
Analysis of Morphological Traits, Mineralization, and Mechanical Properties of Femoral Bones in Young and Adult European Hares (Lepus europaeus)

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Characterization of the Research Area
2.2. Research Material
2.3. DXA Measurements
2.4. Bone Analysis
2.5. Statistical Analysis
3. Results
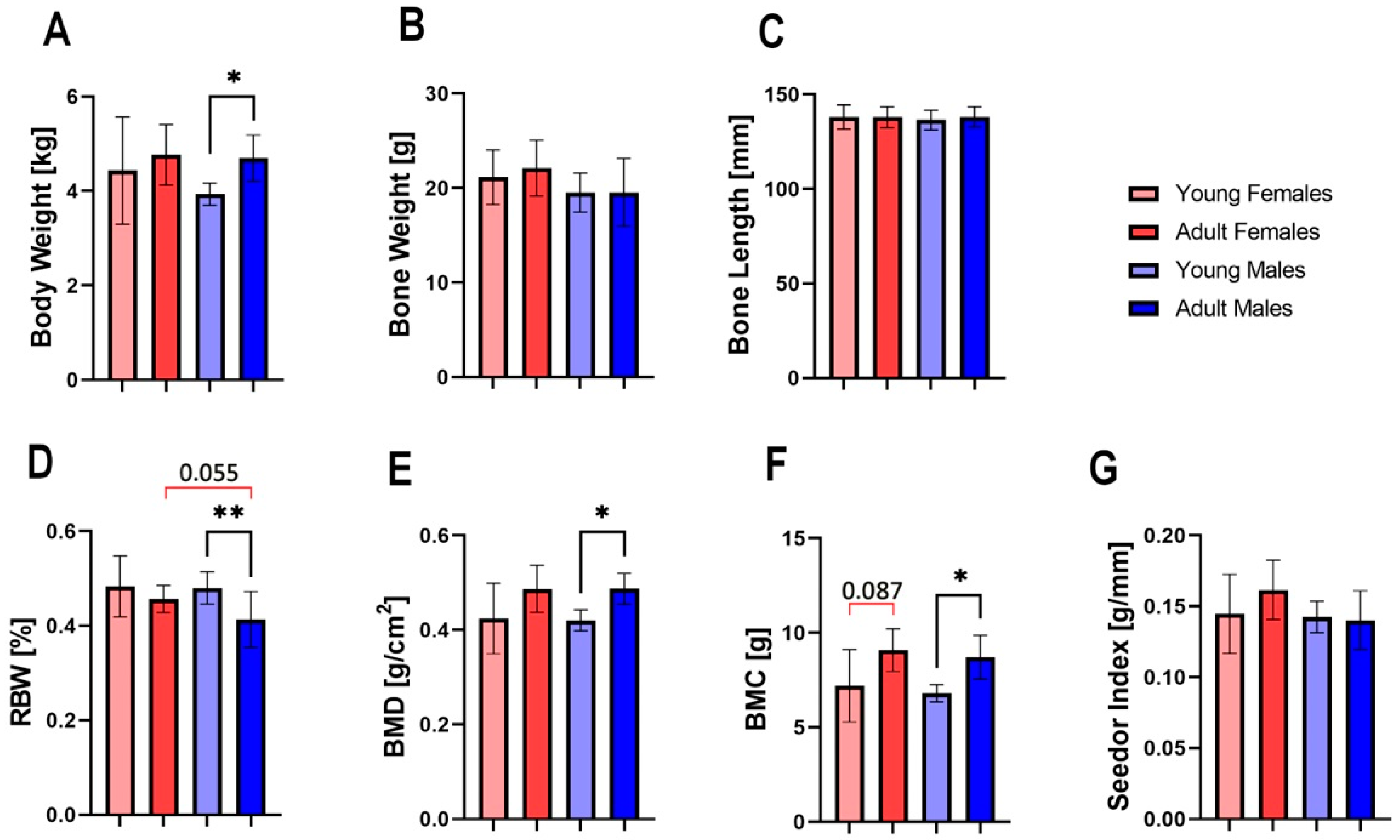
3.1. Bone Properties
3.2. Geometrical Properties
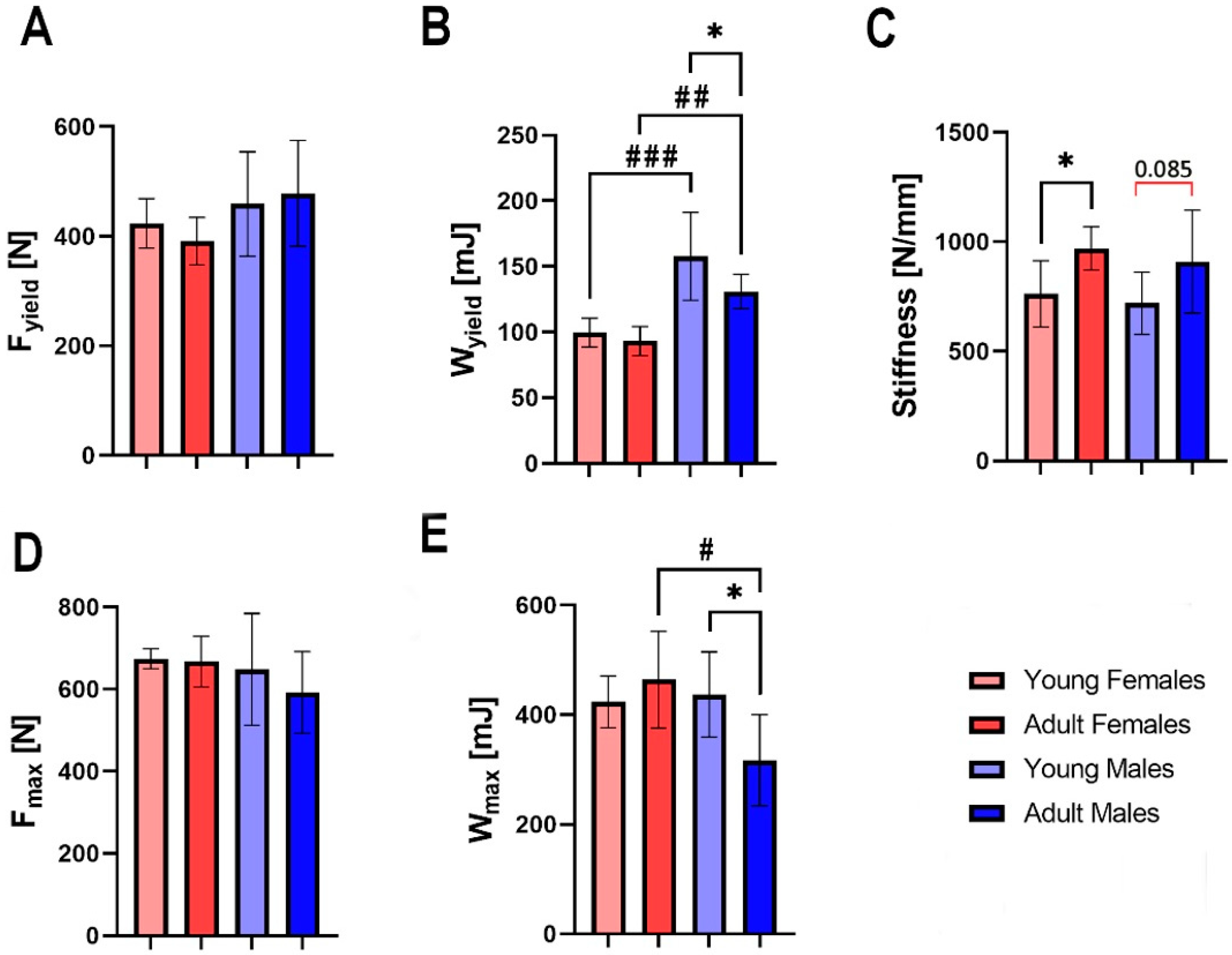
3.3. Mechanical Properties
3.4. Bone Material Properties
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ge, D.; Wen, Z.; Xia, L.; Zhang, Z.; Erbajeva, M.; Huang, C.; Yang, Q. Evolutionary History of Lagomorphs in Response to Global Environmental Change. PLoS ONE 2013, 8, e59668. [Google Scholar] [CrossRef] [PubMed]
- Thulin, C.-G. The Distribution of Mountain Hares Lepus Timidus in Europe: A Challenge from Brown Hares (L. europaeus)?: Distribution of European Mountainhares. Mamm. Rev. 2003, 33, 29–42. [Google Scholar] [CrossRef]
- Reese, A.T.; Lanier, H.C.; Sargis, E.J. Skeletal Indicators of Ecological Specialization in Pika (Mammalia, Ochotonidae): Ecological Specialization in Pika. J. Morphol. 2013, 274, 585–602. [Google Scholar] [CrossRef] [PubMed]
- Young, J.W.; Danczak, R.; Russo, G.A.; Fellmann, C.D. Limb Bone Morphology, Bone Strength, and Cursoriality in Lagomorphs. J. Anat. 2014, 225, 403–418. [Google Scholar] [CrossRef]
- Martin, E.M.; Young, J.W.; Fellmann, C.D.; Kraatz, B.; Sherratt, E. Anatomical Correlates of Cursoriality Are Compromised by Body Size and Propensity to Burrow in a Group of Small Mammals (Lagomorpha). Evol. Biol. 2022, 49, 464–481. [Google Scholar] [CrossRef]
- Terres, J.K. Speed of the Varying Hare. J. Mammal. 1941, 22, 453–454. [Google Scholar] [CrossRef]
- Herrel, A.; Gibb, A.C. Ontogeny of Performance in Vertebrates. Physiol. Biochem. Zool. 2006, 79, 1–6. [Google Scholar] [CrossRef]
- Carrier, D.R. Postnatal Ontogeny of the Musculo-skeletal System in the Black-tailed Jack Rabbit (Lepus californicus). J. Zool. 1983, 201, 27–55. [Google Scholar] [CrossRef]
- Carrier, D. Ontogeny of Jumping Performance in the Black-Tailed Jackrabbit (Lepus californicus). Zoology 1995, 98, 309–313. Available online: https://www.researchgate.net/publication/284772783_Ontogeny_of_jumping_performance_in_the_black-tailed_jackrabbit_Lepus_californicus (accessed on 1 May 2023).
- Williams, S.B.; Payne, R.C.; Wilson, A.M. Functional Specialisation of the Pelvic Limb of the Hare (Lepus europeus). J. Anat. 2007, 210, 472–490. [Google Scholar] [CrossRef]
- Stein, B.R.; Casinos, A. What Is a Cursorial Mammal? J. Zool. 1997, 242, 185–192. [Google Scholar] [CrossRef]
- Morgan, E.F.; Unnikrisnan, G.U.; Hussein, A.I. Bone Mechanical Properties in Healthy and Diseased States. Annu. Rev. Biomed. Eng. 2018, 20, 119–143. [Google Scholar] [CrossRef] [PubMed]
- Burstein, A.H.; Reilly, D.T.; Martens, M. Aging of Bone Tissue: Mechanical Properties. J. Bone Joint Surg. Am. 1976, 58, 82–86. Available online: https://pubmed.ncbi.nlm.nih.gov/1249116/ (accessed on 1 May 2023). [CrossRef] [PubMed]
- Kondracki, J. Geografia Regionalna Polski, 3rd ed.; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2022; ISBN 978-83-01-16022-7. [Google Scholar]
- Flis, M.; Grela, E.R.; Gugała, D.; Rataj, B. Seasonality of the Reproduction of Wild Boars in the Lublin Upland and Weight Characteristics of Their Carcasses. Med. Wet. 2018, 74, 477–480. [Google Scholar] [CrossRef]
- Bray, Y.; Champely, S.; Soyez, D. Age Determination in Leverets of European Hare (Lepus europaeus) Based on Body Measurements. Wildlife Biol. 2002, 8, 31–39. [Google Scholar] [CrossRef]
- Pintur, K.; Popović, N.; Alegro, A.; Severin, K.; Slavica, A.; Kolić, E. Selected Indicators of Brown Hare (Lepus europaeus Pallas, 1778) Population Dynamics in Northwestern Croatia. Vet. Arh. 2006, 76, S199–S209. Available online: http://vetarhiv.vef.unizg.hr/papers/2006-76-7-23.pdf (accessed on 1 May 2023).
- Flis, M.; Czyżowski, P.; Beeger, S.; Rataj, B.; Karpiński, M. Logistic Regression Model for Determination of the Age of Brown Hare (Lepus europaeus Pall.) Based on Body Weight. Animals 2022, 12, 529. [Google Scholar] [CrossRef]
- Muszyński, S.; Kwiecień, M.; Tomaszewska, E.; Świetlicka, I.; Dobrowolski, P.; Kasperek, K.; Jeżewska-Witkowska, G. Effect of Caponization on Performance and Quality Characteristics of Long Bones in Polbar Chickens. Poult. Sci. 2017, 96, 491–500. [Google Scholar] [CrossRef]
- Muszyński, S.; Dajnowska, A.; Arciszewski, M.B.; Rudyk, H.; Śliwa, J.; Krakowiak, D.; Piech, M.; Nowakowicz-Dębek, B.; Czech, A. Effect of Fermented Rapeseed Meal in Feeds for Growing Piglets on Bone Morphological Traits, Mechanical Properties, and Bone Metabolism. Animals 2023, 13, 1080. [Google Scholar] [CrossRef]
- Alexander, R.M.; Jayes, A.S.; Maloiy, G.M.O.; Wathuta, E.M. Allometry of the Limb Bones of Mammals from Shrews (Sorex) to Elephant (Loxodonta). J. Zool. 1979, 189, 305–314. [Google Scholar] [CrossRef]
- Biewener, A.A. Biomechanics of Mammalian Terrestrial Locomotion. Science 1990, 250, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Alexander, R.; Vernon, A. The Mechanics of Hopping by Kangaroos (Macropodidae). J. Zool. 1975, 177, 265–303. [Google Scholar] [CrossRef]
- Brown, J.C.; Yalden, D.W. The Description of Mammals–2 Limbs and Locomotion of Terrestial Mammals. Mamm. Rev. 1973, 3, 107–134. [Google Scholar] [CrossRef]
- Camp, C.L.; Borell, A.E. Skeletal and Muscular Differences in the Hind Limbs of Lepus, Sylvilagus, and Ochotona. J. Mammal. 1937, 18, 315. [Google Scholar] [CrossRef]
- Stott, P. Use of Space by Sympatric European Hares (Lepus europaeus) and European Rabbits (Oryctolagus cuniculus) in Australia. Mamm. Biol. 2003, 68, 317–327. [Google Scholar] [CrossRef]
- Young, J.W.; Foster, A.D.; Russo, G.A.; Smith, G.A.; Butcher, M.T. Only the Good Die Old? Ontogenetic Determinants of Locomotor Performance in Eastern Cottontail Rabbits (Sylvilagus floridanus). Integr. Org. Biol. 2022, 4, obab037. [Google Scholar] [CrossRef]
- Roberts, T.J.; Scales, J.A. Mechanical Power Output during Running Accelerations in Wild Turkeys. J. Exp. Biol. 2002, 205, 1485–1494. [Google Scholar] [CrossRef]
- Williams, S.B.; Tan, H.; Usherwood, J.R.; Wilson, A.M. Pitch Then Power: Limitations to Acceleration in Quadrupeds. Biol. Lett. 2009, 5, 610–613. [Google Scholar] [CrossRef]
- Keith, L.B.; Bloomer, S.E.M. Differential Mortality of Sympatric Snowshoe Hares and Cottontail Rabbits in Central Wisconsin. Can. J. Zool. 1993, 71, 1694–1697. [Google Scholar] [CrossRef]
- McGowan, C.P.; Baudinette, R.V.; Biewener, A.A. Joint Work and Power Associated with Acceleration and Deceleration in Tammar Wallabies (Macropus eugenii). J. Exp. Biol. 2005, 208, 41–53. [Google Scholar] [CrossRef]
- Monte, A.; Nardello, F.; Zamparo, P. Mechanical Advantage and Joint Function of the Lower Limb during Hopping at Different Frequencies. J. Biomech. 2021, 118, 110294. [Google Scholar] [CrossRef]
- Currey, J.D. Bones: Structure and Mechanics; Princeton Univ. Press: Princeton, NJ, USA, 2006; ISBN 978-0-691-12804-7. [Google Scholar]
- Hart, N.H.; Nimphius, S.; Rantalainen, T.; Ireland, A.; Siafarikas, A.; Newton, R.U. Mechanical Basis of Bone Strength: Influence of Bone Material, Bone Structure and Muscle Action. J. Musculoskelet. Neur. Interact. 2017, 17, 114–139. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5601257/ (accessed on 1 May 2023).
- Foster, A.D.; Butcher, M.T.; Smith, G.A.; Russo, G.A.; Thalluri, R.; Young, J.W. Ontogeny of Effective Mechanical Advantage in Eastern Cottontail Rabbits (Sylvilagus floridanus). J. Exp. Biol. 2019, jeb.205237. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Shen, J.; Zhang, J.; Zhou, A.; Guo, W. Correlations between Horizontal Jump and Sprint Acceleration and Maximal Speed Performance: A Systematic Review and Meta-Analysis. PeerJ 2023, 11, e14650. [Google Scholar] [CrossRef]
- Wilson:, M.P.; Espinoza, N.R.; Shah, S.R.; Blob, R.W. Mechanical Properties of the Hindlimb Bones of Bullfrogs and Cane Toads in Bending and Torsion. Anat. Rec. 2009, 292, 935–944. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osiak-Wicha, C.; Tomaszewska, E.; Muszyński, S.; Flis, M.; Arciszewski, M.B. Analysis of Morphological Traits, Mineralization, and Mechanical Properties of Femoral Bones in Young and Adult European Hares (Lepus europaeus). Animals 2023, 13, 2077. https://doi.org/10.3390/ani13132077
Osiak-Wicha C, Tomaszewska E, Muszyński S, Flis M, Arciszewski MB. Analysis of Morphological Traits, Mineralization, and Mechanical Properties of Femoral Bones in Young and Adult European Hares (Lepus europaeus). Animals. 2023; 13(13):2077. https://doi.org/10.3390/ani13132077
Chicago/Turabian StyleOsiak-Wicha, Cezary, Ewa Tomaszewska, Siemowit Muszyński, Marian Flis, and Marcin B. Arciszewski. 2023. "Analysis of Morphological Traits, Mineralization, and Mechanical Properties of Femoral Bones in Young and Adult European Hares (Lepus europaeus)" Animals 13, no. 13: 2077. https://doi.org/10.3390/ani13132077
APA StyleOsiak-Wicha, C., Tomaszewska, E., Muszyński, S., Flis, M., & Arciszewski, M. B. (2023). Analysis of Morphological Traits, Mineralization, and Mechanical Properties of Femoral Bones in Young and Adult European Hares (Lepus europaeus). Animals, 13(13), 2077. https://doi.org/10.3390/ani13132077