
Effects of Concentrate Feeding Sequence on Growth Performance, Nutrient Digestibility, VFA Production, and Fecal Microbiota of Weaned Donkeys

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and Feeding
2.2. Major Reagents, Instruments
2.3. Samples Collection
2.4. Growth Performance, Nutrient Digestibility, and VFA
2.5. Genomic DNA Extraction, PCR Amplification and Purification, Sequencing
2.6. Bioinformatics Analysis
2.7. Statistical Analysis
3. Results
3.1. Effects of Different Concentrate Feeding Sequences on Growth Performance
3.2. Effects of Different Concentrate Feeding Sequences on Apparent Digestibility
3.3. Effect of Different Concentrate Feeding Sequences on VFA Concentration
3.4. Effects of Different Concentrate Feeding Sequences on Intestinal Microflora Composition
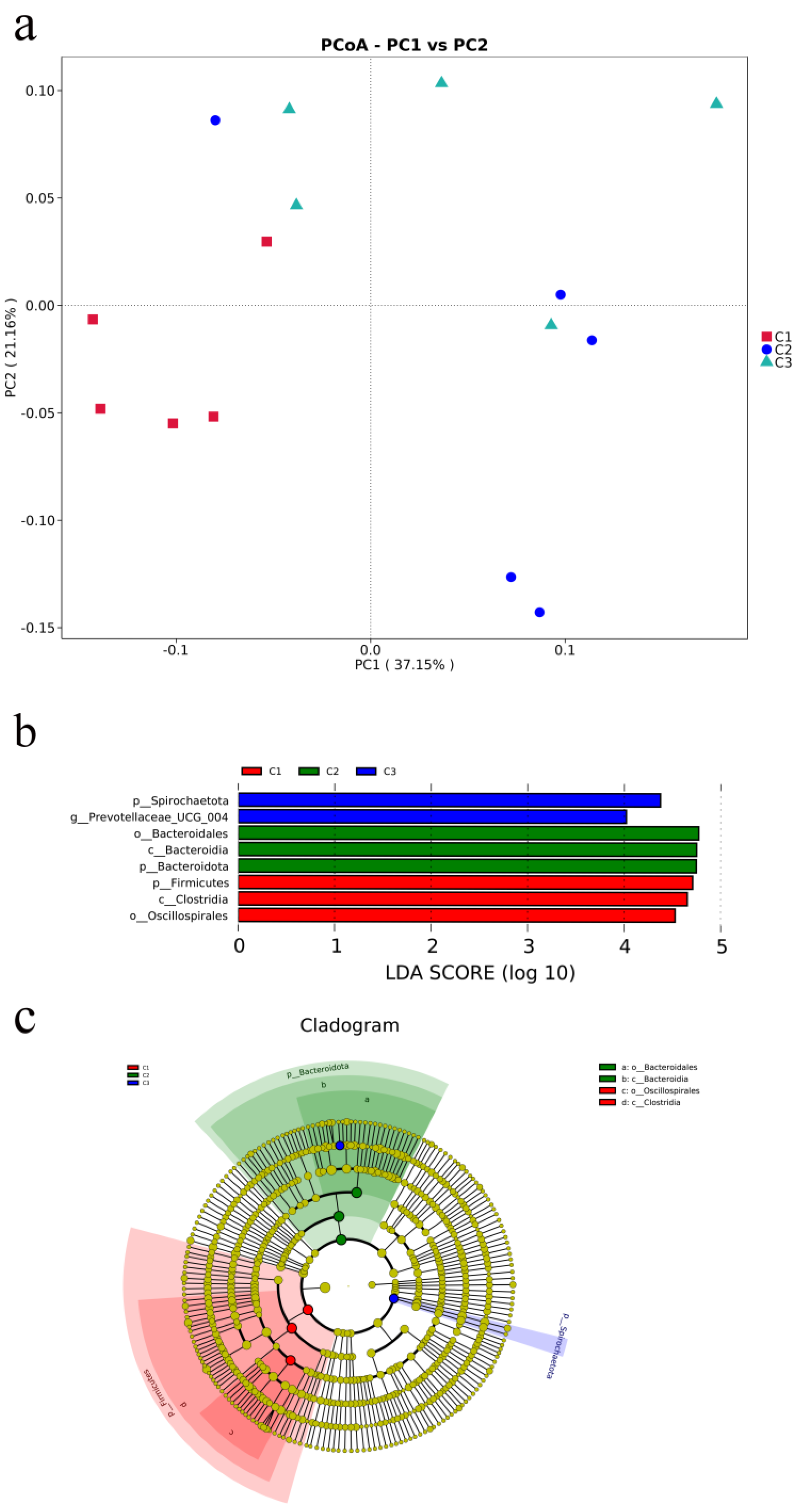
3.4.1. Analysis of α-Diversity of Species
3.4.2. Relative Abundance of Microbial Species at Phylum and Genus Levels
3.4.3. Microbial Community Analysis of Fecal Microbiota
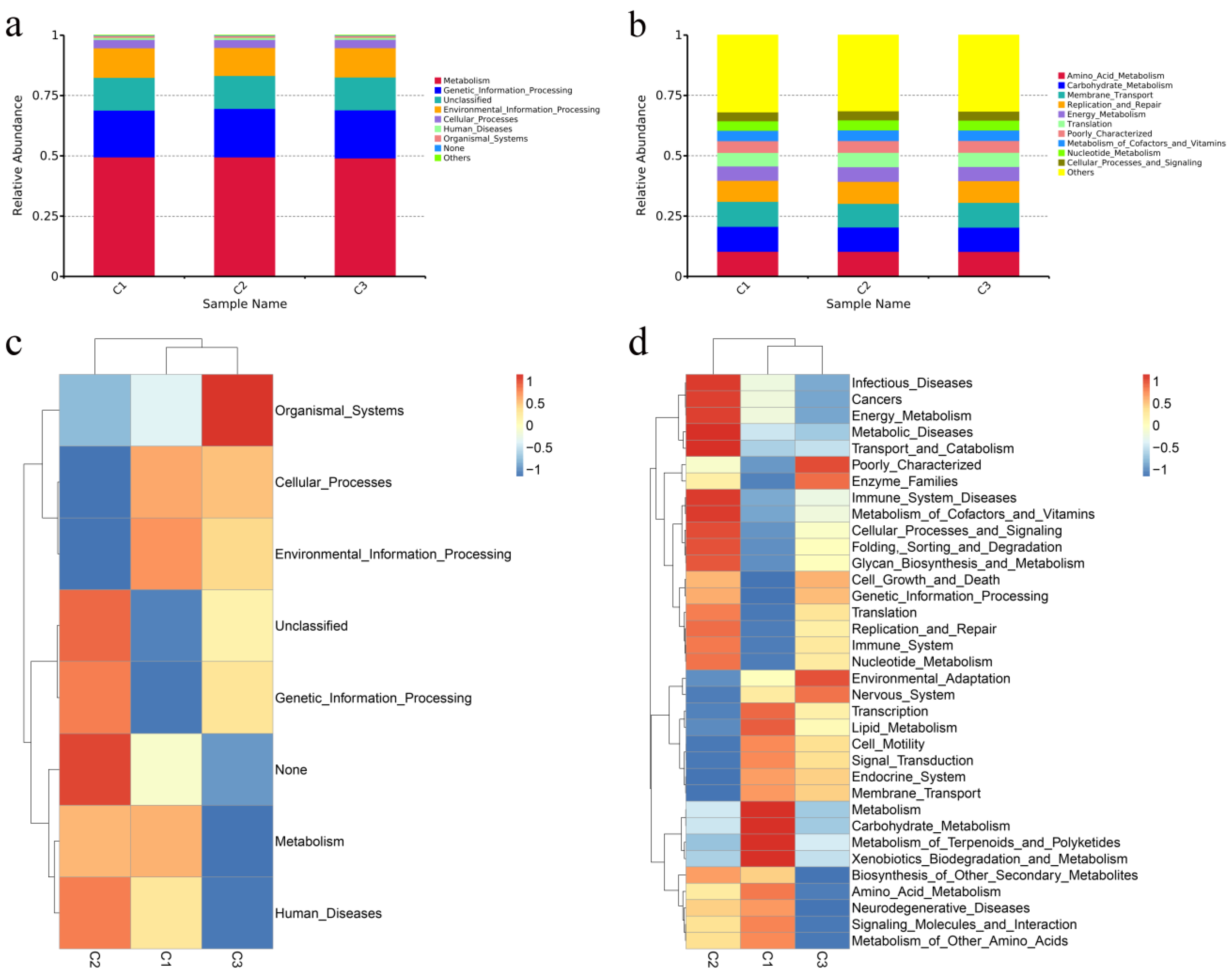
3.4.4. Functional Predictions of the Fecal Microbiota Using PICRUSt
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.F.; Sun, F.F.; Wan, F.C.; Zhao, H.B.; Liu, X.M.; You, W.; Cheng, H.J.; Liu, G.F.; Tan, X.W.; Song, E.L. Effects of Three Feeding Systems on Production Performance, Rumen Fermentation and Rumen Digesta Particle Structure of Beef Cattle. Asian-Australas. J. Anim. Sci. 2016, 29, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, H.; Guo, X. Effects of total mixed ration with various silage on growth performance, serum parameters, ruminal fermentation, and bacteria community profile in beef cattle. Food Sci. Nutr. 2021, 9, 5959–5970. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Choi, S.H.; Park, S.K.; Lee, S.S.; Choi, C.W. Effect of corn grain particle size on ruminal fermentation and blood metabolites of Holstein steers fed total mixed ration. Asian-Australas. J. Anim. Sci. 2018, 31, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, S.M.; Lee, J.; Kim, E.J. Feeding strategies with total mixed ration and concentrate may improve feed intake and carcass quality of Hanwoo steers. J. Anim. Sci. Technol. 2021, 63, 1086–1097. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, M.; Boisson, T.; Erhard, M.H.; Zeitler-Feicht, M.H. Common Feeding Practices Pose a Risk to the Welfare of Horses When Kept on Non-Edible Bedding. Animals 2020, 10, 411. [Google Scholar] [CrossRef]
- Wang, K.Y. Effects of Different Concentrate to Forage Ration in TMR on Digestive Metabolism of Nutrient Matter and Production Performance in Sika Deer. Master’s Thesis, Chinese Academy of Agricultural Sciences: Beijing, China, 2008. [Google Scholar]
- Sahu, N.P.; Kamra, D.N. Microbial Eco-system of the Gastro-intestinal Tract of Wild Herbivorous Animals. J. Appl. Anim. Res. 2002, 21, 207–230. [Google Scholar] [CrossRef]
- Liu, X.; Fan, H.; Ding, X.; Hong, Z.; Nei, Y.; Liu, Z.; Li, G.; Guo, H. Analysis of the gut microbiota by high-throughput sequencing of the V5-V6 regions of the 16S rRNA gene in donkey. Curr. Microbiol. 2014, 68, 657–662. [Google Scholar] [CrossRef]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef]
- Khafipour, E.; Li, S.; Tun, H.M.; Derakhshani, H.; Moossavi, S.; Plaizier, J.C. Effects of grain feeding on microbiota in the digestive tract of cattle. Anim. Front. 2016, 6, 13–19. [Google Scholar] [CrossRef]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Malmuthuge, N.; Guan, L.L. Understanding host-microbial interactions in rumen: Searching the best opportunity for microbiota manipulation. J. Anim. Sci. Biotechnol. 2017, 8, 8. [Google Scholar] [CrossRef]
- Li, F.; Hitch, T.C.A.; Chen, Y.; Creevey, C.J.; Guan, L.L. Comparative metagenomic and metatranscriptomic analyses reveal the breed effect on the rumen microbiome and its associations with feed efficiency in beef cattle. Microbiome 2019, 7, 6. [Google Scholar] [CrossRef]
- Sorensen, R.J.; Drouillard, J.S.; Douthit, T.L.; Ran, Q.; Marthaler, D.G.; Kang, Q.; Vahl, C.I.; Lattimer, J.M. Effect of hay type on cecal and fecal microbiome and fermentation parameters in horses. J. Anim. Sci. 2021, 99, skaa407. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.C.; Whitfield-Cargile, C.M.; Madrigal, R.G.; Cohen, N.D. Comparison of the microbiome, metabolome, and lipidome of obese and non-obese horses. PLoS ONE 2019, 14, e0215918. [Google Scholar] [CrossRef] [PubMed]
- Zarrinpar, A.; Chaix, A.; Yooseph, S.; Panda, S. Diet and feeding pattern affect the diurnal dynamics of the gut microbiome. Cell Metab. 2014, 20, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Xue, D.; Chen, H.; Zhao, X.; Xu, S.; Hu, L.; Xu, T.; Jiang, L.; Zhan, W. Rumen prokaryotic communities of ruminants under different feeding paradigms on the Qinghai-Tibetan Plateau. Syst Appl. Microbiol. 2017, 40, 227–236. [Google Scholar] [CrossRef]
- Cappai, M.G.; Picciau, M.; Pinna, W. An integrated approach towards the nutritional assessment of the Sardinian donkey: A tool for clinical nutritionists. Ital. J. Anim. Sci. 2013, 12, e29. [Google Scholar] [CrossRef]
- AOAC. Official methods of analysis. In Association of Official Analytical Chemists, 15th ed.; AOAC International: Arlington, VA, USA, 1990. [Google Scholar]
- Hassanat, F.; Gervais, R.; Benchaar, C. Methane production, ruminal fermentation characteristics, nutrient digestibility, nitrogen excretion, and milk production of dairy cows fed conventional or brown midrib corn silage. J. Dairy Sci. 2017, 100, 2625–2636. [Google Scholar] [CrossRef]
- Han, X.; Guo, J.; You, Y.; Yin, M.; Ren, C.; Zhan, J.; Huang, W. A fast and accurate way to determine short chain fatty acids in mouse feces based on GC-MS. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1099, 73–82. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef]
- Yoakam, S.; Kirkham, W.; Beeson, W. Effect of protein level on growth in young ponies. J. Anim. Sci. 1978, 46, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Kiani, A.; Hoseini, F.; Ghorbaninejad, P.; Azarfar, A.; Kreuzer, M.; Azizi, A. Interaction between the sequence of feeding of hay and concentrate, and boiling of barley on feed intake, the activity of hydrolytic enzymes and fermentation in the hindgut of Arabian mares. J. Anim. Physiol. Anim. Nutr. 2018, 102, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.L. Effects of Feeding Mode on the Growth Performance, Feed Digestibility and Carcass Quality of Dezhou Donkey. Master’s Thesis, Liaocheng University, Liaocheng, China, 2021. [Google Scholar]
- Moore-Colyer, M.; Tuthill, P.; Bannister, I.; Daniels, S. Growth rates of thoroughbred foals and in vitro gut health parameters when fed a cereal or an all-fiber creep feed. J. Equine Vet. Sci. 2020, 93, 103191. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.H.; Xin, P.P.; Wen, H.M.; Sun, P.; Du, X.X.; Zhang, S.; Jiang, B.Y. Effect of feed types on growth performance and serum biochemical indexesof donkey foal. Feed. Res. 2021, 44, 103–106. [Google Scholar] [CrossRef]
- Yue, Y.; Li, L.; Tong, M.; Li, S.; Zhao, Y.; Guo, X.; Guo, Y.; Shi, B.; Yan, S. Effect of varying dietary crude protein level on milk production, nutrient digestibility, and serum metabolites by lactating donkeys. Animals 2022, 12, 2066. [Google Scholar] [CrossRef]
- Sadet-Bourgeteau, S.; Philippeau, C.; Julliand, V. Effect of concentrate feeding sequence on equine hindgut fermentation parameters. Animal 2017, 11, 1146–1152. [Google Scholar] [CrossRef] [PubMed]
- Jouany, J.-P.; Gobert, J.; Medina, B.; Bertin, G.; Julliand, V. Effect of live yeast culture supplementation on apparent digestibility and rate of passage in horses fed a high-fiber or high-starch diet1. J. Anim. Sci. 2008, 86, 339–347. [Google Scholar] [CrossRef]
- Raspa, F.; Dinardo, F.R.; Vervuert, I.; Bergero, D.; Bottero, M.T.; Pattono, D.; Dalmasso, A.; Vinassa, M.; Valvassori, E.; Bruno, E. A Fibre-vs. cereal grain-based diet: Which is better for horse welfare? Effects on intestinal permeability, muscle characteristics and oxidative status in horses reared for meat production. J. Anim. Physiol. Anim. Nutr. 2022, 106, 313–326. [Google Scholar] [CrossRef]
- McLean, B.; Hyslop, J.; Longland, A.; Cuddeford, D.; Hollands, T. Development of the mobile bag technique to determine the degradation kinetics of purified starch sources in the pre-caecal segment of the equine digestive tract. Proc. Br. Soc. Anim. Sci. 1999, 1999, 138. [Google Scholar] [CrossRef]
- McLean, B.; Hyslop, J.; Longland, A.; Cuddeford, D.; Hollands, T. Physical processing of barley and its effects on intra-caecal fermentation parameters in ponies. Anim. Feed. Sci. Technol. 2000, 85, 79–87. [Google Scholar] [CrossRef]
- De Fombelle, A.; Frumholtz, P.; Poillion, D.; Drogoul, C.; Phillipeau, C.; Jacotot, E.; Julliand, V. Effect of the botanical origin of starch on its prececal digestibility measured with the mobile bag technique. Proc. Equine Nutr. Physiol. Soc. 2001, 82, 153–155. [Google Scholar]
- Liu, B.; Cui, Y.; Ali, Q.; Zhu, X.; Li, D.; Ma, S.; Wang, Z.; Wang, C.; Shi, Y. Gut microbiota modulate rabbit meat quality in response to dietary fiber. Front. Nutr. 2022, 9, 849429. [Google Scholar] [CrossRef] [PubMed]
- Vermorel, M.; Martin-Rosset, W. Concepts, scientific bases, structure and validation of the French horse net energy system (UFC). Livest. Prod. Sci. 1997, 47, 261–275. [Google Scholar] [CrossRef]
- Garber, A.; Hastie, P.; Murray, J.-A. Factors influencing equine gut microbiota: Current knowledge. J. Equine Vet. Sci. 2020, 88, 102943. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Barcenilla, A.; Pryde, S.E.; Martin, J.C.; Duncan, S.H.; Stewart, C.S.; Henderson, C.; Flint, H.J. Phylogenetic relationships of butyrate-producing bacteria from the human gut. Appl. Environ. Microbiol. 2000, 66, 1654–1661. [Google Scholar] [CrossRef]
- Wambacq, W.; Van Doorn, D.; Rovers-Paap, P.; Ducatelle, R.; Vlaminck, L.; Lourenco, M.; Hesta, M. Dietary supplementation of micro-encapsulated sodium butyrate in healthy horses: Effect on gut histology and immunohistochemistry parameters. BMC Vet. Res. 2020, 16, 1–9. [Google Scholar] [CrossRef]
- Rombeau, J.L.; Kripke, S.A. Metabolic and Intestinal Effects of Short-Chain Fatty Acids. J. Parenter. Enter. Nutr. 1990, 14, 181S–185S. [Google Scholar] [CrossRef]
- Grimm, P.; Philippeau, C.; Julliand, V. Faecal parameters as biomarkers of the equine hindgut microbial ecosystem under dietary change. Animal 2017, 11, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Dishaw, L.J.; Cannon, J.P.; Litman, G.W.; Parker, W. Immune-directed support of rich microbial communities in the gut has ancient roots. Dev. Comp. Immunol. 2014, 47, 36–51. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Bou, G.; Su, S.; Xing, J.; Qu, H.; Zhang, X.; Wang, X.; Zhao, Y.; Dugarjaviin, M. Microbial diversity within the digestive tract contents of Dezhou donkeys. PLoS ONE 2019, 14, e0226186. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, J.; Dang, W.; Irwin, D.M.; Wang, Z.; Zhang, S. Unveiling the biogeography and potential functions of the intestinal digesta-and mucosa-associated microbiome of donkeys. Front. Microbiol. 2020, 11, 596882. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Zhao, Y.; Liu, Z.; Liu, G.; Du, M.; Wu, J.; Bai, D.; Li, B.; Bou, G.; Zhang, X. Characterization and comparison of the bacterial microbiota in different gastrointestinal tract compartments of Mongolian horses. Microbiologyopen 2020, 9, 1085–1101. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, B.; Bai, D.; Huang, J.; Shiraigo, W.; Yang, L.; Zhao, Q.; Ren, X.; Wu, J.; Bao, W. Comparison of fecal microbiota of Mongolian and Thoroughbred Horses by high-throughput sequencing of the V4 Region of the 16S rRNA gene. Asian-Australas. J. Anim. Sci. 2016, 29, 1345. [Google Scholar] [CrossRef]
- Wunderlich, G.; Bull, M.; Ross, T.; Rose, M.; Chapman, B. Understanding the microbial fibre degrading communities & processes in the equine gut. Anim. Microbiome 2023, 5, 1–17. [Google Scholar]
- Liu, L.; Huang, W.C.; Liu, Y.; Li, M. Corrigendum to “Diversity of cellulolytic microorganisms and microbial cellulases”. Int. Biodeterior. Biodegrad. 2021, 165, 105325. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, C.; Wang, Y.; Du, M.; Zhang, G.; Lee, Y. Dietary Energy Level Impacts the performance of donkeys by manipulating the gut microbiome and metabolome. Front. Vet. Sci. 2021, 8, 694357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Huang, B.; Gao, X.; Shi, X.; Wang, X.; Wang, T.; Wang, Y.; Liu, G.; Wang, C. Dynamic changes in fecal microbiota in donkey foals during weaning: From pre-weaning to post-weaning. Front. Microbiol. 2023, 14, 1105330. [Google Scholar] [CrossRef]
- Liu, G.Q.; Gerelchimeg; Zhang, X.Z.; Xing, J.Y.; Qu, H.L.; Wang, T.; Su, S.F.; Liu, M.L.; Zhao, Y.P. Effects of Feeding Mode on Growth Performance, Nutrient Digestibility and Cecal Microbial Diversity of Dezhou Donkeys. Chin. J. Anim. Nutr. 2020, 32, 706–714. [Google Scholar]
- Harlow, B.E.; Lawrence, L.M.; Hayes, S.H.; Crum, A.; Flythe, M.D. Effect of Dietary Starch Source and Concentration on Equine Fecal Microbiota. PLoS ONE 2016, 11, e0154037. [Google Scholar] [CrossRef] [PubMed]
- Mshelia, E.S.; Adamu, L.; Wakil, Y.; Turaki, U.A.; Gulani, I.A.; Musa, J. The association between gut microbiome, sex, age and body condition scores of horses in Maiduguri and its environs. Microb. Pathog. 2018, 118, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Morrison, P.K.; Newbold, C.J.; Jones, E.; Worgan, H.J.; Grove-White, D.H.; Dugdale, A.H.; Barfoot, C.; Harris, P.A.; Argo, C.M. Effect of age and the individual on the gastrointestinal bacteriome of ponies fed a high-starch diet. PLoS ONE 2020, 15, e0232689. [Google Scholar] [CrossRef] [PubMed]
- Theelen, M.J.; Luiken, R.E.; Wagenaar, J.A.; Sloet van Oldruitenborgh-Oosterbaan, M.M.; Rossen, J.W.; Zomer, A.L. The equine faecal microbiota of healthy horses and ponies in the Netherlands: Impact of host and environmental factors. Animals 2021, 11, 1762. [Google Scholar] [CrossRef]
- Zeng, B.; Han, S.; Wang, P.; Wen, B.; Jian, W.; Guo, W.; Yu, Z.; Du, D.; Fu, X.; Kong, F.; et al. The bacterial communities associated with fecal types and body weight of rex rabbits. Sci. Rep. 2015, 5, 9342. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Items | Content |
|---|---|
| Ingredients | |
| Corn grain (GB2 grade) | 35.43 |
| Wheat flour (NY/T1 grade) | 20.00 |
| Corn germ meal | 25.00 |
| Cornstarch residue | 10.00 |
| Mineral meal | 1.97 |
| Calcium hydrogen phosphate | 1.14 |
| Salt | 0.60 |
| Lysine | 0.85 |
| Threonine | 0.10 |
| Flavoring agent | 0.02 |
| Magnesium sulfate | 0.20 |
| Microminerals for donkey 1 | 0.20 |
| Donkey vitamin 2 | 0.15 |
| Soybean meal (GB1 grade) | 4.34 |
| Total | 100 |
| Nutrient levels 3 | |
| Crude protein | 16.03 |
| Crude fiber | 13.28 |
| Ether extract | 2.08 |
| Acid detergent fiber | 23.08 |
| Neutral detergent fiber | 16.93 |
| Nutrient Levels | Content |
|---|---|
| Crude protein | 6.62 |
| Crude fiber | 67.24 |
| Ether extract | 0.39 |
| Acid detergent fiber | 79.51 |
| Neutral detergent fiber | 82.02 |
| Indicators | C1 | C2 | C3 | p Value |
|---|---|---|---|---|
| Initial BW, kg | 117.00 ± 5.56 | 117.20 ± 4.44 | 117.20 ± 5.30 | 0.999 |
| Final BW, kg | 127.60 ± 5.92 | 131.50 ± 5.13 | 132.90 ± 6.33 | 0.803 |
| ADFI, kg/day | 2.69 ± 0.02 b | 2.79 ± 0.02 a | 2.76 ± 0.02 a | 0.022 |
| ADG, kg/day | 0.30 ± 0.02 b | 0.41 ± 0.04 a | 0.45 ± 0.03 a | 0.003 |
| F/G | 9.30 ± 0.69 a | 7.28 ± 0.59 b | 6.33 ± 0.34 b | 0.003 |
| Indicators | C1 | C2 | C3 | p Value |
|---|---|---|---|---|
| CP | 73.05 ± 0.008 b | 76.82 ± 0.009 a | 77.72 ± 0.005 a | 0.002 |
| EE | 35.40 ± 0.019 b | 50.41 ± 0.019 a | 46.63 ± 0.026 a | 0.002 |
| CF | 63.77 ± 0.539 | 63.07 ± 0.555 | 64.00 ± 0.947 | 0.637 |
| NDF | 67.09 ± 0.827 | 66.33 ± 0.858 | 67.83 ± 0.675 | 0.434 |
| ADF | 59.03 ± 0.490 | 58.58 ± 0.275 | 60.36 ± 0.712 | 0.081 |
| Indicators | C1 | C2 | C3 | p Value |
|---|---|---|---|---|
| Acetic acid | 220.97 ± 15.48 a | 153.74 ± 16.38 b | 133.94 ± 20.76 b | 0.017 |
| Propionic acid | 43.65 ± 6.85 | 38.46 ± 8.50 | 36.20 ± 5.30 | 0.750 |
| Butyric acid | 12.45 ± 3.43 | 10.14 ± 2.33 | 9.80 ± 2.29 | 0.764 |
| Isobutyric acid | 8.36 ± 0.90 a | 4.90 ± 0.18 b | 4.11 ± 0.35 b | 0.001 |
| Valeric acid | 4.78 ± 0.50 a | 2.48 ± 0.55 b | 1.85 ± 0.23 b | 0.003 |
| Isovaleric acid | 5.85 ± 0.78 a | 3.93 ± 0.50 b | 2.73 ± 0.44 b | 0.014 |
| Caproic acid | 0.19 ± 0.03 a | 0.10 ± 0.04 ab | 0.05 ± 0.01 b | 0.032 |
| Total VFA | 296.26 ± 19.64 a | 213.74 ± 26.04 b | 188.67 ± 23.80 b | 0.024 |
| Characteristic | C1 | C2 | C3 | SEM | p Value |
|---|---|---|---|---|---|
| Sequences | 80,677.00 | 79,425.00 | 82,771.75 | 624.090 | 0.071 |
| OTUs | 1614.25 a | 1245.00 b | 1283.50 b | 67.530 | 0.028 |
| Chao1 index | 1617.40 a | 1246.41 b | 1286.06 b | 67.520 | 0.027 |
| Shannon index | 9.52 a | 8.51 b | 8.88 ab | 0.180 | 0.040 |
| Simpson index | 1.00 | 0.99 | 0.99 | 0.002 | 0.305 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, L.; Xing, J.; Qi, X.; Lu, T.; Jin, Y.; Akhtar, M.F.; Li, L.; Liu, G. Effects of Concentrate Feeding Sequence on Growth Performance, Nutrient Digestibility, VFA Production, and Fecal Microbiota of Weaned Donkeys. Animals 2023, 13, 2893. https://doi.org/10.3390/ani13182893
Xie L, Xing J, Qi X, Lu T, Jin Y, Akhtar MF, Li L, Liu G. Effects of Concentrate Feeding Sequence on Growth Performance, Nutrient Digestibility, VFA Production, and Fecal Microbiota of Weaned Donkeys. Animals. 2023; 13(18):2893. https://doi.org/10.3390/ani13182893
Chicago/Turabian StyleXie, Lan, Jingya Xing, Xingzhen Qi, Ting Lu, Yaqian Jin, Muhammad Faheem Akhtar, Lanjie Li, and Guiqin Liu. 2023. "Effects of Concentrate Feeding Sequence on Growth Performance, Nutrient Digestibility, VFA Production, and Fecal Microbiota of Weaned Donkeys" Animals 13, no. 18: 2893. https://doi.org/10.3390/ani13182893
APA StyleXie, L., Xing, J., Qi, X., Lu, T., Jin, Y., Akhtar, M. F., Li, L., & Liu, G. (2023). Effects of Concentrate Feeding Sequence on Growth Performance, Nutrient Digestibility, VFA Production, and Fecal Microbiota of Weaned Donkeys. Animals, 13(18), 2893. https://doi.org/10.3390/ani13182893