


The Benefits of Increased Space and Habitat Complexity for the Welfare of Zoo-Housed King Penguins (Aptenodytes patagonicus)

 ,
,  ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
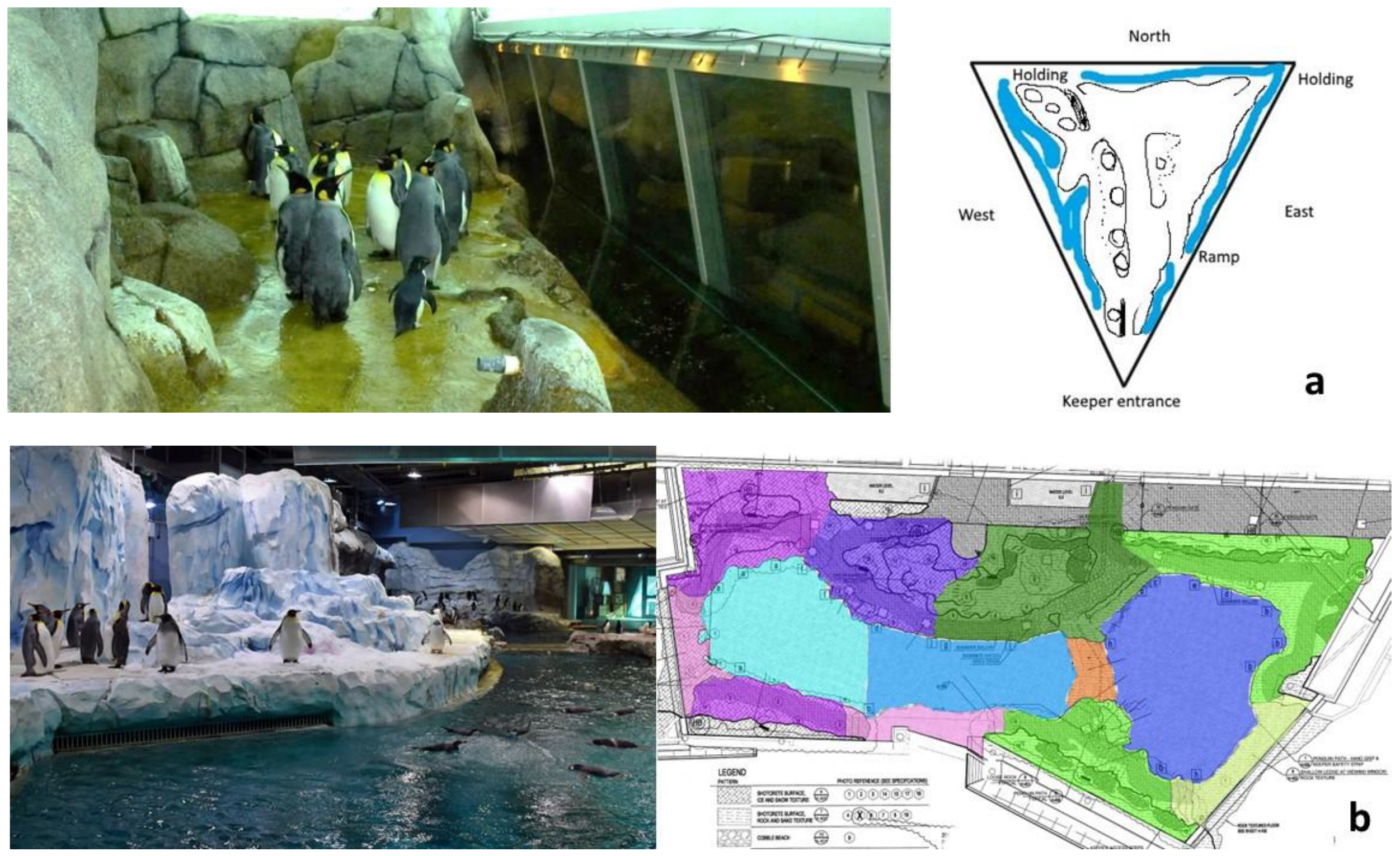
2.1. Study and Habitat (Exhibit) Design
2.2. Subjects and Husbandry
2.3. Observational Data Collection
2.4. Time–Depth Recorders
2.5. Data Analysis
3. Results
3.1. Time–Depth Recorder Data
3.2. Behavioral Comparison across Study Phases
3.3. Aquatic Behaviors
3.4. Other Solitary Behaviors
3.5. Social Behaviors
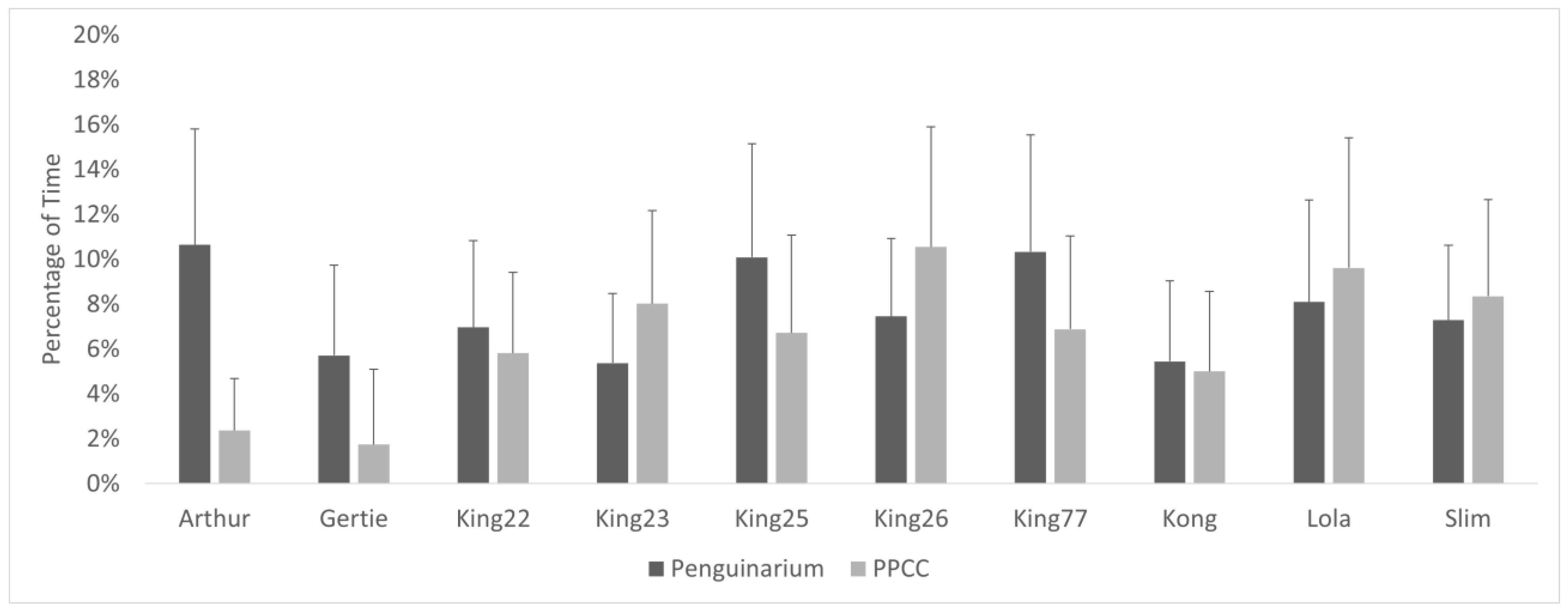
3.6. Social Proximity
4. Discussion
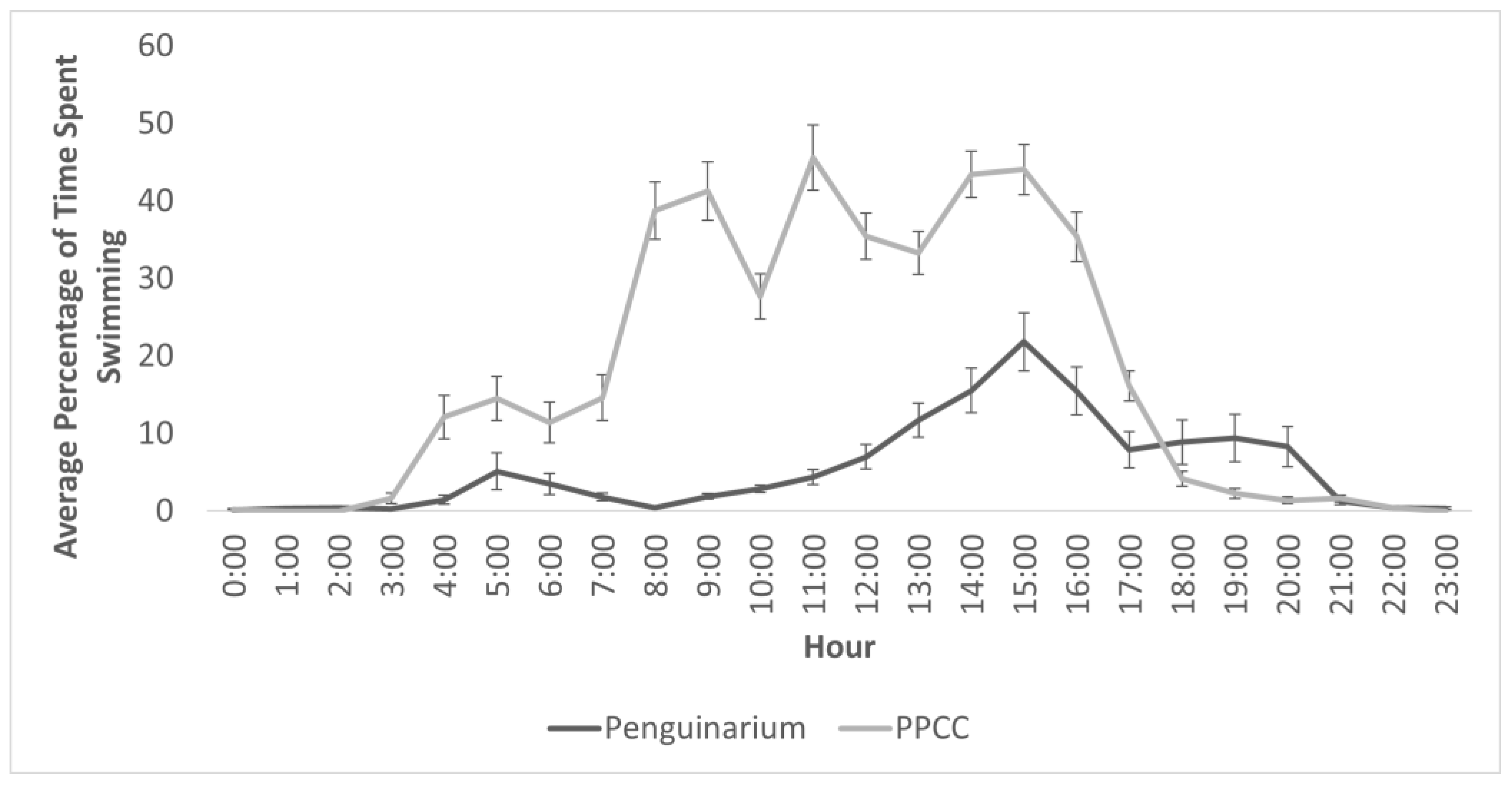
4.1. Changes in Aquatic Behaviors
4.2. Changes in Other Solitary Behaviors
4.3. Changes in Social Dynamics
5. Conclusions: Implications for Habitat Design and Penguin Welfare
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kelling, A.S.; Gaalema, D.E. Postoccupancy evaluations in zoological settings. Zoo Biol. 2011, 30, 597–610. [Google Scholar] [CrossRef]
- Coe, J.C. Naturalizing habitats for captive primates. Zoo Biol. 1989, 8, 117–125. [Google Scholar] [CrossRef]
- Maple, T.L.; Finlay, T.W. Post-occupancy evaluation in the zoo. Appl. Anim. Behav. Sci. 1987, 18, 5–18. [Google Scholar] [CrossRef]
- Coe, J.C. Steering the ark toward Eden: Design for animal well-being. J. Am. Vet. Med. Assoc. 2003, 223, 977–980. [Google Scholar] [CrossRef] [PubMed]
- Fàbregas, M.C.; Guillén-Salazar, F.; Garcés-Narro, C. Do naturalistic enclosures provide suitable environments for zoo animals? Zoo Biol. 2012, 31, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 Five Domains Model: Including Human–Animal Interactions in Assessments of Animal Welfare. Animals 2020, 10, 1870. [Google Scholar] [CrossRef]
- Lawrence, K.; Sherwen, S.L.; Larsen, H. Natural Habitat Design for Zoo-Housed Elasmobranch and Teleost Fish Species Improves Behavioural Repertoire and Space Use in a Visitor Facing Exhibit. Animals 2021, 11, 2979. [Google Scholar] [CrossRef]
- Fieschi-Méric, L.; Ellis, C.; Servini, F.; Tapley, B.; Michaels, C.J. An Improvement in Enclosure Design Can Positively Impact Welfare, Reduce Aggressiveness and Stabilise Hierarchy in Captive Galapagos Giant Tortoises. J. Zool. Bot. Gard. 2022, 3, 499–512. [Google Scholar] [CrossRef]
- Brooking, Z.A.; Price, D.J. The effect of human presence on the behavioural and distributional patterns of two species of captive penguins, African (Spheniscus demersus) and gentoo (Pygoscelis papua), on being transferred into their new enclosure at Living Coasts, Devon. In Proceedings of the Sixth Annual Symposium on Zoo Research, Edinburgh, UK, 8–9 July 2004; pp. 170–179. [Google Scholar]
- Chiew, S.J.; Butler, L.K.; Sherwen, L.S.; Coleman, J.G.; Fanson, V.K.; Hemsworth, H.P. Effects of Regulating Visitor Viewing Proximity and the Intensity of Visitor Behaviour on Little Penguin (Eudyptula minor) Behaviour and Welfare. Animals 2019, 9, 285. [Google Scholar] [CrossRef]
- Chiew, S.J.; Butler, K.L.; Sherwen, S.L.; Coleman, G.J.; Melfi, V.; Burns, A.; Hemsworth, P.H. Effect of Covering a Visitor Viewing Area Window on the Behaviour of Zoo-Housed Little Penguins (Eudyptula minor). Animals 2020, 10, 1224. [Google Scholar] [CrossRef]
- Edes, A.N.; Baskir, E.; Bauman, K.L.; Chandrasekharan, N.; Macek, M.; Tieber, A. Effects of crowd size, composition, and noise level on pool use in a mixed-species penguin colony. Anim. Behav. Cogn. 2021, 8, 507–520. [Google Scholar] [CrossRef]
- Hartell-DeNardo, J.; Kozlowski, C.; Baskir, E.; Macek, M.; Dorsey, C.; Powell, D.M. Behavior and adrenal physiology of Magellanic penguins (Spheniscus magellanicus) serving as ambassador animals. Zoo Biol. 2022, 42, 243–253. [Google Scholar] [CrossRef]
- Saiyed, T.S.; Hopper, M.L.; Cronin, A.K. Evaluating the Behavior and Temperament of African Penguins in a Non-Contact Animal Encounter Program. Animals 2019, 9, 326. [Google Scholar] [CrossRef] [PubMed]
- Sherwen, S.L.; Magrath, M.J.L.; Butler, K.L.; Hemsworth, P.H. Little penguins, Eudyptula minor, show increased avoidance, aggression and vigilance in response to zoo visitors. Appl. Anim. Behav. Sci. 2015, 168, 71–76. [Google Scholar] [CrossRef]
- Williams, E.; Carter, A.; Rendle, J.; Ward, S.J. Understanding impacts of zoo visitors: Quantifying behavioural changes of two popular zoo species during COVID-19 closures. Appl. Anim. Behav. Sci. 2021, 236, 105253. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.R.; Deere, N.J.; Little, H.A.; Snipp, R.; Goulder, J.; Mayer-Clarke, S. Husbandry and enclosure influences on penguin behavior and conservation breeding. Zoo Biol. 2016, 35, 385–397. [Google Scholar] [CrossRef]
- Reisfeld, L.; Moraes, K.; Spaulussi, L.; Cardoso, R.C.; Ippolito, L.; Gutierrez, R.; Silvatti, B.; Pizzutto, C.S. Behavioral Responses of Magellanic Penguins (Spheniscus magellanicus) (Foster) to Saltwater Versus Freshwater. Zoo Biol. 2013, 32, 575–577. [Google Scholar] [CrossRef]
- Larsson, A. Development and Evaluation of Environmental Enrichment for Captive Humboldt penguins; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2012. [Google Scholar]
- Collins, C.; Quirke, T.; Overy, L.; Flannery, K.; O’Riordan, R. The effect of the zoo setting on the behavioural diversity of captive gentoo penguins and the implications for their educational potential. J. Zoo Aquar. Res. 2016, 4, 85–90. [Google Scholar]
- Reisfeld, L.; Barbirato, M.; Ippolito, L.; Cardoso, R.C.; Nichi, M.; Sgai, M.; Pizzutto, C.S. Reducing bumblefoot lesions in a group of captive Magellanic penguins (Spheniscus magellanicus) with the use of environmental enrichment. Pesqui. Vet. Bras. 2013, 33, 791–795. [Google Scholar] [CrossRef]
- Blay, N.; Cote, I.M. Optimal conditions for breeding of captive Humboldt penguins (Spheniscus humboldti): A survey of British zoos. Zoo Biol. 2001, 20, 545–555. [Google Scholar] [CrossRef]
- Heintz, M.R.; Fuller, G.A.; Woodhouse, S.J.; Murray, A.; Allard, S.M. Case studies of the impacts of cataract surgery on crested penguin welfare. J. Appl. Anim. Welf. Sci. 2019, 22, 329–341. [Google Scholar] [CrossRef]
- Andrews, R.D.; Enstipp, M.R. Diving physiology of seabirds and marine mammals: Relevance, challenges and some solutions for field studies. Comp. Biochem. Physiol. Part A 2016, 202, 38–52. [Google Scholar] [CrossRef]
- Gauthier-Clerc, M.; Le Maho, Y.; Gendner, J.-P.; Handrich, Y. Moulting fast and time constraint for reproduction in the king penguin. Polar Biol. 2002, 25, 288–295. [Google Scholar] [CrossRef]
- Bost, C.A.; Georges, J.Y.; Guinet, C.; Cherel, Y.; Putz, K.; Charrassin, J.B.; Handrich, Y.; Zorn, T.; Lage, J.; LeMaho, Y. Foraging habitat and food intake of satellite-tracked king penguins during the austral summer at Crozet Archipelago. Mar. Ecol. Prog. Ser. 1997, 150, 21–33. [Google Scholar] [CrossRef]
- Charrassin, J.B.; Bost, C.A. Utilisation of the oceanic habitat by king penguins over the annual cycle. Mar. Ecol. Prog. Ser. 2001, 221, 285–297. [Google Scholar] [CrossRef]
- Jouventin, P.; Capdeville, D.; Cuenot-Chaillet, F.; Boiteau, C. Exploitation of pelagic resources by a non-flying seabird: Satellite tracking of the king penguin throughout the breeding cycle. Mar. Ecol. Prog. Ser. 1994, 106, 11–19. [Google Scholar] [CrossRef]
- Groscolas, R.; Cherel, Y. How to molt while fasting in the cold—The metabolic and hormonal adaptations of emperor and king penguins. Ornis Scand. 1992, 23, 328–334. [Google Scholar] [CrossRef]
- Van Den Hoff, J.; Copley, P. Aspects of the breeding cycle of king penguins Aptenodytes patagonicus at Heard Island. Mar. Ornithol. 1993, 21, 49–55. [Google Scholar]
- CôTé, S.D. Aggressiveness in king penguins in relation to reproductive status and territory location. Anim. Behav. 2000, 59, 813–821. [Google Scholar] [CrossRef]
- Waterfall, K.; Kottyan, J.; Sirpenski, G.; Huetnner, S.; Schneider, T. AZA Penguin Regional Collection Plan 2020–2025; Association of Zoos and Aquariums: Silver Spring, MD, USA, 2020. [Google Scholar]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 48, 227–265. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.R.; Niemann, T.; Wark, J.D.; Heintz, M.R.; Horrigan, A.; Cronin, K.A.; Shender, M.A.; Gillespie, K. ZooMonitor, Version 3; Lincoln Park Zoo: Chicago, IL, USA, 2016. [Google Scholar]
- Fuller, G.; Heintz, M.R.; Allard, S. Validation and welfare assessment of flipper-mounted time-depth recorders for monitoring penguins in zoos and aquariums. Appl. Anim. Behav. Sci. 2019, 212, 114–122. [Google Scholar] [CrossRef]
- Berry, K.J.; Mielke, P.W. Exact and Monte Carlo Resampling Procedures for the Wilcoxon-Mann-Whitney and Kruskal-Wallis Tests. Percept. Mot. Ski. 2000, 91, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Colgrave, N.J.; Engel, J.; Plowman, A.B. Randomization tests. In Zoo Research Guidelines: Statistics for Typical Zoo Datasets; Plowman, A.B., Ed.; BIAZA: London, UK, 2006; pp. 7–16. [Google Scholar]
- Plowman, A.B. BIAZA statistics guidelines: Toward a common application of statistical tests for zoo research. Zoo Biol. 2008, 27, 226–233. [Google Scholar] [CrossRef]
- Clarke, A.G. Factors affecting pool use by captive Humboldt penguins (Spheniscus humboldti). In Proceedings of the Fifth Annual Symposium on Zoo Research, Marwell Zoological Park, Winchester, UK, 7–8 July 2003; pp. 190–204. [Google Scholar]
- Fernandez, E.J.; Myers, M.; Hawkes, N.C. The Effects of Live Feeding on Swimming Activity and Exhibit Use in Zoo Humboldt Penguins (Spheniscus humboldti). J. Zool. Bot. Gard. 2021, 2, 88–100. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Kinley, R.C.; Timberlake, W. Training penguins to interact with enrichment devices for lasting effects. Zoo Biol. 2019, 38, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Kalafut, K.L.; Kinley, R. Using radio frequency identification for behavioral monitoring in little blue penguins. J. Appl. Anim. Welf. Sci. 2020, 23, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Brando, S.; Buchanan-Smith, H.M. The 24/7 approach to promoting optimal welfare for captive wild animals. Behav. Process. 2017, 156, 83–95. [Google Scholar] [CrossRef]
- Challet, E.; Bost, C.A.; Handrich, Y.; Gendner, J.P.; Lemaho, Y. Behavioral time budget of breeding king penguins (Aptenodytes patagonica). J. Zool. 1994, 233, 669–681. [Google Scholar] [CrossRef]
- Cannell, B.L.; Cullen, J.M. The foraging behaviour of little penguins Eudyptula minor at different light levels. Ibis 1998, 140, 467–471. [Google Scholar] [CrossRef]
- Foerder, P.; Chodorow, M.; Moore, D.E. Behavioural synchrony in two species of communally housed captive penguins. Behaviour 2013, 150, 1357–1374. [Google Scholar] [CrossRef]
- Galante, J.; Margulis, S.W. Comparing Data Collection Tools for Zoo Management Decision-Making: A Case Study Examining Behavioral Measures of Humboldt Penguin Bond Strength. Animals 2022, 12, 3031. [Google Scholar] [CrossRef] [PubMed]
- Koene, P.; Vanderschoot, K.; Nijboer, J. Environmental change and welfare of captive king penguins (Aptenodytes patagonicus). J. Appl. Anim. Welf. Sci. 2013, 16, 385–386. [Google Scholar] [CrossRef]
- Le Vaillant, M.; Ropert-Coudert, Y.; Le Maho, Y.; Le Bohec, C. Individual parameters shape foraging activity in breeding king penguins. Behav. Ecol. 2016, 27, 352–362. [Google Scholar] [CrossRef]
- Merritt, K.; King, N.E. Behavioral sex differences and activity patterns of captive Humboldt penguins (Spheniscus humboldti). Zoo Biol. 1987, 6, 129–138. [Google Scholar] [CrossRef]
- Little, K.A.; Sommer, V. Change of enclosure in langur monkeys: Implications for the evaluation of environmental enrichment. Zoo Biol. 2002, 21, 549–559. [Google Scholar] [CrossRef]
- Quintavalle Pastorino, G.; Preziosi, R.; Faustini, M.; Curone, G.; Albertini, M.; Nicoll, D.; Moffat, L.; Pizzi, R.; Mazzola, S. Comparative Personality Traits Assessment of Three Species of Communally Housed Captive Penguins. Animals 2019, 9, 376. [Google Scholar] [CrossRef]
- Scott, N.L.; LaDue, C.A. The behavioral effects of exhibit size versus complexity in African elephants: A potential solution for smaller spaces. Zoo Biol. 2019, 38, 448–457. [Google Scholar] [CrossRef]
- Dawkins, M.S. The Science of Animal Welfare: Understanding What Animals Want; Oxford University Press: Oxford, UK, 2021. [Google Scholar]
- Ross; Schapiro, S.J.; Hau, J.; Lukas, K.E. Space use as an indicator of enclosure appropriateness: A novel measure of captive animal welfare. Appl. Anim. Behav. Sci. 2009, 121, 42–50. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Phase | Sub-Period | Data Collected: Hours (Number of Observations) | Habitat | Dates | Description |
|---|---|---|---|---|---|
| Phase 1 2015–2017 | Penguinarium I | 223.3 (1476) | Penguinarium | 2 March 2015–29 February 2016 | Regular data collection prior to the first move |
| 8 June 2015–24 July 2015 | TDR data collection during the breeding season | ||||
| 4 December 2015–19 January 2016 | TDR data collection during the non-breeding season | ||||
| Interlude | 0 (0) | Penguinarium | 1 March 2016–12 May 2016 | King penguins not observed during their molt in the Penguinarium | |
| PPCC II | 321.4 (2069) | PPCC | 13 May 2016–31 July 2017 | Regular data collection after their first move to the PPCC | |
| 6 December 2016–18 January 2017 | TDR data collection during the non-breeding season | ||||
| 8 June 2017–23 July 2017 | TDR data collection during the breeding season | ||||
| No data collection | 0 (0) | PPCC | 1 August 2017–4 August 2019 | Penguins not observed | |
| Phase 2 2019–2022 | PPCC III | 8.4 (103) | PPCC | 5 August 2019–6 September 2019 | Regular data collection |
| Penguinarium IV | 88.6 (1078) | Penguinarium | 10 September 2019–28 June 2021 | Penguins not viewable by the public. Note that this period and the one below overlap, as the king penguins were moved gradually to the PPCC as they completed their annual molt. | |
| PPCC V | 53.1 (642) | PPCC | 10 June 2021–29 April 2022 | Penguins not viewable by the public until building re-opening on 14 February 2022 | |
| Name | Sex | Age at Observation Onset | Study Periods |
|---|---|---|---|
| Arthur | M | 2.62 | III–V |
| Gertie | F | 3.68 | III–V |
| King 22 * | M | 13.49 | I–V (all) |
| King 23 | M | 13.48 | I and II |
| King 25 * | F | 14.01 | I–V (all) |
| King 26 | F | 4.92 | I and II |
| King 77 * | F | 26.92 | I–V (all) |
| Kong * | M | 19.58 | I–V (all) |
| Lola | F | 26.97 | I and II |
| Slim | M | 21.60 | I and II |
| Behavior | Operational Definition |
|---|---|
| Activity (Intervals/Scans) | |
| Allopreen * | Rubbing the head, bill, or flipper on another individual, or “mouthing” another individual |
| Receive allopreen * | Receiving preening from another penguin |
| Contact aggression * | Pecking, wing blows, or gripping and twisting with bill directed at another penguin |
| Receive contact aggression * | Receiving contact aggression from another penguin |
| Noncontact aggression * | Lunging at another penguin or attempting to peck one without actually making contact, or pointing the head toward and aggressively vocalizing at another penguin |
| Receive noncontact aggression * | The focal penguin is the recipient of lunging, attempted pecks, or aggressive vocal displays from another penguin. |
| Display | Performing an ecstatic or mutual display |
| Mate * | Any breeding behavior between penguins, including dorso-ventral mounting and cloacal rubbing; also includes rapidly moving flippers (“vibrating”) on the body of another |
| Chick-directed | Any behavior directed toward a chick, including regurgitating food, preening chicks, and restraining chicks from moving |
| Keeper-directed | Following, pointing, leaning towards (>45°) and vocalizing at or otherwise interacting with a human (keeper) in the exhibit; includes breeding behaviors directed towards a keeper, e.g., flipper vibrating |
| Investigate | Pecking at or manipulating rockwork or objects on land or in the pool, including enrichment items, ice cubes, whale bones on land, bubbles and floating debris in the pool, kelp, underwater decorations, or shaved ice/snow |
| Bathe | Preening or adjustment movements in water, including tail wags and shaking movements; can occur in the pool or using water features (fountains, etc.) on land |
| Surface swim | Penguin uses the water to propel itself at a depth less than 1 m, including porpoising or floating; does not include walking on submerged land |
| Dive | Swimming underwater at depths greater than 1 m |
| Walk | Moving at any pace by lifting and setting down each foot in turn; includes using feet on the ground to propel the body across submerged land |
| Vocalize | Producing sounds using the mouth; bill is open |
| Feed | Ingesting or chewing food or drinking water; includes being fed by keeper and manipulating food in the water (even if ingestion is not observed) |
| Shiver | Slightly shaking the whole body; less intense than adjustment movements and involves the whole body instead of just the flippers (flipper vibrating is a component of mating behavior) |
| Preen | Includes contact between the bill and feathers, rubbing the head over another body part, wing rub on head or neck, or scratching with feet |
| Adjust | Shaking and stretching movements, including head shake, head bob, body shake, tail wag, or rapid wing flap; may perform these behaviors in the water as part of a bathing bout |
| Stand | Resting in an upright position; may be moving the head or eyes to observe the surroundings, or head may be tucked under the wing while the penguin is sleeping |
| Lay | Resting on the belly or in any position other than standing |
| Other | Any other behavior not detailed elsewhere involving vigorous movement; may include shaking the head in the nest (looks like a partial ecstatic display) or making biting movements in the air |
| Not visible | Behavior cannot be definitively determined and/or the observer has no idea where the penguin is located |
| Additional All-Occurrence Behaviors | |
| Ecstatic display | A penguin is standing up on its toes, flippers extended vertically, head and bill also held up vertically, and vocalizing; in king penguins, the display involves tilting the head back, trumpeting, and then bowing; may also involve rapid bill movements (“clacking”) in kings |
| Mutual display | Two penguins in proximity to one another simultaneously performing an ecstatic display, or two penguins (usually kings) rubbing their beaks together or “kissing” |
| Porpoise | Leaping in and out of the water in short, shallow arcs; must include two leaps in quick succession to count as a porpoising bout |
| Toboggan | Horizontal movement on land by sliding on the belly, often using the feet or flippers to propel the body forward |
| Land bathe | Preening or adjustment movements including tail wags and shaking movements occurring in land-based water features such as the splash zone, waterfalls, or hose jets |
| Water bathe | Preening or adjustment movements including tail wags and shaking movements occurring when the penguin is in the pool or on submerged land |
| Comparison | Average ± SE | Statistic (S) | Z | Pr > |Z| | Pr > |S-Mean| |
|---|---|---|---|---|---|
| Percentage of time between habitats * | Peng.: 5.44 ± 0.97% PPCC: 17.71 ± 1.28% | 49.00 | −1.9954 | 0.0460 | 0.0494 |
| Surface by habitat ** | Peng.: 69.31 ± 3.12% PPCC: 40.06 ± 3.00% | 166.00 | −2.9355 | 0.0032 | 0.0024 |
| 0–2 m by habitat | Peng.: 30.69 ± 3.12% PPCC: 24.92 ± 2.95% | 220.00 | −0.7934 | 0.4276 | 0.4373 |
| 2–4 m by habitat *** | Peng.: -- PPCC: 27.59 ± 0.49% | 344.00 | 4.5829 | <0.0001 | <0.0001 |
| 4–6 m by habitat *** | Peng.: -- PPCC: 7.37 ± 0.07% | 344.00 | 4.5829 | <0.0001 | <0.0001 |
| 6–8 m by habitat *** | Peng.: -- PPCC: 0.06 ± 0.00% | 320.00 | 3.8083 | 0.0001 | <0.0001 |
| Average depth between habitats *** | Peng.: 0.06 ± 0.01 m PPCC: 1.43 ± 0.03 m | 332.00 | 3.6495 | 0.0005 | 0.0001 |
| Behavior | Average ± SE | Statistic (S) | Z | Pr > |Z| | Pr > |S-Mean| |
|---|---|---|---|---|---|
| Interval Behaviors | |||||
| Swim Phase 1 *** | Peng.: 7.63 ± 3.13% PPCC: 24.65 ± 5.19% | 2,333,269 | −13.4543 | <0.0001 | <0.0001 |
| Swim Phase 2 *** | Peng.: 0.89 ± 1.29% PPCC: 12.03 ± 5.21% | 728,818 | 10.3590 | <0.0001 | <0.0001 |
| Feed Phase 1 *** | Peng.: 0.16 ± 0.19% PPCC: 0.29 ± 0.32% | 2,333,269 | −13.4543 | <0.0001 | <0.0001 |
| Feed Phase 2 *** | Peng.: 0.22 ± 0.36% PPCC: 0.38 ± 0.52% | 728,818 | 10.3590 | <0.0001 | <0.0001 |
| Lay Phase 1*** | Peng.: 2.36 ± 1.73% PPCC.: 3.77 ± 2.01% | 2,561,076 | −4.6298 | <0.0001 | <0.0001 |
| Lay Phase 2 ** | Peng.: 3.92 ± 3.06% PPCC: 6.56 ± 3.92% | 691,578 | 2.6400 | 0.0042 | 0.0076 |
| Walk Phase 1 *** | Peng.: 18.63 ± 3.13% PPCC: 6.88 ± 1.56% | 3,014,571 | 14.8676 | <0.0001 | <0.0001 |
| Walk Phase 2 *** | Peng.: 19.05 ± 4.47% PPCC: 5.84 ± 2.45% | 564,435 | −12.3639 | <0.0001 | <0.0001 |
| All-Occurrence Behaviors | |||||
| Displays Phase 1 *** | Peng.: 0.39 ± 0.46 PPCC: 0.70 ± 0.55 | 2,582,905 | −3.6770 | 0.0002 | 0.0002 |
| Displays Phase 2 * | Peng.: 0.88 ± 0.88 PPCC: 0.71 ± 1.05 | 673,110.5 | −2.0448 | 0.0409 | 0.0494 |
| Bathing Phase 1 *** | Peng.: 0.00 ± 0.00 PPCC: 0.85 ± 0.62 | 2,532,078 | −9.2052 | <0.0001 | <0.0001 |
| Bathing Phase 2 *** | Peng.: 0.28 ± 0.59 PPCC: 1.44 ± 1.26 | 696,364 | 5.4141 | <0.0001 | <0.0001 |
| Agonism—given Phase 1 *** | Peng.: 3.59 ± 1.23 PPCC: 1.37 ± 0.63 | 2,784,090 | 8.6045 | <0.0001 | <0.0001 |
| Agonism—given Phase 2 * | Peng.: 4.95 ± 1.68 PPCC: 3.61 ± 1.33 | 666,041 | −2.1660 | 0.0303 | 0.0309 |
| Agonism—received Phase 1 *** | Peng.: 3.01 ± 1.00 PPCC: 1.05 ± 0.53 | 2,805,129 | 10.2069 | <0.0001 | <0.0001 |
| Agonism—received Phase 2 * | Peng.: 2.08 ± 1.30 PPCC: 1.51 ± 1.12 | 667,904.5 | −2.1100 | 0.0175 | 0.0335 |
| Agonism total Phase 1*** | Peng.: 6.60 ± 1.95 PPCC: 2.42 ± 1.03 | 2,875,473 | 11.6583 | <0.0001 | <0.0001 |
| Agonism total Phase 2 ** | Peng.: 4.95 ± 2.56 PPCC: 3.61 ± 2.19 | 658,138 | −3.0385 | 0.0024 | 0.0022 |
| Allopreening Phase 1 ** | Peng.: 0.22 ± 0.26 PPCC: 0.43 ± 0.36 | 2,593,365 | −2.8504 | 0.0044 | 0.0041 |
| Allopreening Phase 2 | Peng.: 0.28 ± 0.48 PPCC: 0.25 ± 0.38 | 680,505 | 0.4882 | 0.6254 | 0.6650 |
| Proximity Measures | |||||
| Alone Phase 1 * | Peng.: 46.72 ± 4.40% PPCC: 44.03 ± 4.67% | 2,681,066 | 2.1504 | 0.0315 | 0.0307 |
| Alone Phase 2 | Peng.: 34.77 ± 6.01% PPCC: 38.57 ± 6.96% | 687,145 | 0.7278 | 0.4668 | 0.4676 |
| Proximate to another king Phase 1 | Peng.: 45.70 ± 4.36% PPCC: 46.94 ± 4.66% | 2,572,424 | −1.4941 | 0.1352 | 0.1361 |
| Proximate to another king Phase 2 | Peng.: 56.66 ± 6.19% PPCC: 58.92 ± 7.05% | 690,250.5 | 1.0125 | 0.3113 | 0.3132 |
| Proximate to individual of another species Phase 1 | Peng.: 7.44 ± 1.64% PPCC: 8.35 ± 1.96% | 2,625,542 | 0.3365 | 0.7365 | 0.7344 |
| Proximate to individual of another species Phase 2 *** | Peng.: 8.48 ± 2.84% PPCC: 2.51 ± 1.72% | 604,540 | −9.9842 | <0.0001 | <0.0001 |
| Behavior | Average ± SE | Statistic (S) | Z | Pr > |Z| | Pr > |S-Mean| |
|---|---|---|---|---|---|
| Interval Behaviors | |||||
| Swim *** | Peng.: 4.78 ± 2.00% PPCC: 21.31 ± 3.95% | 6,205,560 | −18.1117 | <0.0001 | <0.0001 |
| Feed | Peng.: 0.19 ± 0.18% PPCC: 0.31 ± 0.27% | 6,835,768 | −1.7506 | 0.0800 | 0.0805 |
| Lay *** | Peng.: 3.02 ± 1.59% PPCC: 4.50 ± 1.84% | 6,738,055 | −5.1233 | <0.0001 | <0.0001 |
| Walk *** | Peng.: 18.81 ± 2.58% PPCC: 6.60 ± 1.31% | 7,702,521 | 17.0888 | <0.0001 | <0.0001 |
| All-Occurrence Behaviors | |||||
| Displays * | Peng.: 0.60 ± 0.44 PPCC: 0.70 ± 0.50 | 6,820,722 | −2.0932 | 0.0363 | 0.0397 |
| Bathing *** | Peng.: 0.12 ± 0.23 PPCC: 1.01 ± 0.58 | 6,675,406 | −10.6635 | <0.0001 | <0.0001 |
| Agonism—given *** | Peng.: 3.29 ± 0.99 PPCC: 1.57 ± 0.60 | 7,115,008 | 7.3697 | <0.0001 | <0.0001 |
| Agonism—received *** | Peng.: 2.62 ± 0.79 PPCC: 1.17 ± 0.50 | 7,135,458 | 8.5281 | <0.0001 | <0.0001 |
| Agonism total *** | Peng.: 5.91 ± 1.55 PPCC: 2.74 ± 0.98 | 7,256,697 | 9.9999 | <0.0001 | <0.0001 |
| Allopreening ** | Peng.: 0.25 ± 0.24 PPCC: 0.39 ± 0.27 | 6,810,070 | −3.2308 | 0.0012 | 0.0010 |
| Proximity Measures | |||||
| Alone * | Peng.: 41.68 ± 3.60% PPCC: 42.59 ± 3.86% | 6,722,278 | −2.3989 | 0.0164 | 0.0166 |
| Proximate to another king *** | Peng.: 50.33 ± 3.62% PPCC: 50.11 ± 3.90% | 6,616,997 | −4.2561 | <0.0001 | <0.0001 |
| Proximate to individual of another species ** | Peng.: 7.88 ± 1.49% PPCC: 6.80 ± 1.47% | 6,985,366 | 2.8379 | 0.0023 | 0.0044 |
| Behavior | Effect | Estimate | S.E. | DF | T Value | Pr > |t| | Lower | Upper | |
|---|---|---|---|---|---|---|---|---|---|
| Interval Behaviors | |||||||||
| Swim | Intercept ** | −2.5467 | 0.6182 | 9 | −4.12 | 0.0026 | −3.9453 | −1.1482 | |
| Habitat *** | PPCC | 1.5279 | 0.0925 | 5355 | 16.53 | <0.0001 | 1.3466 | 1.7091 | |
| Habitat | Peng. | 0.0000 | . | . | . | . | . | . | |
| Season | Breeding | −0.1351 | 0.0999 | 5355 | −1.35 | 0.1762 | −0.3309 | 0.0607 | |
| Season *** | Molting | −0.7875 | −0.788 | 5355 | −6.19 | <0.0001 | −1.0369 | −0.5381 | |
| Season | Other | 0.0000 | . | . | . | . | . | . | |
| Lay | Intercept *** | −2.9178 | 0.4356 | 9 | −6.70 | <0.0001 | −3.9031 | −1.9324 | |
| Habitat *** | PPCC | 1.0181 | 0.1641 | 5356 | 6.20 | <0.0001 | 0.6963 | 1.3399 | |
| Habitat | Peng. | 0.0000 | . | . | . | . | . | . | |
| Time of day *** | A.M. | −1.3525 | 0.1631 | 5356 | −8.29 | <0.0001 | −1.6722 | −1.0328 | |
| Time of day | P.M. | 0.0000 | . | . | . | . | . | . | |
| Feed | Intercept *** | −5.6780 | 0.3158 | 8 | −17.98 | <0.0001 | −6.5063 | −4.9497 | |
| Habitat ** | PPCC | 0.6875 | 0.2653 | 5357 | 2.59 | 0.0096 | 0.1674 | 1.2076 | |
| Habitat | Peng. | 0.0000 | . | . | . | . | . | . | |
| Sex * | Female | 0.7156 | 0.3530 | 5357 | 2.03 | 0.0427 | 0.0237 | 1.4076 | |
| Sex | Male | 0.0000 | . | . | . | . | . | . | |
| Walk | Intercept *** | −0.7208 | 0.0945 | 9 | −7.63 | <0.0001 | −0.9345 | −0.5071 | |
| Habitat *** | PPCC | −0.7913 | 0.0679 | 5352 | −11.66 | <0.0001 | −0.9244 | −0.6583 | |
| Habitat | Peng. | 0.0000 | . | . | . | . | . | . | |
| Season *** | Breeding | 0.3272 | 0.0670 | 5352 | 4.88 | <0.0001 | 0.1959 | 0.4585 | |
| Season | Molting | −0.0126 | 0.0783 | 5352 | −0.16 | 0.8726 | −0.166 | 0.1409 | |
| Season | Other | 0.0000 | . | . | . | . | . | . | |
| All-Occurrence Behaviors | |||||||||
| Bathe | Intercept *** | −5.7962 | 0.4530 | 9 | −12.80 | <0.0001 | −6.8209 | −4.7716 | |
| Habitat *** | PPCC | 2.6686 | 0.2891 | 5355 | 9.23 | <0.0001 | 2.1019 | 3.2353 | |
| Habitat | Peng. | 0.0000 | . | . | . | . | . | . | |
| Season *** | Breeding | −0.9195 | 0.2718 | 5355 | −3.38 | 0.0007 | −1.4523 | −0.3867 | |
| Season | Molting | −0.00789 | 0.3034 | 5355 | −0.03 | 0.9792 | −0.6026 | 0.5868 | |
| Season | Other | 0.0000 | . | . | . | . | . | . | |
| Display | Intercept *** | −3.1501 | 0.1787 | 8 | −17.63 | <0.0001 | −3.5621 | −2.7381 | |
| Sex *** | Female | −0.8233 | 0.1928 | 5355 | −4.27 | <0.0001 | −1.2014 | −0.4453 | |
| Sex | Male | 0.0000 | . | . | . | . | . | . | |
| Habitat | PPCC | 0.3595 | 0.1950 | 5355 | 1.84 | 0.0652 | −0.0227 | 0.7417 | |
| Habitat | Peng. | 0.0000 | . | . | . | . | . | . | |
| Season *** | Breeding | −1.2276 | 0.2375 | 5355 | −5.17 | <0.0001 | −1.6932 | −0.7620 | |
| Season * | Molting | 0.5002 | 0.2363 | 5355 | 2.12 | 0.0343 | 0.0370 | 0.9634 | |
| Season | Other | 0.0000 | . | . | . | . | . | . | |
| Agonism | Intercept *** | −1.4374 | 0.1329 | 9 | −10.81 | <0.0001 | −1.7381 | −1.1367 | |
| Habitat *** | PPCC | −0.6516 | 0.1275 | 5353 | −5.11 | <0.0001 | −0.9015 | −0.4017 | |
| Habitat | Peng. | 0.0000 | . | . | . | . | . | . | |
| Season *** | Breeding | 0.7832 | 0.1348 | 5353 | 5.81 | <0.0001 | 0.5190 | 1.0474 | |
| Season * | Molting | −0.3763 | 0.1625 | 5353 | −2.32 | 0.0206 | −0.6948 | 0.0577 | |
| Season | Other | 0.0000 | . | . | . | . | . | . | |
| Proximity Measures | |||||||||
| Alone | Intercept | 0.1559 | 0.1254 | 9 | 1.24 | 0.2453 | −0.1278 | 0.4396 | |
| Habitat *** | PPCC | 0.2638 | 0.0365 | 5352 | 7.23 | <0.0001 | 0.0923 | 0.3353 | |
| Habitat | Peng. | 0.0000 | . | . | . | . | . | . | |
| Season *** | Breeding | 0.1742 | 0.0422 | 5352 | 4.13 | <0.0001 | 0.0916 | 0.2569 | |
| Season *** | Molting | 0.2603 | 0.0470 | 5352 | 5.54 | <0.0001 | 0.1682 | 0.3523 | |
| Season | Other | 0.0000 | . | . | . | . | . | . | |
| Time of day *** | A.M. | −0.090 | 0.0243 | 5352 | −3.71 | 0.0002 | −0.1378 | −0.0425 | |
| Time of day | P.M. | 0.0000 | . | . | . | . | . | . | |
| King | Intercept *** | 1.4124 | 0.2123 | 9 | 6.65 | <0.0001 | 0.9321 | 1.8928 | |
| Age *** | −0.06069 | 0.0075 | 5352 | −8.12 | <0.0001 | −0.075 | −0.0460 | ||
| Habitat *** | PPCC | −0.1451 | 0.0317 | 5352 | −4.58 | <0.0001 | −0.2073 | −0.0830 | |
| Habitat | Peng. | 0.0000 | . | . | . | . | . | . | |
| Season *** | Breeding | −0.1833 | 0.0371 | 5352 | −4.94 | <0.0001 | −0.2561 | −0.1105 | |
| Season *** | Molting | −0.1865 | 0.0417 | 5352 | −4.48 | <0.0001 | −0.2682 | −0.1048 | |
| Season | Other | 0.0000 | . | . | . | . | . | . | |
| Other | Intercept *** | −4.3134 | 0.2003 | 9 | −21.54 | <0.0001 | −4.7665 | −3.8603 | |
| Habitat *** | PPCC | −1.0387 | 0.2600 | 5357 | −3.99 | <0.0001 | −1.5484 | −0.5289 | |
| Habitat | Peng. | 0.0000 | . | . | . | . | . | . | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuller, G.; Jones, M.; Gartland, K.N.; Zalewski, S.; Heintz, M.R.; Allard, S. The Benefits of Increased Space and Habitat Complexity for the Welfare of Zoo-Housed King Penguins (Aptenodytes patagonicus). Animals 2023, 13, 2312. https://doi.org/10.3390/ani13142312
Fuller G, Jones M, Gartland KN, Zalewski S, Heintz MR, Allard S. The Benefits of Increased Space and Habitat Complexity for the Welfare of Zoo-Housed King Penguins (Aptenodytes patagonicus). Animals. 2023; 13(14):2312. https://doi.org/10.3390/ani13142312
Chicago/Turabian StyleFuller, Grace, Megan Jones, Kylen N. Gartland, Sara Zalewski, Matthew R. Heintz, and Stephanie Allard. 2023. "The Benefits of Increased Space and Habitat Complexity for the Welfare of Zoo-Housed King Penguins (Aptenodytes patagonicus)" Animals 13, no. 14: 2312. https://doi.org/10.3390/ani13142312
APA StyleFuller, G., Jones, M., Gartland, K. N., Zalewski, S., Heintz, M. R., & Allard, S. (2023). The Benefits of Increased Space and Habitat Complexity for the Welfare of Zoo-Housed King Penguins (Aptenodytes patagonicus). Animals, 13(14), 2312. https://doi.org/10.3390/ani13142312