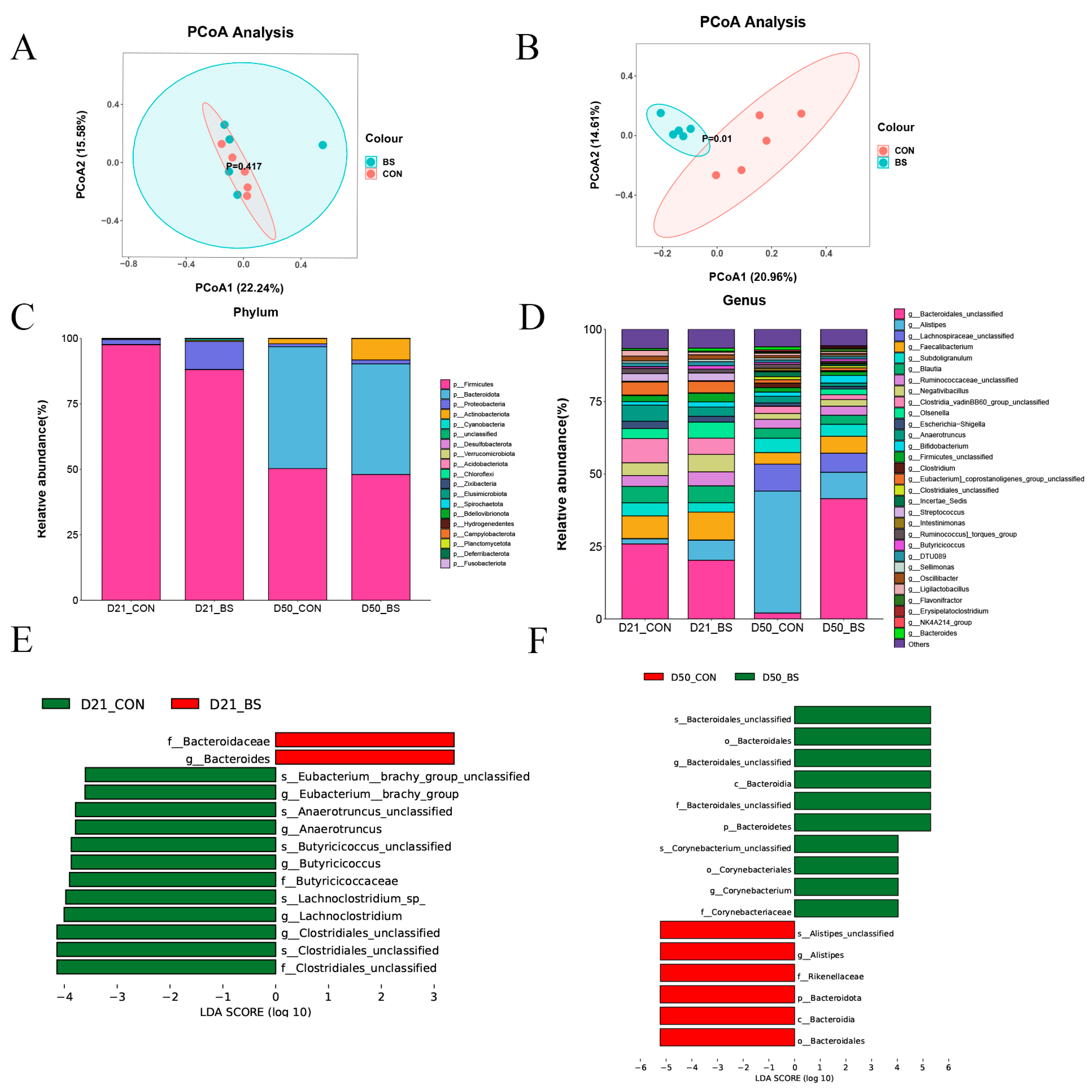
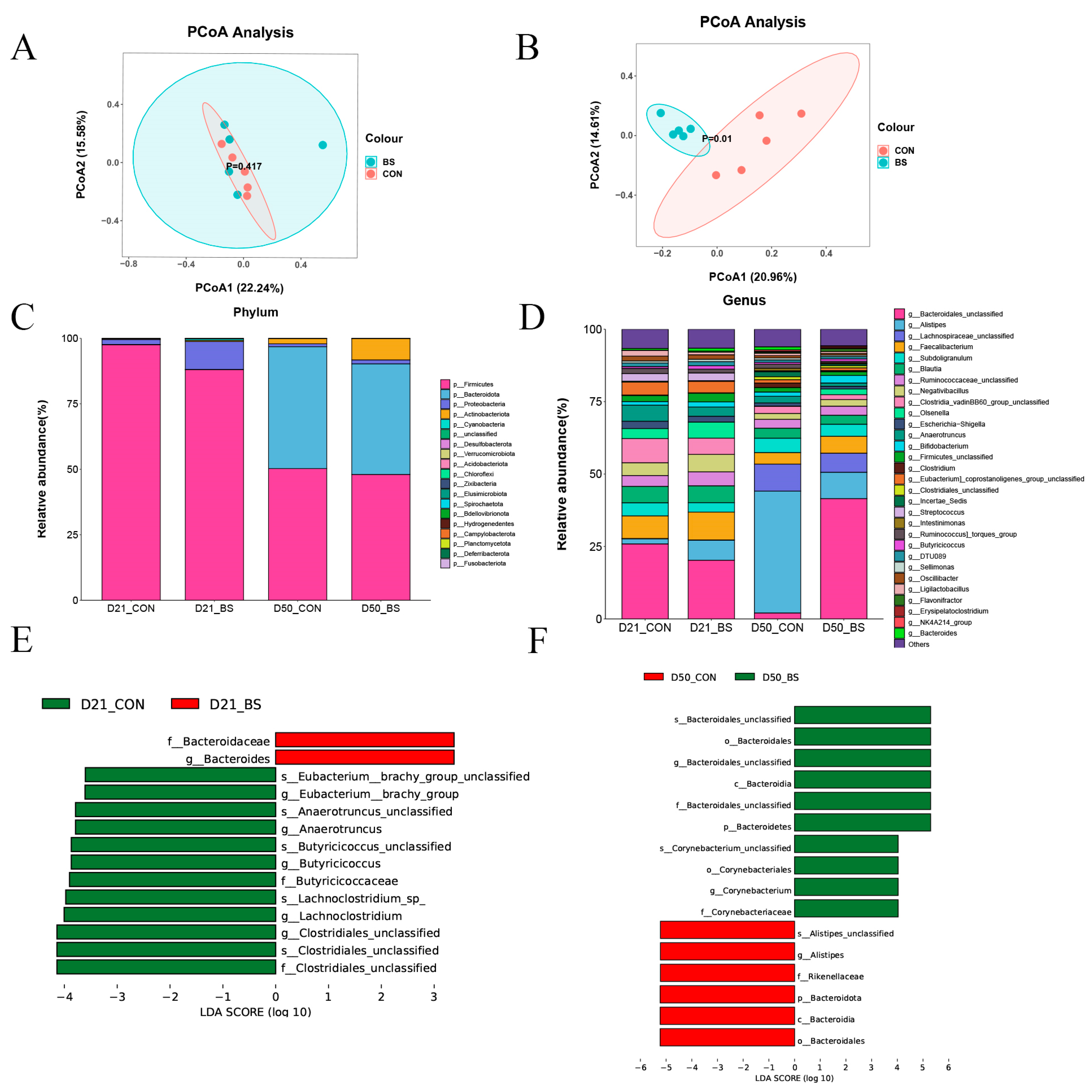
3.7. Gut Microbiota
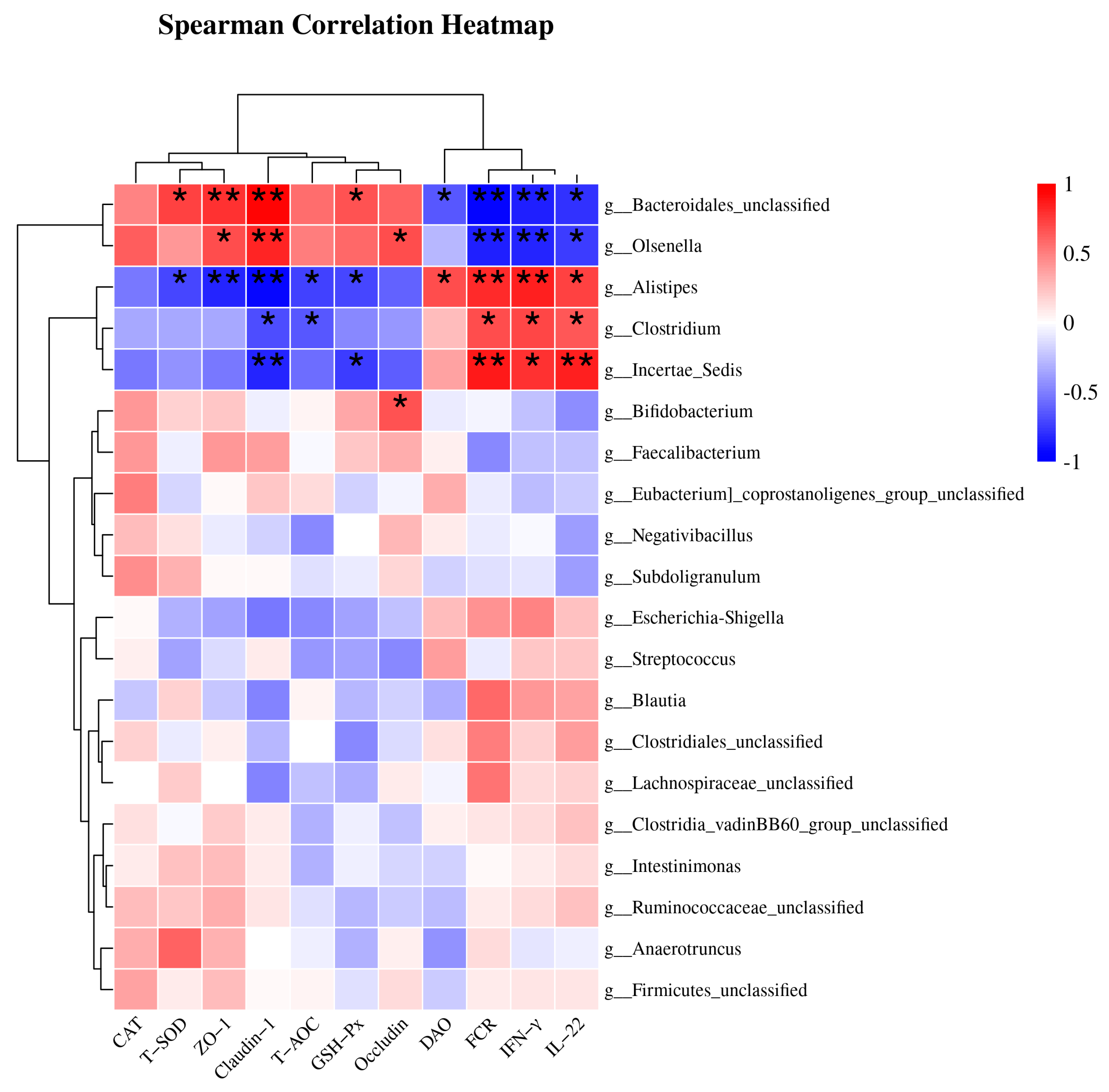
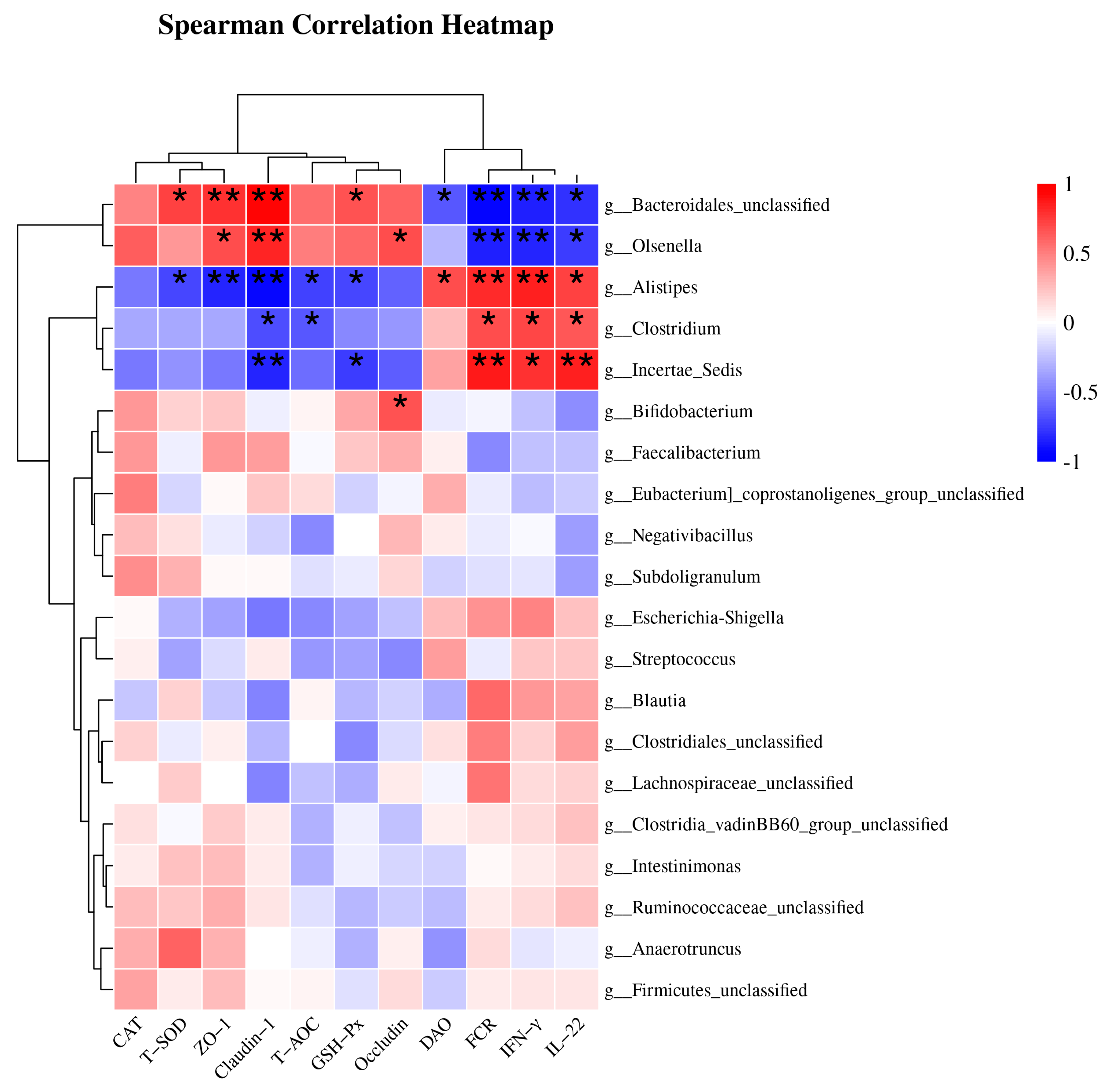
3.8. Spearman Correlation Analysis of Cecal Microbiota
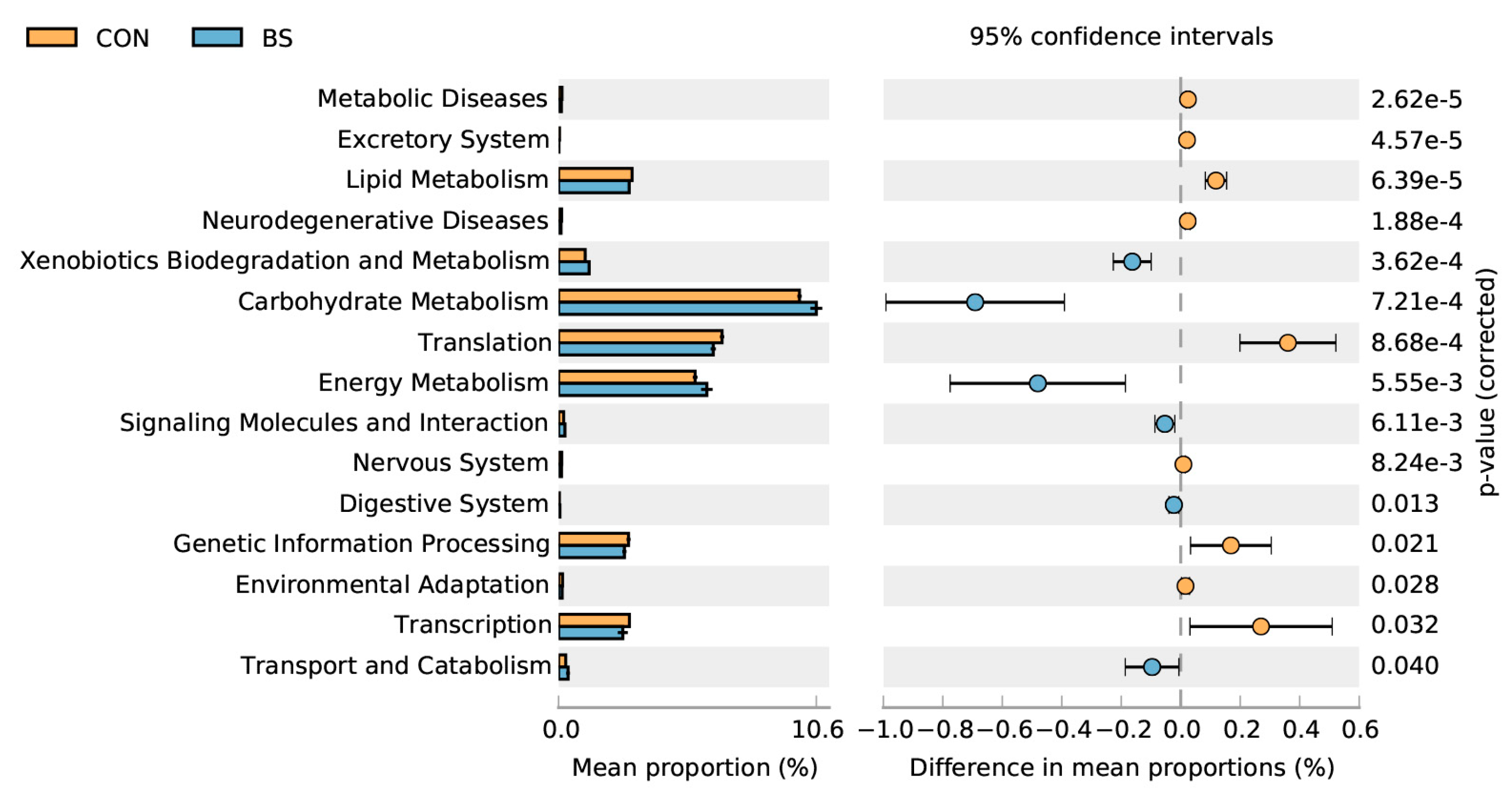
3.9. Predicted Microbial Functional Analysis by PICRUSt2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, Y.; Li, Q.; Zeng, X.; Xu, Y.; Jin, K.; Liu, J.; Cao, G. Effects of Probiotics on the Growth Performance, Antioxidant Functions, Immune Responses, and Caecal Microbiota of Broilers Challenged by Lipopolysaccharide. Front. Vet. Sci. 2022, 9, 846649. [Google Scholar] [CrossRef]
- Tong, Z.; Lei, F.; Liu, L.; Wang, F.; Guo, A. Effects of Plotytarya strohilacea Sieb. et Zuce Tannin on the Growth Performance, Oxidation Resistance, Intestinal Morphology and Cecal Microbial Composition of Broilers. Front. Vet. Sci. 2021, 8, 806105. [Google Scholar] [CrossRef]
- Al-Khalaifa, H.; Al-Nasser, A.; Al-Surayee, T.; Al-Kandari, S.; Mohammed, A. Effect of dietary probiotics and prebiotics on the performance of broiler chickens. Poult. Sci. 2019, 98, 4465–4479. [Google Scholar] [CrossRef]
- Park, I.; Lee, Y.; Goo, D.; Zimmerman, N.P.; Smith, A.H.; Rehberger, T.; Lillehoj, H.S. The effects of dietary Bacillus subtilis supplementation, as an alternative to antibiotics, on growth performance, intestinal immunity, and epithelial barrier integrity in broiler chickens infected with Eimeria maxima. Poult. Sci. 2020, 99, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.I.; Ahmad, N.; Miah, M.A. Comparative analysis of body weight and serum biochemistry in broilers supplemented with some selected probiotics and antibiotic growth promoters. J. Adv. Vet. Anim. Res. 2017, 4, 288–294. [Google Scholar] [CrossRef]
- Elleithy, E.M.M.; Bawish, B.M.; Kamel, S.; Ismael, E.; Bashir, D.W.; Hamza, D.; Fahmy, K.N.E. Influence of dietary Bacillus coagulans and/or Bacillus licheniformis-based probiotics on performance, gut health, gene expression, and litter quality of broiler chickens. Trop. Anim. Health Prod. 2023, 55, 38. [Google Scholar] [CrossRef] [PubMed]
- Luise, D.; Bosi, P.; Raff, L.; Amatucci, L.; Virdis, S.; Trevisi, P. Bacillus spp. Probiotic Strains as a Potential Tool for Limiting the Use of Antibiotics, and Improving the Growth and Health of Pigs and Chickens. Front. Microbiol. 2022, 13, 801827. [Google Scholar] [CrossRef]
- Zhou, X.; Tian, Z.; Wang, Y.; Li, W. Effect of treatment with probiotics as water additives on tilapia (Oreochromis niloticus) growth performance and immune response. Fish. Physiol. Biochem. 2010, 36, 501–509. [Google Scholar] [CrossRef]
- Lee, S.H.; Ingale, S.L.; Kim, J.S.; Kim, K.H.; Lokhande, A.; Kim, E.K.; Kwon, I.K.; Kim, Y.H.; Chae, B.J. Effects of dietary supplementation with Bacillus subtilis LS 1–2 fermentation biomass on growth performance, nutrient digestibility, cecal microbiota and intestinal morphology of weanling pig. Anim. Feed. Sci. Technol. 2014, 188, 102–110. [Google Scholar] [CrossRef]
- Mohamed, T.M.; Sun, W.; Bumbie, G.Z.; Elokil, A.A.; Mohammed, K.A.F.; Zebin, R.; Hu, P.; Wu, L.; Tang, Z. Feeding Bacillus subtilis ATCC19659 to Broiler Chickens Enhances Growth Performance and Immune Function by Modulating Intestinal Morphology and Cecum Microbiota. Front. Microbiol. 2021, 12, 798350. [Google Scholar] [CrossRef]
- Ahmat, M.; Cheng, J.; Abbas, Z.; Cheng, Q.; Fan, Z.; Ahmad, B.; Hou, M.; Osman, G.; Guo, H.; Wang, J. Effects of Bacillus amyloliquefaciens LFB112 on Growth Performance, Carcass Traits, Immune, and Serum Biochemical Response in Broiler Chickens. Antibiotics 2021, 10, 1427. [Google Scholar] [CrossRef]
- Wang, B.; Zhou, Y.; Tang, L.; Zeng, Z.; Gong, L.; Wu, Y.; Li, W.F. Effects of Bacillus amyloliquefaciens Instead of Antibiotics on Growth Performance, Intestinal Health, and Intestinal Microbiota of Broilers. Front. Vet. Sci. 2021, 8, 679368. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Cheng, Y.; Guan, L.; Zhou, Z.; Li, X.; Shi, D.; Xiao, Y. Bacillus amyloliquefaciens TL Downregulates the Ileal Expression of Genes Involved in Immune Responses in Broiler Chickens to Improve Growth Performance. Microorganisms 2021, 9, 382. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, L.; Simon, O.; Vahjen, W. Isolation and identification of mixed linked β-glucan degrading bacteria in the intestine of broiler chickens and partial characterization of respective 1,3-1,4-β-glucanase activities. J. Basic Microbiol. 2006, 46, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Bai, K.; Qiang, H.; Zhang, J.; He, J.; Tian, W. Supplemental effects of probiotic Bacillus subtilis fmbJ on growth performance, antioxidant capacity, and meat quality of broiler chickens. Poult. Sci. 2017, 96, 74–82. [Google Scholar] [CrossRef]
- Lee, T.T.; Chou, C.H.; Wang, C.; Lu, H.Y.; Yang, W.Y. Bacillus amyloliquefaciens and Saccharomyces cerevisiae feed supplements improve growth performance and gut mucosal architecture with modulations on cecal microbiota in red-feathered native chickens. Anim. Biosci. 2022, 35, 869–883. [Google Scholar] [CrossRef]
- Trevisi, P.; Luise, D.; Correa, F.; Bosi, P. Timely Control of Gastrointestinal Eubiosis: A Strategic Pillar of Pig Health. Microorganisms 2021, 9, 313. [Google Scholar] [CrossRef]
- Diaz Carrasco, J.M.; Casanova, N.A.; Fernández Miyakawa, M.E. Microbiota, Gut Health and Chicken Productivity: What Is the Connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef]
- Timmerman, H.M.; Koning, C.J.; Mulder, L.; Rombouts, F.M.; Beynen, A.C. Monostrain, multistrain and multispecies probiotics—A comparison of functionality and efficacy. Int. J. Food Microbiol. 2004, 96, 219–233. [Google Scholar] [CrossRef]
- Sun, W.; Chen, W.; Meng, K.; Cai, L.; Li, G.; Li, X.; Jiang, X. Dietary Supplementation with Probiotic Bacillus licheniformis S6 Improves Intestinal Integrity via Modulating Intestinal Barrier Function and Microbial Diversity in Weaned Piglets. Biology 2023, 12, 238. [Google Scholar] [CrossRef]
- Curone, G.; Biscarini, F.; Cotozzolo, E.; Menchetti, L.; Dal Bosco, A.; Riva, F.; Cremonesi, P.; Agradi, S.; Mattioli, S.; Castiglioni, B.; et al. Could Dietary Supplementation with Different Sources of N-3 Polyunsaturated Fatty Acids Modify the Rabbit Gut Microbiota? Antibiotics 2022, 11, 227. [Google Scholar] [CrossRef]
- Porter, N.T.; Luis, A.S.; Martens, E.C. Bacteroides thetaiotaomicron. Trends Microbiol. 2018, 26, 966–967. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Yang, J.; Li, Y.; Wu, J.; Liang, W.; Yin, B.; Tan, X.; Huang, Y.; Chai, T.; Zhang, H.; et al. Gut Microbial Signatures Can Discriminate Unipolar from Bipolar Depression. Adv. Sci. 2020, 7, 1902862. [Google Scholar] [CrossRef] [PubMed]
- Barcenilla, A.; Pryde, S.E.; Martin, J.C.; Duncan, S.H.; Stewart, C.S.; Henderson, C.; Flint, H.J. Phylogenetic relationships of butyrate-producing bacteria from the human gut. Appl. Environ. Microbiol. 2000, 66, 1654–1661. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Shen, B.; Huang, R.; Liu, H.; Zhang, W.; Song, M.; Liu, K.; Lin, X.; Chen, S.; Liu, Y.; et al. Bacteroides fragilis strain ZY-312 promotes intestinal barrier integrity via upregulating the STAT3 pathway in a radiation-induced intestinal injury mouse model. Front. Nutr. 2022, 9, 1063699. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Xia, X.; Wang, P.; Chen, S.; Yu, C.; Huang, R.; Yang, L. Induction and Amelioration of Methotrexate-Induced Gastrointestinal Toxicity are Related to Immune Response and Gut Microbiota. EBioMedicine 2018, 33, 122–133. [Google Scholar] [CrossRef]
- Suez, J.; Zmora, N.; Zilberman-Schapira, G.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Zur, M.; Regev-Lehavi, D.; Ben-Zeev Brik, R.; Federici, S.; et al. Post-Antibiotic Gut Mucosal Microbiome Reconstitution Is Impaired by Probiotics and Improved by Autologous FMT. Cell 2018, 174, 1406–1423.e1416. [Google Scholar] [CrossRef]
- Choi, H.; Na, K.J. Pan-cancer analysis of tumor metabolic landscape associated with genomic alterations. Mol. Cancer 2018, 17, 150. [Google Scholar] [CrossRef]
- Zhu, C.; Gong, L.; Huang, K.; Li, F.; Tong, D.; Zhang, H. Effect of Heat-Inactivated Compound Probiotics on Growth Performance, Plasma Biochemical Indices, and Cecal Microbiome in Yellow-Feathered Broilers. Front. Microbiol. 2020, 11, 585623. [Google Scholar] [CrossRef]
- Zhu, L.; Fu, J.; Xiao, X.; Wang, F.; Jin, M.; Fang, W.; Wang, Y.; Zong, X. Faecal microbiota transplantation-mediated jejunal microbiota changes halt high-fat diet-induced obesity in mice via retarding intestinal fat absorption. Microb. Biotechnol. 2022, 15, 337–352. [Google Scholar] [CrossRef]
- Kim, K.; He, Y.; Xiong, X.; Ehrlich, A.; Liu, Y. Dietary supplementation of Bacillus subtilis influenced intestinal health of weaned pigs experimentally infected with a pathogenic E. coli. J. Anim. Sci. Biotechnol. 2019, 10, 52. [Google Scholar] [CrossRef]
- Tang, W.; Qian, Y.; Yu, B.; Zhang, T.; Gao, J.; He, J.; Huang, Z.; Zheng, P.; Mao, X.; Luo, J. Effects of Bacillus subtilis DSM32315 supplementation and dietary crude protein level on performance, gut barrier function and microbiota profile in weaned piglets1. J. Anim. Sci. 2019, 97, 2125–2138. [Google Scholar] [CrossRef] [PubMed]
- Gunal, M.; Yayli, G.; Kaya, O.; Karahan, N.; Sulak, O. The Effects of Antibiotic Growth Promoter, Probiotic or Organic Acid Supplementation on Performance, Intestinal Microflora and Tissue of Broilers. Int. J. Poult. Sci. 2006, 5, 605–609. [Google Scholar] [CrossRef]
- Samli, H.E.; Senkoylu, N.; Koc, F.; Kanter, M.; Agma, A. Effects of Enterococcus faecium and dried whey on broiler performance, gut histomorphology and intestinal microbiota. Arch. Anim. Nutr. 2007, 61, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Zulkifli, I.; Abdulllah, N.; Azrin, N.M.; Ho, Y.W. Growth performance and immune response of two commercial broiler strains fed diets containing Lactobacillus cultures and oxytetracycline under heat stress conditions. Br. Poult. Sci. 2000, 41, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Wei, H.; Xu, C.; Xie, X.; Jiang, S.; Peng, J. Maternal Soluble Fiber Diet during Pregnancy Changes the Intestinal Microbiota, Improves Growth Performance, and Reduces Intestinal Permeability in Piglets. Appl. Environ. Microbiol. 2018, 84, e01047-18. [Google Scholar] [CrossRef]
- Okyere, S.K.; Wen, J.; Cui, Y.; Xie, L.; Gao, P.; Zhang, M.; Wang, J.; Wang, S.; Ran, Y.; Ren, Z.; et al. Bacillus toyonensis SAU-19 and SAU-20 Isolated from Ageratina adenophora Alleviates the Intestinal Structure and Integrity Damage Associated with Gut Dysbiosis in Mice Fed High Fat Diet. Front. Microbiol. 2022, 13, 820236. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, L.; Zhan, X.; Zeng, X.; Zhou, L.; Cao, G.; Chen, A.; Yang, C. Effects of dietary supplementation of probiotic, Clostridium butyricum, on growth performance, immune response, intestinal barrier function, and digestive enzyme activity in broiler chickens challenged with Escherichia coli K88. J. Anim. Sci. Biotechnol. 2016, 7, 3. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, Y.H.; Zhou, D.; Wu, Q.; Song, D.; Dicksved, J.; Wang, J.F. Oral administration of a select mixture of Bacillus probiotics affects the gut microbiota and goblet cell function in newly weaned MUC4 resistant pigs following Escherichia coli challenge. Appl. Environ. Microbiol. 2016, 83, AEM.02747-02716. [Google Scholar] [CrossRef]
- Wang, Y.; Du, W.; Lei, K.; Wang, B.; Li, W. Effects of Dietary Bacillus licheniformis on Gut Physical Barrier, Immunity, and Reproductive Hormones of Laying Hens. Probiotics Antimicrob. Proteins 2017, 9, 292–299. [Google Scholar] [CrossRef]
- Kan, L.; Guo, F.; Liu, Y.; Pham, V.H.; Guo, Y.; Wang, Z. Probiotics Bacillus licheniformis Improves Intestinal Health of Subclinical Necrotic Enteritis-Challenged Broilers. Front. Microbiol. 2021, 12, 868. [Google Scholar] [CrossRef] [PubMed]
- Otte, J.M.; Podolsky, D.K. Functional modulation of enterocytes by gram-positive and gram-negative microorganisms. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G613. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.M.; Karaca, K.; Pertile, T. Virus-induced immunosuppression in chickens. Poult. Sci. 1994, 73, 1082–1086. [Google Scholar] [CrossRef] [PubMed]
- Shini, S.; Huff, G.R.; Shini, A.; Kaiser, P. Understanding stress-induced immunosuppression: Exploration of cytokine and chemokine gene profiles in chicken peripheral leukocytes. Poult. Sci. 2010, 115, 117–123. [Google Scholar] [CrossRef]
- Gomes, A.V.S.; Quinteiro-Filho, W.M.; Ribeiro, A.; Ferraz-De-Paula, V.; Pinheiro, M.L.; Baskeville, E.; Akamine, A.T.; Astolfi-Ferreira, C.S.; Ferreira, A.J.P.; Palermo-Neto, J. Overcrowding stress decreases macrophage activity and increases Salmonella Enteritidis invasion in broiler chickens. Brain Behav. Immun. 2014, 40, e30–e31. [Google Scholar] [CrossRef]
- Hsieh, W.C.; Hsu, T.S.; Chang, Y.J.; Lai, M.Z. IL-6 receptor blockade corrects defects of XIAP-deficient regulatory T cells. Nat. Commun. 2018, 9, 463. [Google Scholar] [CrossRef]
- Gao, P.; Ma, C.; Sun, Z.; Wang, L.; Huang, S.; Su, X.; Xu, J.; Zhang, H. Feed-additive probiotics accelerate yet antibiotics delay intestinal microbiota maturation in broiler chicken. Microbiome 2017, 5, 91. [Google Scholar] [CrossRef]
- Isolauri, E.; Sütas, Y.; Kankaanp??, P.; Arvilommi, H.; Salminen, S. Probiotics: Effects on immunity. Am. J. Clin. Nutr. 2001, 73, 444S. [Google Scholar] [CrossRef]
- Borchers, A.T.; Selmi, C.; Meyers, F.J.; Keen, C.L.; Gershwin, M.E. Probiotics and immunity. J. Gastroenterol. 2009, 44, 26–46. [Google Scholar] [CrossRef]
- Li, J.; Sung, C.Y.J.; Lee, N.; Ni, Y.; El-Nezami, H. Probiotics modulated gut microbiota suppresses hepatocellular carcinoma growth in mice. Proc. Natl. Acad. Sci. USA 2016, 113, E1306. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Huang, Q.; Mao, Y.; Cui, Z.; Li, W. Immunomodulatory effects of Bacillus subtilis (natto) B4 spores on murine macrophages. Microbiol. Immunol. 2012, 56, 817–824. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.F.; An, L.J.; Jiang, B.; Guan, S.; Bao, Y.M. Alpinia protocatechuic acid protects against oxidative damage in vitro and reduces oxidative stress in vivo. Neurosci. Lett. 2006, 403, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Zeng, X.; Tan, F.; Li, W.; Li, C.; Song, Y.; Zhao, X. Effect of insect tea on D-galactose-induced oxidation in mice and its mechanisms. Food Sci. Nutr. 2019, 7, 4105–4115. [Google Scholar] [CrossRef]
- Zhao, Y.; Zeng, D.; Wang, H.; Qing, X.; Sun, N.; Xin, J.; Luo, M.; Khalique, A.; Pan, K.; Shu, G.; et al. Dietary Probiotic Bacillus licheniformis H2 Enhanced Growth Performance, Morphology of Small Intestine and Liver, and Antioxidant Capacity of Broiler Chickens against Clostridium perfringens-Induced Subclinical Necrotic Enteritis. Probiotics Antimicrob. Proteins 2020, 12, 883–895. [Google Scholar] [CrossRef]
- Gong, L.; Wang, B.; Mei, X.; Xu, H.; Qin, Y.; Li, W.; Zhou, Y. Effects of three probiotic Bacillus on growth performance, digestive enzyme activities, antioxidative capacity, serum immunity, and biochemical parameters in broilers. Anim. Sci. J. 2018, 89, 1561–1571. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets | Starter (1–20 Days) | Finisher (21–50 Days) |
|---|---|---|
| Ingredients (g/kg) | ||
| Corn | 58.75 | 57.66 |
| Soybean meal (43% CP) | 27.70 | 27.21 |
| Corn gluten meal (60% CP) | 5.00 | 5.00 |
| Limestone | 1.43 | 1.22 |
| Calcium hydrogen phosphate (16.5%) | 1.10 | 1.04 |
| L-lysine Sulfate (70%) | 0.66 | 0.39 |
| DL-methionine (98.5%) | 0.33 | 0.21 |
| NaCl | 0.28 | 0.32 |
| L-Threonine | 0.16 | 0.06 |
| Peanut meal | 3.00 | 0 |
| Choline chloride (50%) | 0.08 | 0.08 |
| Premix 1 | 0.35 | 0.35 |
| Phytase (20,000 IU) | 0.01 | 0.01 |
| Sodium humate | 0 | 0.15 |
| Lard | 1.15 | 6.30 |
| Total (%) | 100 | 100 |
| Nutrient content 2 | ||
| ME (Kcal/kg) | 2941.61 | 3231.28 |
| CP (g/kg) | 21.92 | 19.61 |
| CEE (g/kg) | 3.65 | 8.87 |
| CF (g/kg) | 2.51 | 2.33 |
| Ca (g/kg) | 0.90 | 0.80 |
| Total P (%) | 0.57 | 0.54 |
| Lys (%) | 1.48 | 1.21 |
| Met (%) | 0.64 | 0.52 |
| Cys (%) | 0.30 | 0.27 |
| Met + Cys (%) | 0.94 | 0.78 |
| L-Threonine | 0.92 | 0.79 |
| Gene | GenBank | Sequence (5′–3′) | TM °C |
|---|---|---|---|
| β-actin | NM_205518.2 | F: CATTGTCCACCGCAAATGCT R: AAGCCATGCCAATCTCGTCT | 57.2 |
| IL-6 | NM_204628.2 | F: CAGGACGAGATGTGCAAGAA R: TAGCACAGAGACTCGACGTT | 56.6 |
| IL-10 | NM_001004414.4 | F: CGCTGTCACCGCTTCTTCA R: CTTTGTTCTCATCCATCTTCTC | 57.0 |
| IL-1β | XM_046931582.1 | F: CGACATCAACCAGAAGTGCTT R: GTCCAGGCGGTAGAAGATGA | 56.8 |
| IL-22 | NM_001199614.1 | F: GCCCTACATCAGGAATCGCA R: TCTGAGAGCCTGGCCATTTC | 57.8 |
| IL-17A | NM_204460.2 | F: GAAGGTGATACGGCCAGGAC R: TGGGTTAGGCATCCAGCATC | 56.8 |
| IFN-β | NM_001024836.2 | F: TGCAACCATCTTCGTCACCA R: GGAGGTGGAGCCGTATTCT | 56.68 |
| IFN-γ | NM_205149.2 | F: ACACTGACAAGTCAAAGCCGC R: AGTCGTTCATCGGGAGCTTG | 58.6 |
| TNF-α | XM_046927265.1 | F: TGTGTATGTGCAGCAACCCGTAGT R: GGCATTGCAATTTGGACAGAAGT | 57.8 |
| MUC-2 | XM_040673077.2 | F: TTCATGATGCCTGCTCTTGTG R: CCTGAGCCTTGGTACATTCTTGT | 58.0 |
| ZO-1 | XM_046925214.1 | F: CTTCAGGTGTTTCTCTTCCTCCTC R: CTGTGGTTTCATGGCTGGATC | 56.6 |
| Claudin-1 | NM_001013611.2 | F: GGTATGGCAACAGAGTGGCT R: CAGCCAATGAAGAGGGCTGA | 57.0 |
| Occludin | XM_046904540.1 | F: GATGGACAGCATCAACGACC R: CATGCGCTTGATGTGGAAGA | 58.0 |
| Items | CON | BS | p-Value |
|---|---|---|---|
| 1 d BW, g | 36.20 ± 0.01 | 36.20 ± 0.01 | - |
| 21 d BW, g | 535.28 ± 21.96 | 543.06 ± 14.35 | 0.635 |
| 50 d BW, g | 2040.00 ± 115.33 | 2092.67 ± 55.15 | 0.515 |
| 1–21 d | |||
| ADG, g/d | 24.95 ± 1.10 | 25.34 ± 0.72 | 0.634 |
| ADFI, g/d | 35.20 ± 1.88 | 34.79 ± 0.86 | 0.746 |
| FCR | 1.41 ± 0.02 | 1.37 ± 0.04 | 0.180 |
| 21–50 d | |||
| ADG, g/d | 51.88 ± 3.32 | 53.44 ± 1.42 | 0.499 |
| ADFI, g/d | 109.63 ± 6.66 | 103.03 ± 2.30 | 0.093 |
| FCR | 2.12 ± 0.08 | 1.93 ± 0.02 | 0.015 |
| 1–50 d | |||
| ADG, g/d | 40.83 ± 2.46 | 41.90 ± 1.23 | 0.537 |
| ADFI, g/d | 79.25 ± 3.52 | 75.18 ± 1.69 | 0.145 |
| FCR | 1.94 ± 0.06 | 1.80 ± 0.03 | 0.020 |
| Items | CON | BS | p-Value |
|---|---|---|---|
| jejunum | |||
| 21 d Villus height, µm | 1029.94 ± 45.42 | 1062.01 ± 120.92 | 0.594 |
| 21 d Crypt depth, µm | 111.80 ± 12.99 | 120.75 ± 15.37 | 0.349 |
| 21 d V/C | 9.28 ± 0.85 | 9.03 ± 2.42 | 0.833 |
| 50 d Villus height, µm | 1090.97 ± 17.91 | 1204.35 ± 38.96 | 0.001 |
| 50 d Crypt depth, µm | 111.59 ± 13.17 | 118.24 ± 12.82 | 0.442 |
| 50 d V/C | 9.87 ± 0.97 | 10.29 ± 1.04 | 0.557 |
| ileum | |||
| 21 d Villus height, µm | 589.94 ± 42.97 | 623.35 ± 45.29 | 0.266 |
| 21 d Crypt depth, µm | 81.80 ± 5.51 | 79.70 ± 2.57 | 0.463 |
| 21 d V/C | 7.22 ± 0.47 | 7.82 ± 0.46 | 0.079 |
| 50 d Villus height, µm | 780.74 ± 53.21 | 873.91 ± 14.20 | 0.005 |
| 50 d Crypt depth, µm | 115.59 ± 4.82 | 112.89 ± 5.56 | 0.435 |
| 50 d V/C | 6.77 ± 0.60 | 7.75 ± 0.29 | 0.011 |
| Items | CON | BS | p-Value |
|---|---|---|---|
| serum | |||
| D 21 | |||
| IL-2, ng/L | 36.85 ± 11.64 | 44.90 ± 13.98 | 0.351 |
| IL-6, ng/L | 61.11 ± 25.48 | 31.17 ± 13.88 | 0.024 |
| TNF-α, ng/L | 132.61 ± 14.63 | 139.84 ± 15.07 | 0.463 |
| TGF-β, ng/L | 3110.92 ± 247.55 | 2899.92 ± 185.31 | 0.166 |
| IL-10, ng/L | 11.40 ± 5.91 | 12.34 ± 3.99 | 0.775 |
| sIgA, ng/L | 234.49 ± 38.47 | 255.39 ± 51.42 | 0.487 |
| D 50 | |||
| IL-2, ng/L | 21.71 ± 5.24 | 25.50 ± 7.64 | 0.388 |
| IL-6, ng/L | 90.20 ± 14.48 | 87.11 ± 13.12 | 0.733 |
| TNF-α, ng/L | 191.10 ± 14.90 | 167.30 ± 15.75 | 0.040 |
| TGF-β, ng/L | 2630.09 ± 264.19 | 2448.74 ± 152.73 | 0.221 |
| IL-10, ng/L | 4.54 ± 1.72 | 4.27 ± 2.99 | 0.869 |
| sIgA, ng/L | 169.85 ± 12.65 | 173.92 ± 28.14 | 0.776 |
| jejunal mucosa | |||
| D 21 | |||
| IL-6 | 1.09 ± 0.10 | 1.23 ± 0.54 | 0.636 |
| IL-10 | 1.04 ± 0.13 | 1.88 ± 0.42 | 0.009 |
| IL-1β | 1.04 ± 0.03 | 1.73 ± 0.68 | 0.089 |
| IL-22 | 1.10 ± 0.09 | 1.53 ± 0.26 | 0.019 |
| IL-17A | 1.02 ± 0.04 | 1.34 ± 0.72 | 0.418 |
| IFN-γ | 1.05 ± 0.09 | 1.78 ± 0.53 | 0.035 |
| TNF-α | 1.05 ± 0.07 | 1.40 ± 0.64 | 0.312 |
| D 50 | |||
| IL-6 | 1.02 ± 0.05 | 0.63 ± 0.35 | 0.069 |
| IL-10 | 1.02 ± 0.05 | 1.01 ± 0.20 | 0.932 |
| IL-1β | 1.03 ± 0.04 | 0.64 ± 0.36 | 0.092 |
| IL-22 | 1.07 ± 0.10 | 0.33 ± 0.20 | 0.001 |
| IL-17A | 1.03 ± 0.04 | 0.60 ± 0.38 | 0.067 |
| IFN-γ | 1.04 ± 0.09 | 0.64 ± 0.32 | 0.049 |
| TNF-α | 1.06 ± 0.07 | 1.35 ± 0.36 | 0.166 |
| Items | CON | BS | p-Value |
|---|---|---|---|
| serum | |||
| D 21 | |||
| MDA, nmol/mL | 5.01 ± 1.50 | 3.93 ± 1.02 | 0.787 |
| T-AOC, U/mL | 3.84 ± 1.45 | 6.38 ± 1.16 | 0.016 |
| CAT, U/mL | 14.60 ± 2.61 | 21.41 ± 2.92 | 0.006 |
| T-SOD, U/mL | 28.52 ± 9.76 | 42.87 ± 7.28 | 0.030 |
| GSH-Px, umol/L | 708.93 ± 235.55 | 946.41 ± 357.81 | 0.250 |
| D 50 | |||
| MDA, nmol/mL | 3.70 ± 1.11 | 2.84 ± 0.53 | 0.156 |
| T-AOC, U/mL | 3.70 ± 1.13 | 7.39 ± 3.14 | 0.039 |
| CAT, U/mL | 3.80 ± 0.77 | 5.16 ± 0.87 | 0.031 |
| T-SOD, U/mL | 43.95 ± 7.04 | 53.99 ± 4.33 | 0.026 |
| GSH-Px, umol/L | 1178.90 ± 273.97 | 1787.62 ± 230.15 | 0.003 |
| liver | |||
| D 21 | |||
| MDA, nmol/mgprot | 4.73 ± 1.13 | 4.35 ± 1.39 | 0.648 |
| T-AOC, U/mgprot | 6.49 ± 0.26 | 7.38 ± 1.04 | 0.105 |
| CAT, U/mgprot | 18.50 ± 6.43 | 26.49 ± 5.49 | 0.068 |
| T-SOD, U/mgprot | 151.88 ± 34.60 | 222.66 ± 41.59 | 0.019 |
| GSH-Px, umol/mgprot | 52.14 ± 7.83 | 73.29 ± 10.26 | 0.006 |
| D 50 | |||
| MDA, nmol/mgprot | 5.33 ± 0.69 | 5.22 ± 0.44 | 0.772 |
| T-AOC, U/mgprot | 5.34 ± 0.31 | 6.63 ± 1.03 | 0.029 |
| CAT, U/mgprot | 16.99 ± 3.65 | 27.49 ± 8.41 | 0.034 |
| T-SOD, U/mgprot | 268.32 ± 33.59 | 350.72 ± 57.95 | 0.025 |
| GSH-Px, umol/mgprot | 63.81 ± 8.16 | 87.35 ± 20.96 | 0.047 |
| Items | CON | BS | p-Value |
|---|---|---|---|
| D 21 | |||
| Triglyceride, mmol/L | 0.50 ± 0.08 | 0.66 ± 0.06 | 0.004 |
| Total cholesterol, mmol/L | 4.21 ± 0.47 | 3.78 ± 0.42 | 0.121 |
| High-density lipoprotein, mmol/L | 9.46 ± 1.15 | 8.89 ± 1.19 | 0.423 |
| Low density lipoprotein, mmol/L | 1.01 ± 0.13 | 1.07 ± 0.13 | 0.477 |
| Albumin, g/L | 20.87 ± 1.55 | 18.03 ± 1.77 | 0.049 |
| Uric acid, umol/L | 188.43 ± 56.90 | 217.13 ± 42.72 | 0.346 |
| Lactate dehydrogenase, U/L | 571.51 ± 52.76 | 525.00 ± 69.27 | 0.220 |
| Alkaline phosphatase, mg/L | 609.34 ± 129.37 | 526.01 ± 82.99 | 0.214 |
| Aspartate aminotransferase, U/L | 27.66 ± 7.30 | 36.97 ± 6.59 | 0.043 |
| Alanine aminotransferase, U/L | 34.34 ± 7.66 | 45.43 ± 22.07 | 0.271 |
| D 50 | |||
| Triglyceride, mmol/L | 0.75 ± 0.19 | 0.86 ± 0.20 | 0.317 |
| Total cholesterol, mmol/L | 4.26 ± 0.67 | 4.75 ± 1.48 | 0.476 |
| High-density lipoprotein, mmol/L | 3.62 ± 1.51 | 5.65 ± 0.43 | 0.010 |
| Low density lipoprotein, mmol/L | 3.43 ± 0.86 | 2.97 ± 1.07 | 0.432 |
| Albumin, g/L | 20.87 ± 4.03 | 20.28 ± 2.84 | 0.775 |
| Uric acid, umol/L | 353.52 ± 72.75 | 320.46 ± 60.03 | 0.411 |
| Lactate dehydrogenase, U/L | 607.26 ± 25.79 | 566.99 ± 48.10 | 0.101 |
| Alkaline phosphatase, mg/L | 155.56 ± 29.88 | 174.18 ± 40.97 | 0.389 |
| Aspartate aminotransferase, U/L | 15.36 ± 3.88 | 22.65 ± 8.07 | 0.074 |
| Alanine aminotransferase, U/L | 21.12 ± 12.45 | 13.93 ± 9.18 | 0.281 |
| Items | CON | BS | p-Value |
|---|---|---|---|
| D 21 | |||
| Chao 1 | 502.72 ± 124.09 | 495.09 ± 165.60 | 0.936 |
| Shannon | 6.96 ± 0.30 | 6.41 ± 1.40 | 0.411 |
| Simpson | 0.98 ± 0.01 | 0.96 ± 0.04 | 0.409 |
| Goods_coverage | 1.00 | 1.00 | - |
| Pielou_e | 0.81 ± 0.07 | 0.72 ± 0.12 | 0.199 |
| Observed_otus | 501.80 ± 123.43 | 494.40 ± 165.47 | 0.938 |
| D 50 | |||
| Chao 1 | 408.64 ± 51.88 | 441.90 ± 69.34 | 0.415 |
| Shannon | 4.83 ± 0.42 | 5.26 ± 0.54 | 0.206 |
| Simpson | 0.84 ± 0.06 | 0.89 ± 0.07 | 0.254 |
| Goods_coverage | 1.00 | 1.00 | - |
| Pielou_e | 0.56 ± 0.04 | 0.60 ± 0.08 | 0.313 |
| Observed_otus | 406.60 ± 51.85 | 440.00 ± 69.27 | 0.413 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Xiao, G.; Wang, Q.; Zhang, Q.; Tian, J.; Li, W.; Gong, L. Effects of Dietary Bacillus subtilis HC6 on Growth Performance, Antioxidant Capacity, Immunity, and Intestinal Health in Broilers. Animals 2023, 13, 2915. https://doi.org/10.3390/ani13182915
Liu S, Xiao G, Wang Q, Zhang Q, Tian J, Li W, Gong L. Effects of Dietary Bacillus subtilis HC6 on Growth Performance, Antioxidant Capacity, Immunity, and Intestinal Health in Broilers. Animals. 2023; 13(18):2915. https://doi.org/10.3390/ani13182915
Chicago/Turabian StyleLiu, Shun, Gengsheng Xiao, Qi Wang, Qingyang Zhang, Jinpeng Tian, Weifen Li, and Li Gong. 2023. "Effects of Dietary Bacillus subtilis HC6 on Growth Performance, Antioxidant Capacity, Immunity, and Intestinal Health in Broilers" Animals 13, no. 18: 2915. https://doi.org/10.3390/ani13182915
APA StyleLiu, S., Xiao, G., Wang, Q., Zhang, Q., Tian, J., Li, W., & Gong, L. (2023). Effects of Dietary Bacillus subtilis HC6 on Growth Performance, Antioxidant Capacity, Immunity, and Intestinal Health in Broilers. Animals, 13(18), 2915. https://doi.org/10.3390/ani13182915