
Troglonectes canlinensis sp. nov. (Teleostei: Nemacheilidae), a New Troglomorphic Loach from Guangxi, China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
| Identification Key to Species of Troglonectes |
| 1. Eye present··················································································································································································································2 |
| –. Eye degenerated or absent·····················································································································································································5 |
| 2. Body scaled after dorsal fin origin·········································································································································Tr. furcocaudalis |
| –. Whole body scaled except for head and thorax··············································································································································3 |
| 3. Caudal fin forked·············································································································································································Tr. duanensis |
| –. Caudal fin truncated································································································································································································4 |
| 4. Caudal peduncle length 12.0–13.6% SL··································································································································Tr. hechiensis |
| –. Caudal peduncle length 10.8–12.0% SL·····························································································································Tr. retrodorsalis |
| 5. Eye degenerated with black pigment····························································································································································6 |
| –. Eye absent··················································································································································································································10 |
| 6. Body scaleless·············································································································································································································7 |
| –. Whole body scaled except for head and thorax··············································································································································8 |
| 7. Upper adipose keel height larger than caudal peduncle depth·············································································Tr. microphthalmus |
| –. Upper adipose keel height mostly 1/2 the caudal peduncle depth···········································································Tr. donglanensis |
| 8. Posterior chamber of swim bladder degenerated············································································································Tr. lingyunensis |
| –. Posterior chamber of swim bladder developed·············································································································································9 |
| 9. Total of 12–13 inner gill rakers on first gill arch·····················································································································Tr. macrolepis |
| –. Total of 8–10 inner gill rakers on first gill arch······································································································Tr. canlinensis sp. nov. |
| 10. Caudal fin truncated································································································································································Tr. shuilongensis |
| –. Caudal fin emarginated or forked····································································································································································11 |
| 11. Caudal fin emarginated······················································································································································································12 |
| –. Caudal fin forked····································································································································································································14 |
| 12. Lateral line complete···································································································································································Tr. jiarongensis |
| –. Lateral line incomplete or absent······································································································································································13 |
| 13. Lateral line incomplete································································································································································Tr. translucens |
| –. Lateral line absent················································································································································································Tr. lihuensis |
| 14. Lateral line absent································································································································································································15 |
| –. Lateral line complete or incomplete································································································································································16 |
| 15. Standard length 2.6–3.5 times the lateral head length·······································································································Tr. barbatus |
| –. Standard length 4.3–4.9 times the lateral head length································································································Tr. huanjiangensis |
| 16. Lateral line complete···························································································································································································17 |
| –. Lateral line incomplete·························································································································································································19 |
| 17. Dorsal fin with six branched rays, anal fin with four branched rays······································································Tr. maolanensis |
| –. Dorsal fin with eight or nine branched rays, anal fin with six branched rays···················································································18 |
| 18. Standard length 10.1–14.0 times the caudal peduncle depth··················································································Tr. daqikongensis |
| –. Standard length 14.5–18.1 times the caudal peduncle depth····················································································Tr. longibarbatus |
| 19. Pelvic fin origin opposite the dorsal fin origin···············································································································Tr. dongganensis |
| –. Pelvic fin origin anterior to the dorsal fin origin······················································································································Tr. elongatus |
4. Discussion
5. Conclusions
6. Nomenclatural Acts Registration
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Du, L.N.; Chen, X.Y.; Yang, J.X. A review of the Nemacheilinae genus Oreonectes Günther with descriptions of two new species (Teleostei: Balitoridae). Zootaxa 2008, 1729, 23–36. [Google Scholar] [CrossRef]
- Zhang, C.G.; Xing, Y.C.; Zhao, Y.H.; Zhou, W.; Tang, W.Q. Species Diversity and Distribution of Inland Fishes in China; Science Publishing Company: Beijing, China, 2017; pp. 127–151. (In Chinese) [Google Scholar]
- Chen, Y.R.; Yang, J.X.; Sket, B.; Aljancic, G. A new blind cave loach of Paracobitis with comment on its characters evolution. Zool. Res. 1998, 19, 59–63. [Google Scholar]
- Li, W.X.; Ran, J.C.; Chen, H.M. A new species of Paracobitis from Guizhou, China. J. Zhanjiang Ocean Univ. 2006, 26, 1–2. [Google Scholar]
- Lin, Y.; Li, C.; Song, J.K. A new species of troglobitic loach (Cypriniformes, Balitoridae), Triplophysa jiarongensis, from GuiZhou, China. Acta Zootaxonomica Sin. 2012, 37, 640–647. [Google Scholar]
- Huang, J.Q.; Yang, J.; Wu, Z.Q.; Zhao, Y.H. Oreonectes guilinensis (Teleostei, Cypriniformes, Nemacheilidae), a new loach species from Guangxi, China. J. Fish Biol. 2020, 96, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Wang, Y.L.; Xiao, N.; Yang, X.; Zhou, J.J.; Deng, H.Q.; Zhou, J. Phylogenetic relationships of the Chinese cave loaches (Cypriniformes, Nemacheilidae) revealed by phylogenetic analysis of mitochondrial. Zool. Res. 2023, 44, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.X.; Liu, J.H.; Du, L.N.; Luo, F.G. A new loach species of Troglonectes (Teleostei: Nemacheilidae) from Guangxi, China. Zool. Res. 2021, 42, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Du, L.N.; Yang, J.; Min, R.; Chen, X.Y.; Yang, J.X. A review of the Cypriniform tribe Yunnanilini Prokofiev, 2010 from China, with an emphasis on five genera based on morphologies and complete mitochondrial genomes of some species. Zool. Res. 2021, 42, 310–334. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Zhao, Y.H.; Zhang, C.G. A new blind loach, Oreonectes elongatus sp. nov. (Cypriniformes: Balitoridae) from Guangxi, China. Environ. Biol. Fishes 2012, 93, 483–490. [Google Scholar] [CrossRef]
- Lan, J.H.; Gan, X.; Wu, T.J.; Yang, J. Cave Fishes of Guangxi, China; Science Press: Beijing, China, 2013; pp. 104–139. (In Chinese) [Google Scholar]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Ma, L.; Zhao, Y.H.; Yang, J.X. Chapter 28—Cavefish of China. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 237–254. [Google Scholar]

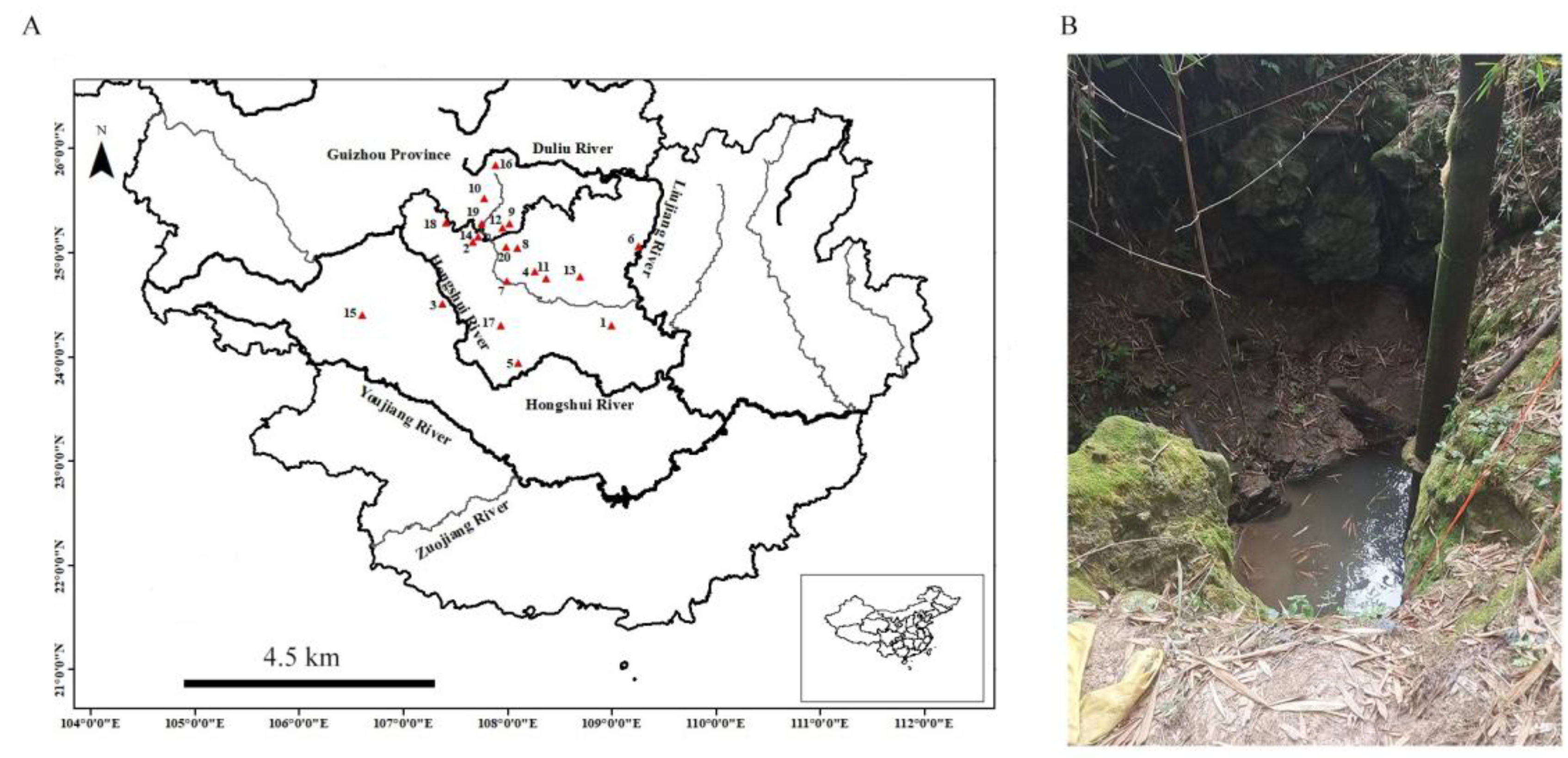

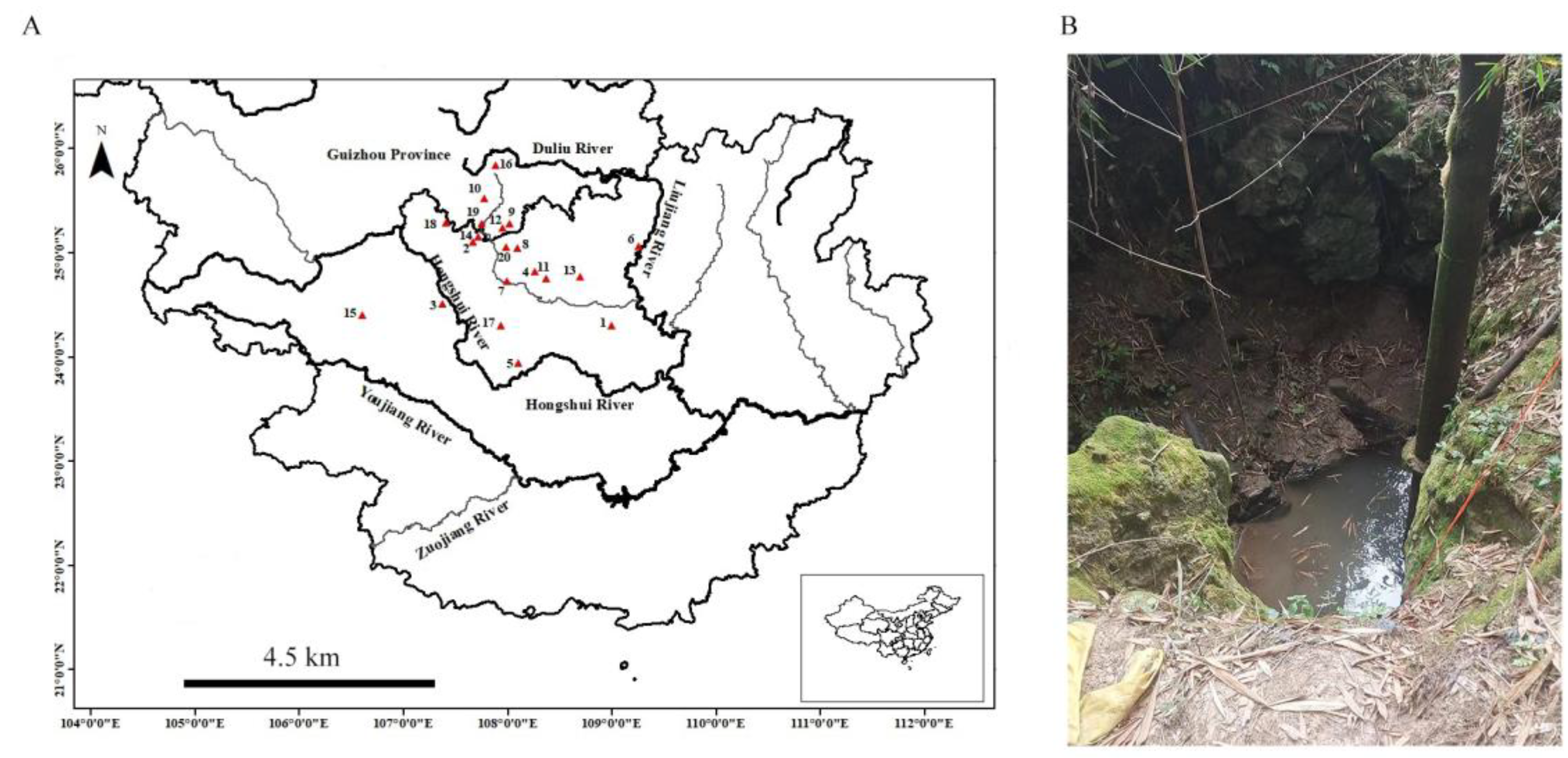
{kind=link}
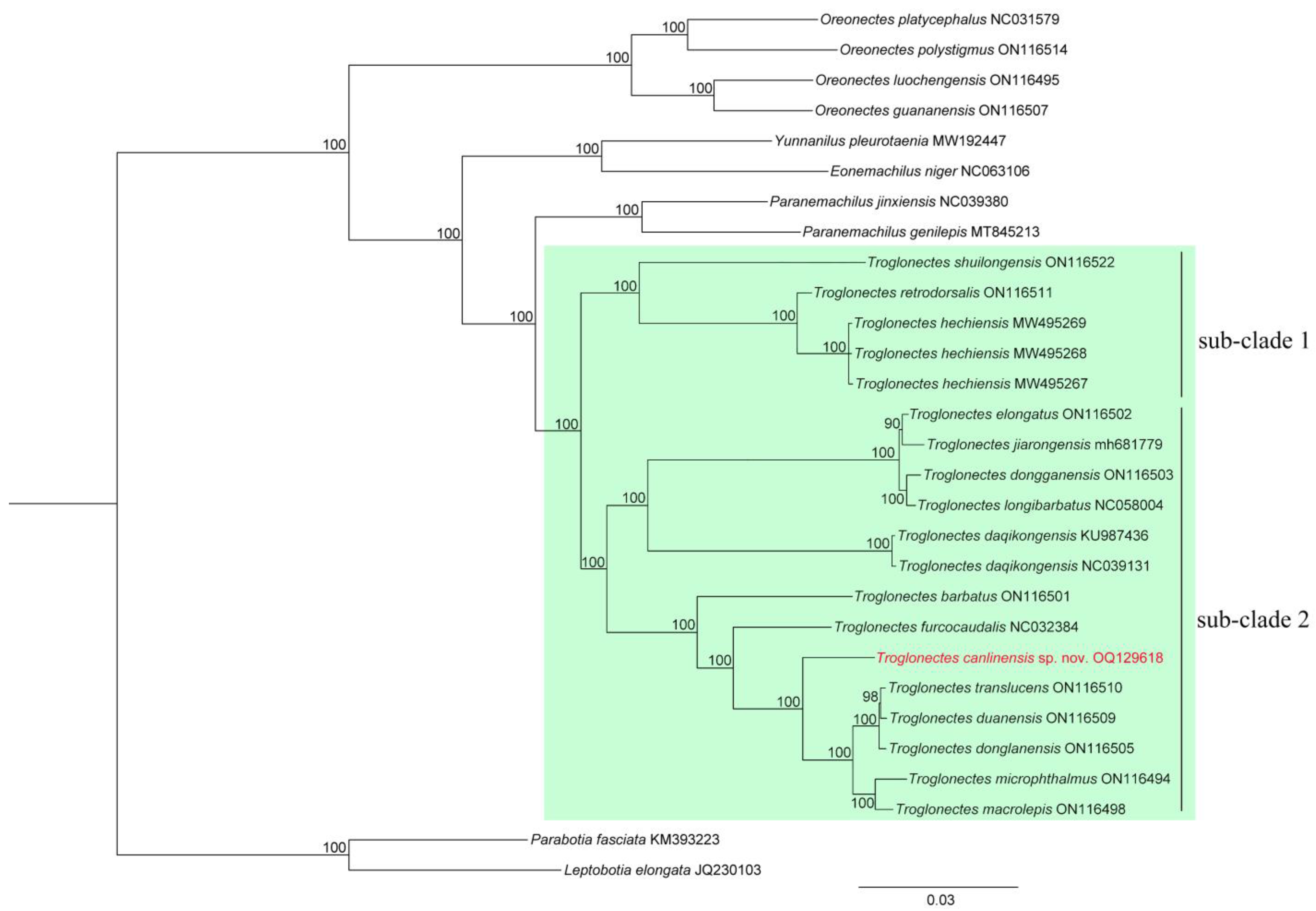
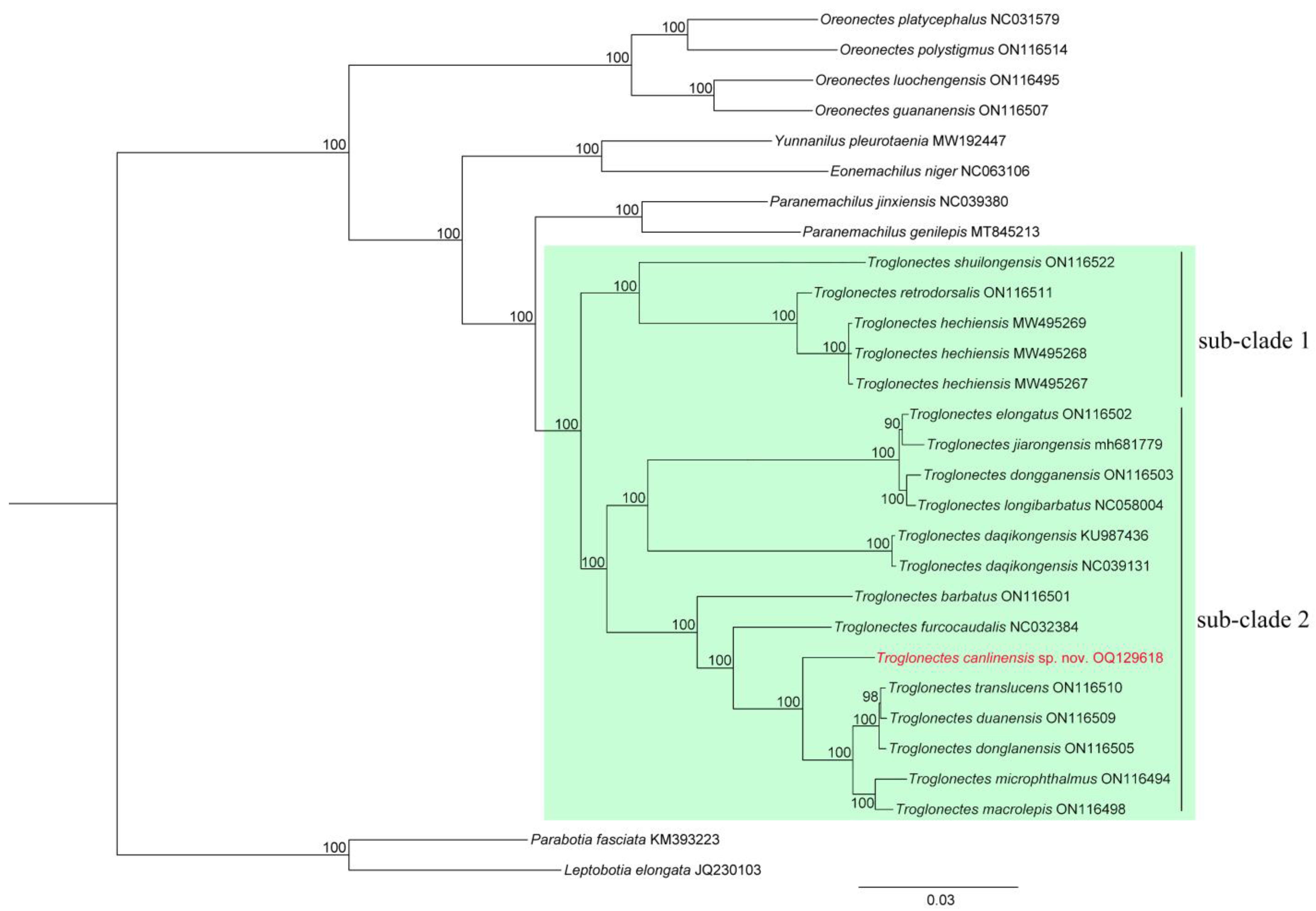
{kind=link}
{kind=link}
| Holotype | Range (n = 10) | Mean ± SD | |
|---|---|---|---|
| Total length (mm) | 43.0 | 35.6–65.3 | 46.1 ± 8.6 |
| Standard length (mm) | 36.0 | 29.9–54.3 | 38.7 ± 7.3 |
| Percentage of standard length (%) | |||
| Body depth | 22.0 | 18.2–22.0 | 19.8 ± 1.1 |
| Lateral head length | 28.0 | 25.8–29.6 | 27.7 ± 1.1 |
| Predorsal length | 57.8 | 54.2–58.7 | 56.9 ± 1.2 |
| Prepelvic length | 61.0 | 57.8–61.2 | 59.8 ± 1.1 |
| Preanal length | 80.0 | 75.6–81.3 | 79.3 ± 1.7 |
| Preanus length | 75.7 | 74.1–77.4 | 75.9 ± 1.2 |
| Caudal peduncle length | 11.5 | 10.9–14.6 | 12.8 ± 1.3 |
| Caudal peduncle depth | 13.9 | 11.3–13.9 | 12.2 ± 0.7 |
| Head width | 18.1 | 16.0–18.1 | 16.9 ± 0.8 |
| Pectoral fin length | 16.1 | 15.3–16.7 | 16.1 ± 0.4 |
| Pelvic fin length | 11.4 | 11.4–14.3 | 12.9 ± 0.8 |
| Percentage of lateral head length (%) | |||
| Eye diameter | 10.3 | 7.5–11.6 | 10.1 ± 1.1 |
| Interorbital width | 35.81 | 26.5–39.4 | 32.4 ± 4.4 |
| Snout length | 35.5 | 34.6–46.6 | 40.5 ± 3.4 |
| Head width | 64.5 | 54.2–67.6 | 61.3 ± 4.0 |
| Head depth | 51.6 | 46.9–55.9 | 51.3 ± 2.6 |
| Maxillary barbel length | 28.6 | 19.3–37.1 | 29.1 ± 5.4 |
| Outer barbel length | 26.7 | 21.9–36.3 | 29.7 ± 3.9 |
| Inner barbel length | 15.8 | 12.0–19.5 | 17.4 ± 2.2 |
| Percentage of caudal peduncle length (%) | |||
| Caudal peduncle depth | 120.5 | 84.1–120.5 | 96.5 ± 12.5 |
| Dorsal fin rays | 4, 9 | 4, 8–9 | |
| Pectoral fin rays | 1, 9 | 1, 9–10 | |
| Pelvic fin rays | 1, 6 | 1, 5–6 | |
| Anal fin rays | 3, 6 | 3, 5–6 | |
| Caudal fin rays | 14 | 13–14 | |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Tr. barbatus | - | ||||||||||||||
| 2 | Tr. canlinensis | 0.075 | ||||||||||||||
| 3 | Tr. daqikongensis | 0.105 | 0.090 | |||||||||||||
| 4 | Tr. dongganensis | 0.119 | 0.099 | 0.107 | ||||||||||||
| 5 | Tr. donglanensis | 0.072 | 0.033 | 0.101 | 0.096 | |||||||||||
| 6 | Tr. duanensis | 0.071 | 0.033 | 0.102 | 0.095 | 0.003 | ||||||||||
| 7 | Tr. elongatus | 0.118 | 0.098 | 0.108 | 0.010 | 0.093 | 0.092 | |||||||||
| 8 | Tr. furcocaudalis | 0.066 | 0.048 | 0.096 | 0.098 | 0.046 | 0.043 | 0.097 | ||||||||
| 9 | Tr. hechiensis | 0.099 | 0.098 | 0.102 | 0.098 | 0.085 | 0.084 | 0.097 | 0.084 | |||||||
| 10 | Tr. jiarongensis | 0.122 | 0.098 | 0.107 | 0.010 | 0.098 | 0.097 | 0.007 | 0.101 | 0.096 | ||||||
| 11 | Tr. longibarbatus | 0.117 | 0.095 | 0.103 | 0.004 | 0.092 | 0.091 | 0.006 | 0.096 | 0.094 | 0.006 | |||||
| 12 | Tr. macrolepis | 0.068 | 0.030 | 0.099 | 0.097 | 0.016 | 0.017 | 0.096 | 0.047 | 0.084 | 0.098 | 0.093 | ||||
| 13 | Tr. microphthalmus | 0.069 | 0.031 | 0.098 | 0.098 | 0.014 | 0.014 | 0.097 | 0.046 | 0.086 | 0.099 | 0.094 | 0.008 | |||
| 14 | Tr. retrodorsalis | 0.097 | 0.079 | 0.099 | 0.098 | 0.085 | 0.084 | 0.097 | 0.082 | 0.012 | 0.095 | 0.094 | 0.081 | 0.082 | ||
| 15 | Tr. shuilongensis | 0.115 | 0.099 | 0.109 | 0.100 | 0.101 | 0.100 | 0.097 | 0.101 | 0.089 | 0.101 | 0.096 | 0.102 | 0.098 | 0.085 | |
| 16 | Tr. translucens | 0.072 | 0.031 | 0.100 | 0.094 | 0.002 | 0.003 | 0.091 | 0.046 | 0.083 | 0.096 | 0.090 | 0.016 | 0.014 | 0.083 | 0.100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.-J.; Ge, J.-K.; Bao, C.-Y.; Du, L.-N.; Luo, F.-G.; Zou, T.-X. Troglonectes canlinensis sp. nov. (Teleostei: Nemacheilidae), a New Troglomorphic Loach from Guangxi, China. Animals 2023, 13, 1712. https://doi.org/10.3390/ani13101712
Li S-J, Ge J-K, Bao C-Y, Du L-N, Luo F-G, Zou T-X. Troglonectes canlinensis sp. nov. (Teleostei: Nemacheilidae), a New Troglomorphic Loach from Guangxi, China. Animals. 2023; 13(10):1712. https://doi.org/10.3390/ani13101712
Chicago/Turabian StyleLi, Shu-Jing, Jia-Kai Ge, Chun-Yan Bao, Li-Na Du, Fu-Guang Luo, and Tong-Xiang Zou. 2023. "Troglonectes canlinensis sp. nov. (Teleostei: Nemacheilidae), a New Troglomorphic Loach from Guangxi, China" Animals 13, no. 10: 1712. https://doi.org/10.3390/ani13101712
APA StyleLi, S.-J., Ge, J.-K., Bao, C.-Y., Du, L.-N., Luo, F.-G., & Zou, T.-X. (2023). Troglonectes canlinensis sp. nov. (Teleostei: Nemacheilidae), a New Troglomorphic Loach from Guangxi, China. Animals, 13(10), 1712. https://doi.org/10.3390/ani13101712