

Polysaccharide of Atractylodes macrocephala Koidz Alleviates Cyclophosphamide-Induced Thymus Ferroptosis in Gosling
 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
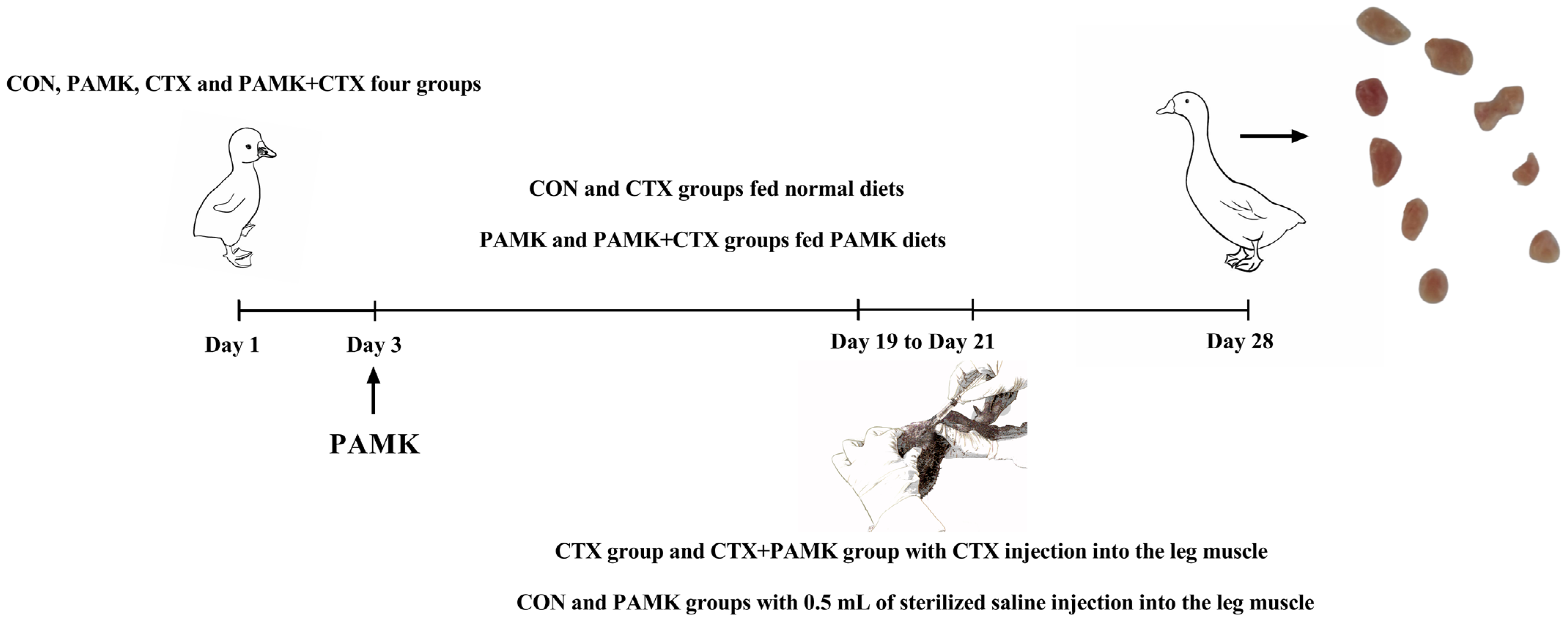
2.1. Animal Experiments
2.2. Transcriptome Sequencing
2.3. Real-Time Quantitative PCR Assay
2.4. Western Blot Analysis
2.5. Thymus Histology
2.6. Ultramicroscopic Morphology Observation
2.7. Immunofluorescence Staining
2.8. ROS Assay
2.9. Glutatione (GSH), Malonedialdehyde (MDA), and Tissue Fe Level Assays
2.10. Statistical Analysis
3. Results
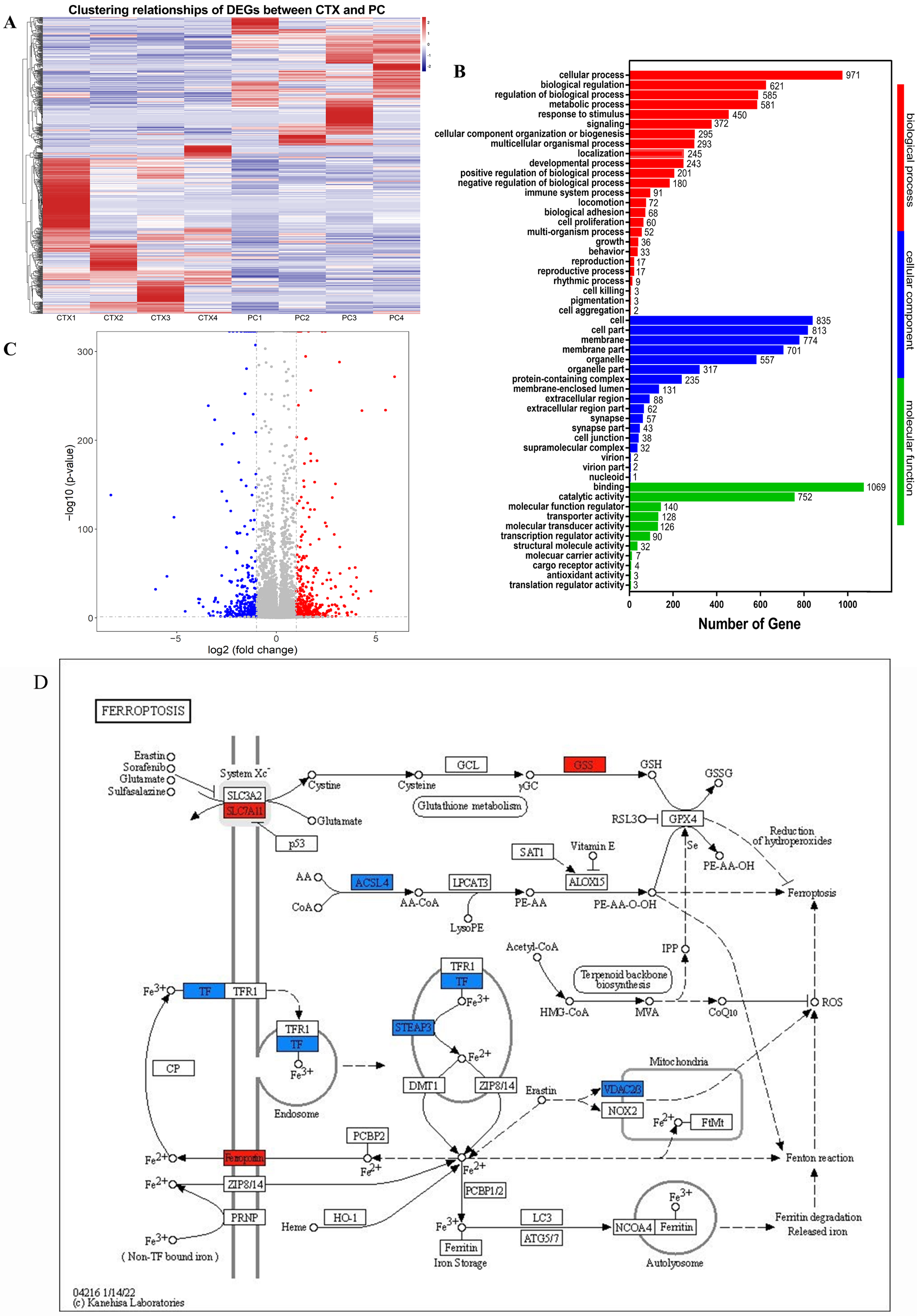
3.1. Transcriptome Analysis Reveals PAMK Might Regulate Ferroptosis Pathway
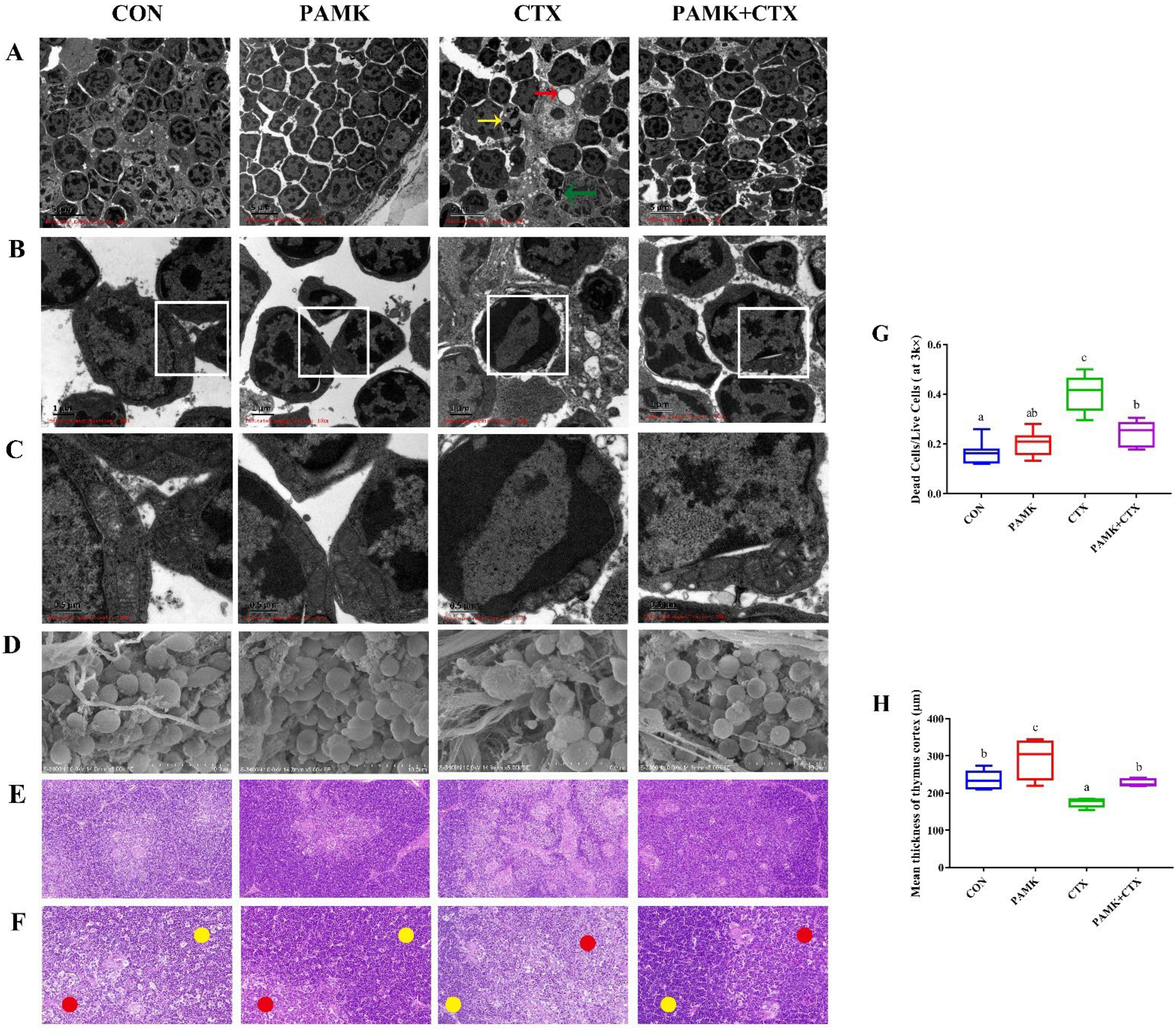
3.2. PAMK Alleviates CTX-Induced Thymus Injury, Cell Death, and Mitochondrial Damage
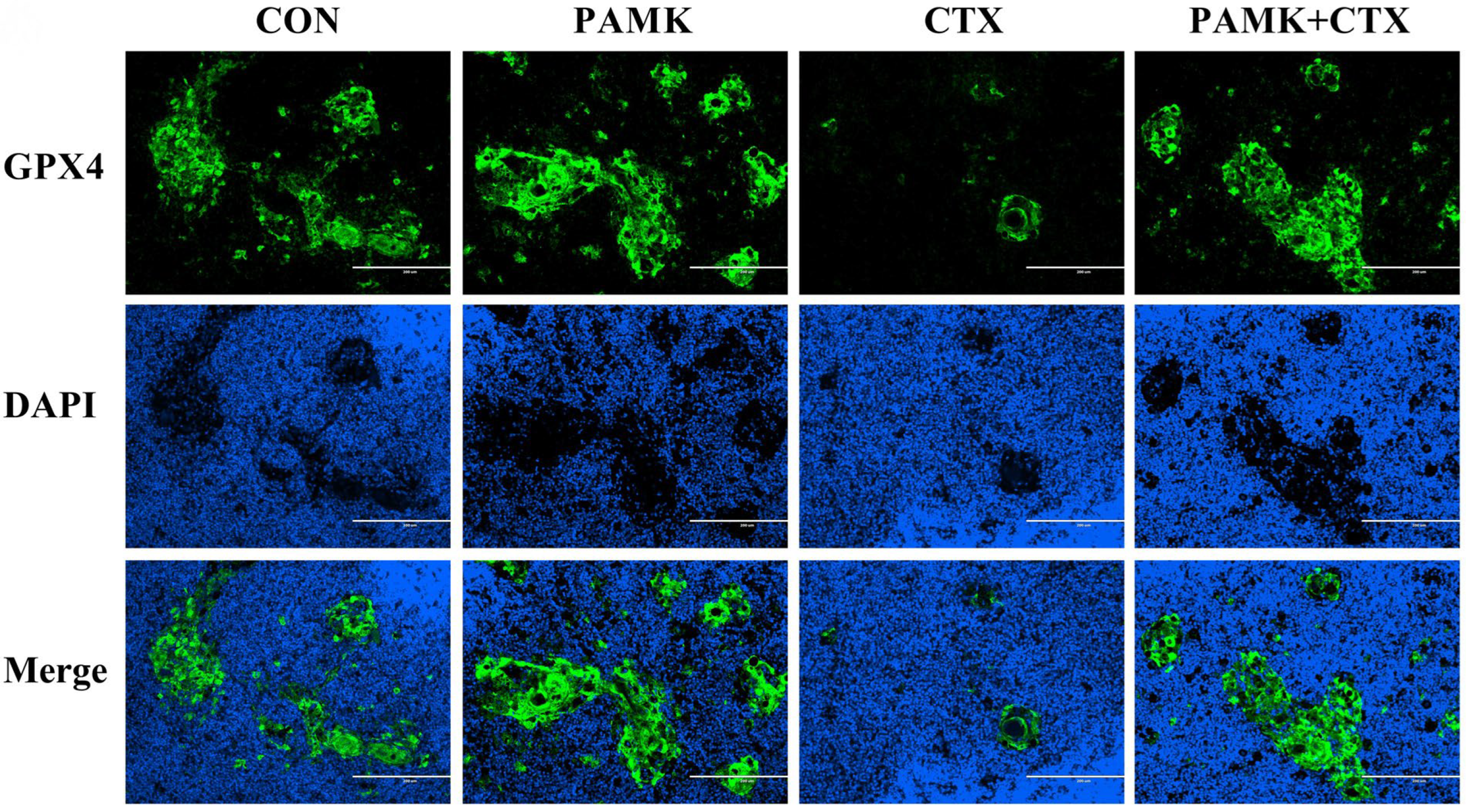
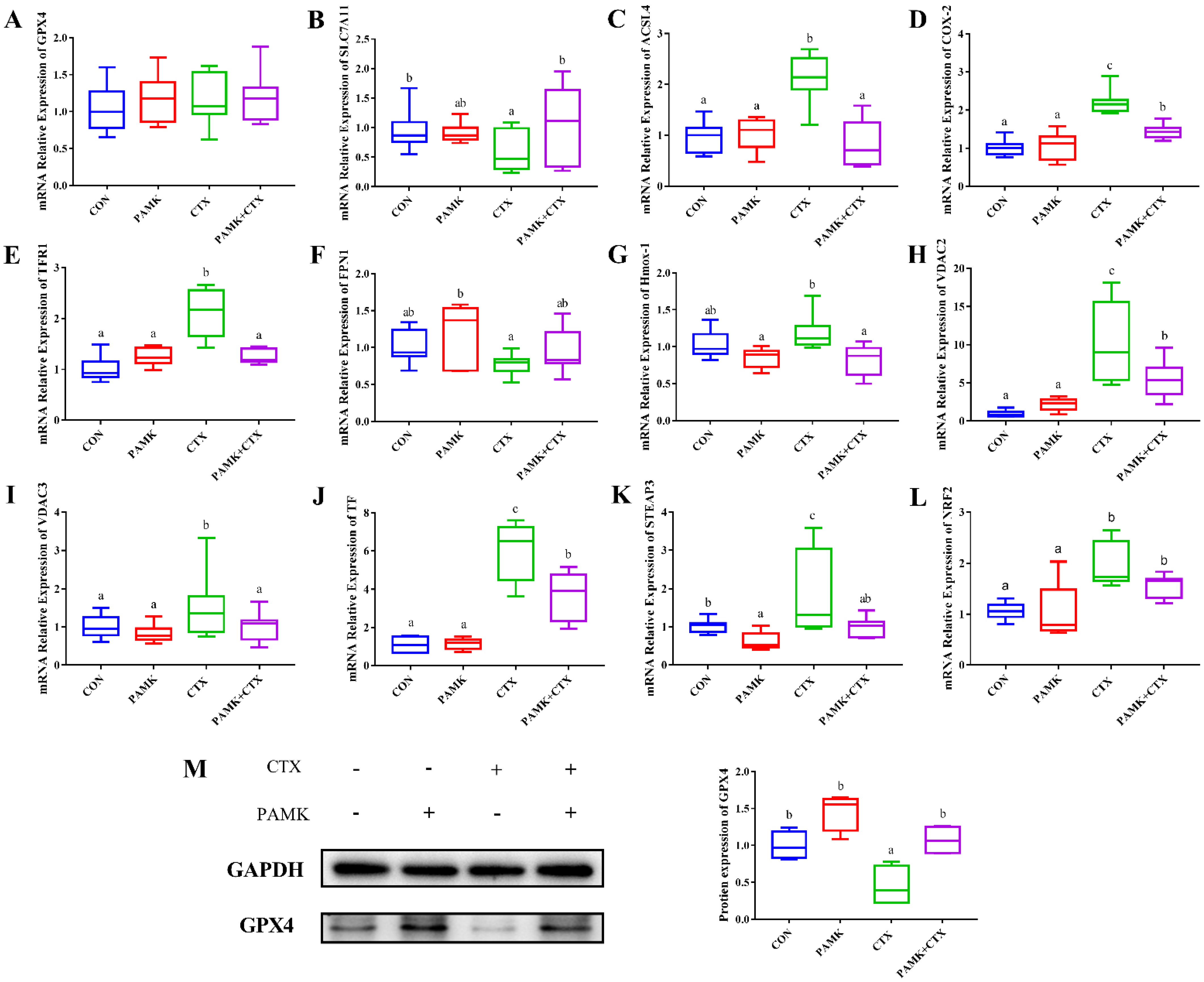
3.3. PAMK Alleviated the Decline of GPX4 in the Thymus
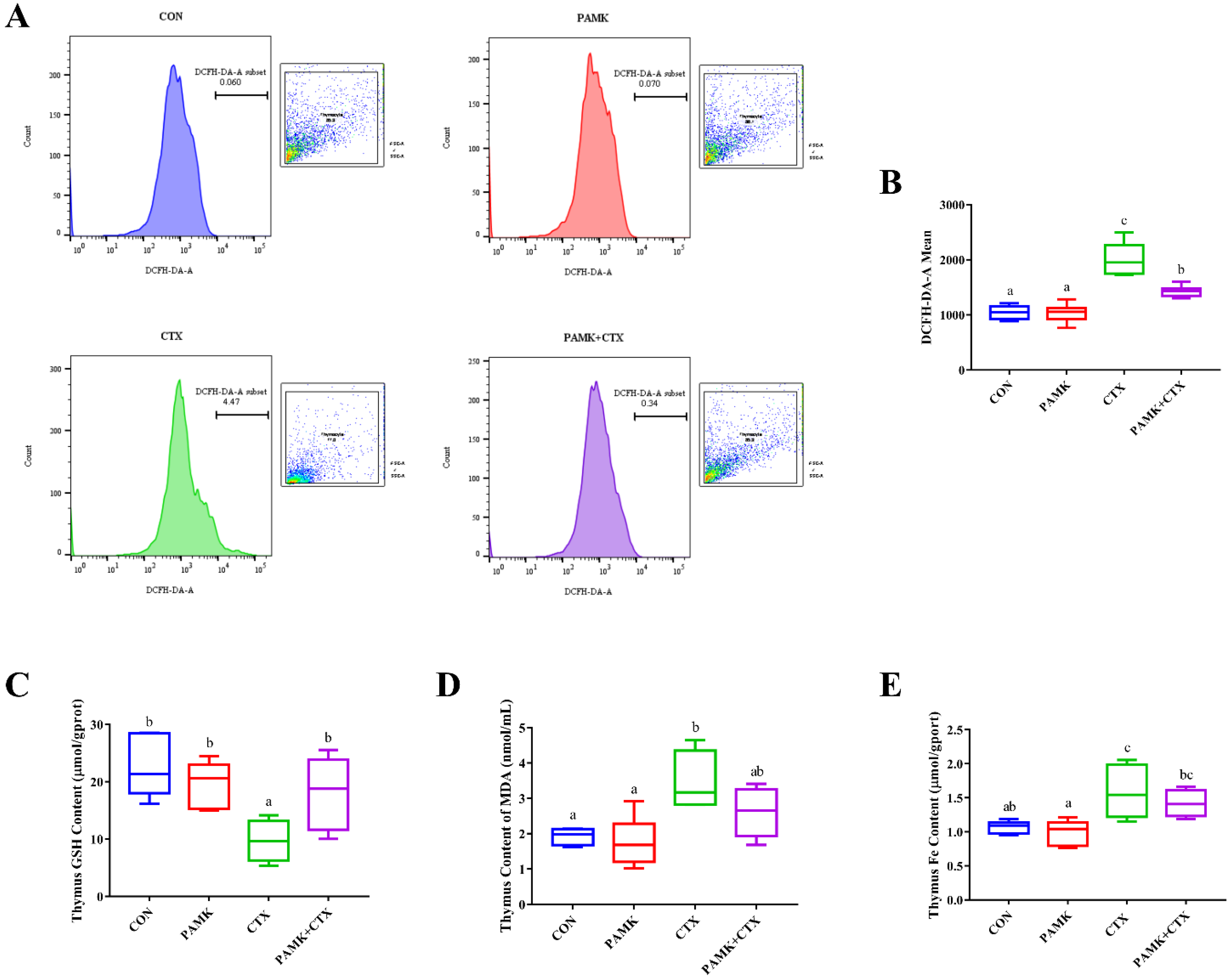
3.4. PAMK Alleviated CTX-Induced Elevated Levels of Oxidative Stress and Iron in the Thymus
3.5. PAMK Blocked the CTX-Induced Activation of Ferroptosis Pathway in the Thymus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PAMK | polysaccharide of Atractylodes macrocephala koidz |
| CTX | cyclophosphamide |
| ROS | reactive oxygen species |
| GSH | glutatione |
| MDA | malonedialdehyde |
| SEM | scanning electronic microscope |
| TEM | transmission electron microscope |
| HE | hematoxylin-eosin |
| PBS | phosphate buffer saline |
| BSA | bovine serum albumin |
| DAPI | 4′,6-diamidino-2-phenylindole |
| AA | arachidonic acid |
| GPX4 | glutathione peroxidase 4 |
| ACSL4 | Acyl-CoA synthetase long chain family member 4 |
| TFR1 | transferrin receptor 1 |
| FPN1 | ferroportin 1 |
| FTH1 | ferritin heavy chain 1 |
| Hmox-1 | heme oxygenase 1 |
| COX-2 | cyclooxygenase 2 |
| TF | transferrin |
| VDAC2/3 | voltage dependent anion channel 2/3 |
| STEAP3 | six-transmembrane epithelial antigen of prostate 3 |
| NRF2 | nuclear factor erythroid2-related factor 2 |
References
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Youssef, L.A.; Rebbaa, A.; Pampou, S.; Weisberg, S.P.; Stockwell, B.R.; Hod, E.A.; Spitalnik, S.L. Increased erythrophagocytosis induces ferroptosis in red pulp macrophages in a mouse model of transfusion. Blood 2018, 131, 2581–2593. [Google Scholar] [CrossRef] [PubMed]
- Drijvers, J.M.; Gillis, J.E.; Muijlwijk, T.; Nguyen, T.H.; Gaudiano, E.F.; Harris, I.S.; LaFleur, M.W.; Ringel, A.E.; Yao, C.-H.; Kurmi, K.; et al. Pharmacologic Screening Identifies Metabolic Vulnerabilities of CD8+ T Cells. Cancer Immunol. Res. 2021, 9, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Bai, L.; Qu, C.; Dai, E.; Liu, J.; Kang, R.; Zhou, D.; Tang, D.; Zhao, Y. PPARG-mediated ferroptosis in dendritic cells limits antitumor immunity. Biochem. Biophys. Res. Commun. 2021, 576, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Yunis, E.J.; Stutman, O.; Good, R.A. Thymus, immunity and autoimmunity. Ann. N. Y. Acad. Sci. 2010, 183, 205–220. [Google Scholar] [CrossRef]
- Duncan, I.J.H. Science-based assessment of animal welfare: Farm animals. Rev. Sci. Et Tech. 2005, 24, 483–492. [Google Scholar] [CrossRef]
- Wanas, H.; El-Shabrawy, M.; Mishriki, A.; Attia, H.; Emam, M.; Aboulhoda, B.E. Nebivolol protects against cyclophosphamide-induced nephrotoxicity through modulation of oxidative stress, inflammation, and apoptosis. Clin. Exp. Pharmacol. Physiol. 2021, 48, 811–819. [Google Scholar] [CrossRef]
- Singh, C.; Prakash, C.; Tiwari, K.N.; Mishra, S.K.; Kumar, V. Premna integrifolia ameliorates cyclophosphamide-induced hepatotoxicity by modulation of oxidative stress and apoptosis. Biomed. Pharmacother. 2018, 107, 634–643. [Google Scholar] [CrossRef]
- Ypsilantis, P.; Tentes, L.; Assimakopoulos, S.F.; Kortsaris, A.; Scopa, C.D.; Pitiakoudis, M.; Simopoulos, C. Dose related effects of ifosfamide on enterocyte apoptosis in different sites of the rabbit intestine. Toxicology 2004, 200, 135–143. [Google Scholar] [CrossRef]
- Shi, H.; Hou, B.; Li, H.; Zhou, H.; Du, B. Cyclophosphamide Induces the Ferroptosis of Tumor Cells through Heme Oxygenase-1. Front. Pharmacol. 2022, 13, 839464. [Google Scholar] [CrossRef]
- Xiang, X.; Cao, N.; Chen, F.; Qian, L.; Wang, Y.; Huang, Y.; Tian, Y.; Xu, D.; Li, W. Polysaccharide of Atractylodes macrocephala Koidz (PAMK) Alleviates Cyclophosphamide-induced Immunosuppression in Mice by Upregulating CD28/IP3R/PLCγ-1/AP-1/NFAT Signal Pathway. Front. Pharmacol. 2020, 11, 529657. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Li, W.; Chen, F.; Yang, S.; Huang, Y.; Tian, Y.; Xu, D.; Cao, N. Polysaccharide of Atractylodes macrocephala Koidz regulates LPS-mediated mouse hepatitis through the TLR4-MyD88-NFκB signaling pathway. Int. Immunopharmacol. 2021, 98, 107692. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Guo, S.; Xu, D.; Li, B.; Cao, N.; Tian, Y.; Jiang, Q. Polysaccharide of Atractylodes macrocephala Koidz (PAMK) Relieves Immunosuppression in Cyclophosphamide-Treated Geese by Maintaining a Humoral and Cellular Immune Balance. Molecules 2018, 23, 932. [Google Scholar] [CrossRef]
- Li, W.; Xiang, X.; Cao, N.; Chen, W.; Tian, Y.; Zhang, X.; Shen, X.; Jiang, D.; Xu, D.; Xu, S. Polysaccharide of Atractylodes macrocephala koidz activated T lymphocytes to alleviate cyclophosphamide-induced immunosuppression of geese through novel_mir2/CD28/AP-1 signal pathway. Poult. Sci. 2021, 100, 101129. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Li, B.; Cao, N.; Li, W.; Tian, Y.; Huang, Y. The protective effects of polysaccharide of Atractylodes macrocephala Koidz (PAMK) on the chicken spleen under heat stress via antagonizing apoptosis and restoring the immune function. Oncotarget 2017, 8, 70394–70405. [Google Scholar] [CrossRef] [PubMed]
- Emadi, A.; Jones, R.J.; Brodsky, R.A. Cyclophosphamide and cancer: Golden anniversary. Nat. Rev. Clin. Oncol. 2009, 6, 638–647. [Google Scholar] [CrossRef]
- Ibrahim, H.M.; Mohammed-Geba, K.; Tawfic, A.A.; El-Magd, M.A. Camel milk exosomes modulate cyclophosphamide-induced oxidative stress and immuno-toxicity in rats. Food Funct. 2019, 10, 7523–7532. [Google Scholar] [CrossRef]
- Bai, R.-B.; Zhang, Y.-J.; Fan, J.-M.; Jia, X.-S.; Li, D.; Wang, Y.-P.; Zhou, J.; Yan, Q.; Hu, F.-D. Immune-enhancement effects of oligosaccharides from Codonopsis pilosula on cyclophosphamide induced immunosuppression in mice. Food Funct. 2020, 11, 3306–3315. [Google Scholar] [CrossRef]
- Miao, W.; He, L.; Zhang, Y.; Zhu, X.; Jiang, Y.; Liu, P.; Zhang, T.; Li, C. Ferroptosis is partially responsible for dexamethasone-induced T cell ablation, but not osteoporosis in larval zebrafish. Ecotoxicol. Environ. Saf. 2022, 242, 113872. [Google Scholar] [CrossRef]
- Li, Q.; Li, Q.-Q.; Jia, J.-N.; Sun, Q.-Y.; Zhou, H.-H.; Jin, W.-L.; Mao, X.-Y. Baicalein Exerts Neuroprotective Effects in FeCl3-Induced Posttraumatic Epileptic Seizures via Suppressing Ferroptosis. Front. Pharmacol. 2019, 10, 638. [Google Scholar] [CrossRef]
- Guan, X.; Li, Z.; Zhu, S.; Cheng, M.; Ju, Y.; Ren, L.; Yang, G.; Min, D. Galangin attenuated cerebral ischemia-reperfusion injury by inhibition of ferroptosis through activating the SLC7A11/GPX4 axis in gerbils. Life Sci. 2021, 264, 118660. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Zhang, R. Icariin enhances cell survival in lipopolysaccharide-induced synoviocytes by suppressing ferroptosis via the Xc-/GPX4 axis. Exp. Ther. Med. 2021, 21, 72. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Xiang, X.; Li, B.; Wang, Y.; Qian, L.; Tian, Y.; Huang, Y.; Xu, D.; Cao, N. PAMK Relieves LPS-Induced Enteritis and Improves Intestinal Flora Disorder in Goslings. Evid.-Based Complement. Altern. Med. 2021, 2021, 9721316–9721353. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhou, X.; Xu, S.; Cao, N.; Li, B.; Chen, W.; Yang, B.; Yuan, M.; Xu, D. Lipopolysaccharide-induced splenic ferroptosis in goslings was alleviated by polysaccharide of Atractylodes macrocephala koidz associated with proinflammatory factors. Poult. Sci. 2022, 101, 101725. [Google Scholar] [CrossRef] [PubMed]
- Cengiz, M.; Yildiz, S.C.; Demir, C.; Şahin, I.K.; Teksoy, Ö.; Ayhanci, A. Hepato-preventive and anti-apoptotic role of boric acid against liver injury induced by cyclophosphamide. J. Trace Elements Med. Biol. 2019, 53, 1–7. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Kalimuthu, K.; Park, Y.S.; Luo, X.; Choudry, M.H.A.; Bartlett, D.L.; Lee, Y.J. BAX-dependent mitochondrial pathway mediates the crosstalk between ferroptosis and apoptosis. Apoptosis 2020, 25, 625–631. [Google Scholar] [CrossRef]
- Pandrangi, S.L.; Chittineedi, P.; Chalumuri, S.S.; Meena, A.S.; Mosquera, J.A.N.; Llaguno, S.N.S.; Pamuru, R.R.; Mohiddin, G.J.; Mohammad, A. Role of Intracellular Iron in Switching Apoptosis to Ferroptosis to Target Therapy-Resistant Cancer Stem Cells. Molecules 2022, 27, 3011. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, D.; Hong, M.; Liu, J.; Li, Y.; Hao, J.; Lu, L.; Yin, Z.; Wu, Y. Apoptosis, Pyroptosis, and Ferroptosis Conspiringly Induce Immunosuppressive Hepatocellular Carcinoma Microenvironment and γδ T-Cell Imbalance. Front. Immunol. 2022, 13, 845974. [Google Scholar] [CrossRef]
- Fu, Y.-P.; Feng, B.; Zhu, Z.-K.; Feng, X.; Chen, S.-F.; Li, L.-X.; Yin, Z.-Q.; Huang, C.; Chen, X.-F.; Zhang, B.-Z.; et al. The Polysaccharides from Codonopsis pilosula Modulates the Immunity and Intestinal Microbiota of Cyclophosphamide-Treated Immunosuppressed Mice. Molecules 2018, 23, 1801. [Google Scholar] [CrossRef]
- Meng, M.; Guo, M.; Feng, C.; Wang, R.; Cheng, D.; Wang, C. Water-soluble polysaccharides from Grifola Frondosa fruiting bodies protect against immunosuppression in cyclophosphamide-induced mice via JAK2/STAT3/SOCS signal transduction pathways. Food Funct. 2019, 10, 4998–5007. [Google Scholar] [CrossRef]
- Bai, Y.-T.; Chang, R.; Wang, H.; Xiao, F.-J.; Ge, R.-L.; Wang, L.-S. ENPP2 protects cardiomyocytes from erastin-induced ferroptosis. Biochem. Biophys. Res. Commun. 2018, 499, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Friedmann Angeli, J.P.; Schneider, M.; Proneth, B.; Tyurina, Y.Y.; Tyurin, V.A.; Hammond, V.J.; Herbach, N.; Aichler, M.; Walch, A.; Eggenhofer, E.; et al. Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nat. Cell Biol. 2014, 16, 1180–1191. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of Ferroptotic Cancer Cell Death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Freigang, S.; Schneider, C.; Conrad, M.; Bornkamm, G.W.; Kopf, M. T cell lipid peroxidation induces ferroptosis and prevents immunity to infection. J. Exp. Med. 2015, 212, 555–568. [Google Scholar] [CrossRef]
- Watanabe, N.; Wang, Y.-H.; Lee, H.K.; Ito, T.; Wang, Y.-H.; Cao, W.; Liu, Y.-J. Hassall’s corpuscles instruct dendritic cells to induce CD4+CD25+ regulatory T cells in human thymus. Nature 2005, 436, 1181–1185. [Google Scholar] [CrossRef]
- Ganz, T. Iron in innate immunity: Starve the invaders. Curr. Opin. Immunol. 2009, 21, 63–67. [Google Scholar] [CrossRef]
- Nairz, M.; Schroll, A.; Sonnweber, T.; Weiss, G. The struggle for iron—A metal at the host-pathogen interface. Cell. Microbiol. 2010, 12, 1691–1702. [Google Scholar] [CrossRef]
- Grenier, D.; Goulet, V.; Mayrand, D. The Capacity of Porphyromonas gingivalis to Multiply Under Iron-limiting Conditions Correlates with its Pathogenicity in an Animal Model. J. Dent. Res. 2001, 80, 1678–1682. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, J.; Guo, W.; Li, H.; Lei, L. Periodontitis-level butyrate-induced ferroptosis in periodontal ligament fibroblasts by activation of ferritinophagy. Cell Death Discov. 2020, 6, 119. [Google Scholar] [CrossRef]
- Sazawal, S.; Black, R.E.; Ramsan, M.; Chwaya, H.M.; Stoltzfus, R.J.; Dutta, A.; Dhingra, U.; Kabole, I.; Deb, S.; Othman, M.K.; et al. Effects of routine prophylactic supplementation with iron and folic acid on admission to hospital and mortality in preschool children in a high malaria transmission setting: Community-based, randomised, placebo-controlled trial. Lancet 2006, 367, 133–143. [Google Scholar] [CrossRef]
- Liu, L.; Liu, B.; Guan, G.; Kang, R.; Dai, Y.; Tang, D. Cyclophosphamide-induced GPX4 degradation triggers parthanatos by activating AIFM1. Biochem. Biophys. Res. Commun. 2022, 606, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Yan, R.; Zhu, J.; Cheng, S.; Kong, C.; Chen, W.; Fang, S.; Wang, Y.; Yang, Y.; Qiu, R.; et al. Integrative analysis of the molecular mechanisms, immunological features and immunotherapy response of ferroptosis regulators across 33 cancer types. Int. J. Biol. Sci. 2022, 18, 180–198. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; An, P.; Xie, E.; Wu, Q.; Fang, X.; Gao, H.; Zhang, Z.; Li, Y.; Wang, X.; Zhang, J.; et al. Characterization of ferroptosis in murine models of hemochromatosis. Hepatology 2017, 66, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Karuppagounder, S.S.; Alin, L.; Chen, Y.; Brand, D.; Bourassa, M.W.; Bs, K.D.; Bs, C.M.W.; Ms, C.A.N.; Kumar, A.; Perry, S.; et al. N-acetylcysteine targets 5 lipoxygenase-derived, toxic lipids and can synergize with prostaglandin E2 to inhibit ferroptosis and improve outcomes following hemorrhagic stroke in mice. Ann. Neurol. 2018, 84, 854–872. [Google Scholar] [CrossRef] [PubMed]
- Maloberti, P.M.; Duarte, A.B.; Orlando, U.D.; Pasqualini, M.E.; Solano, A.R.; López-Otín, C.; Podesta, E.J. Functional Interaction between Acyl-CoA Synthetase 4, Lipooxygenases and Cyclooxygenase-2 in the Aggressive Phenotype of Breast Cancer Cells. PLoS ONE 2010, 5, e15540. [Google Scholar] [CrossRef]
- Chiang, S.-K.; Chen, S.-E.; Chang, L.-C. The Role of HO-1 and Its Crosstalk with Oxidative Stress in Cancer Cell Survival. Cells 2021, 10, 2401. [Google Scholar] [CrossRef]
- Chiang, S.-K.; Chen, S.-E.; Chang, L.-C. A Dual Role of Heme Oxygenase-1 in Cancer Cells. Int. J. Mol. Sci. 2018, 20, 39. [Google Scholar] [CrossRef]
- Gao, G.; Li, J.; Zhang, Y.; Chang, Y.-Z. Cellular Iron Metabolism and Regulation. Adv. Exp. Med. Biol. 2019, 1173, 21–32. [Google Scholar] [CrossRef]
- Mou, Y.; Wang, J.; Wu, J.; He, D.; Zhang, C.; Duan, C.; Li, B. Ferroptosis, a new form of cell death: Opportunities and challenges in cancer. J. Hematol. Oncol. 2019, 12, 34. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, F.; An, P.; Guo, X.; Shen, Y.; Tao, Y.; Wu, Q.; Zhang, Y.; Yu, Y.; Ning, B.; et al. Ferroportin1 deficiency in mouse macrophages impairs iron homeostasis and inflammatory responses. Blood 2011, 118, 1912–1922. [Google Scholar] [CrossRef]
- DeHart, D.N.; Fang, D.; Heslop, K.; Li, L.; Lemasters, J.J.; Maldonado, E.N. Opening of voltage dependent anion channels promotes reactive oxygen species generation, mitochondrial dysfunction and cell death in cancer cells. Biochem. Pharmacol. 2018, 148, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Lipper, C.H.; Stofleth, J.T.; Bai, F.; Sohn, Y.-S.; Roy, S.; Mittler, R.; Nechushtai, R.; Onuchic, J.N.; Jennings, P.A. Redox-dependent gating of VDAC by mitoNEET. Proc. Natl. Acad. Sci. USA 2019, 116, 19924–19929. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer (5′ → 3′) | Gen Bank Access |
|---|---|---|
| β-actin-F | GCACCCAGCACGATGAAAAT | XM_013174886.1 |
| β-actin-R | GACAATGGAGGGTCCGGATT | |
| GPX4-F | TCGATGTGAATGGGGACGAC | XM_013200057.1 |
| GPX4-F | GTCCTTCTCGATGACGTAGGG | |
| ACSL4-F | GCGGCTGAAACCCTCTTCTT | XM_013185083.1 |
| ACSL4-R | GCCAACAGTGGACACAAGCTA | |
| TFR1-F | AGAATGGCTGGAGGGGTACT | XM_013195023.1 |
| TFR1-R | TTCTCTCCAGCAGCGCATAC | |
| FTH1-F | ATGGTCATGGGCTTTCCCC | XM_013177583.1 |
| FTH1-R | AATGAAGTCACACAGATGCGG | |
| FPN1-F | CTGGGGAGATCGTATGTGGC | XM_013178636.1 |
| FPN1-R | AGGATGTCTGGGCCACTTTG | |
| Hmox-1-F | ATATGAGCACGGTCCAGCG | XM_013181078.2 |
| Hmox-1-R | TCGTGACTATGAAGCCGAGC | |
| COX-2-F | TGTCCTTTCACTGCTTTCCAT | XM_013177944.1 |
| COX-2-R | TTCCATTGCTGTGTTTGAGGT | |
| TF-F | ATTACTTCAGTGCGGGCTGT | XM_013186329.2 |
| TF-R | CTCGACCAGACACCGGAAA | |
| VDAC2-F | GGAAGCTGCAACACGAAGAAC | XM_013176155.2 |
| VDAC2-R | ACCAACCCAAACCCATATCCT | |
| VDAC3-F | CCAGTGGGGTGCTGGAATTTA | XM_013194404.2 |
| VDAC3-R | TCCCAATGTGTTGTCCGTGT | |
| STEAP3-F | CCGTCAAGCAGTCCACCCT | XM_048079666.1 |
| STEAP3-R | ACAGTACATGGGACGAGCAG | |
| NRF2-F | GGGATGCCCGGACATGAA | XM_013171581.2 |
| NRF2-R | CGTCTAACTCCAGCTGAGCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Cao, N.; Xu, D.; Tian, Y.; Shen, X.; Jiang, D.; Huang, Y.; Li, W.; Li, B. Polysaccharide of Atractylodes macrocephala Koidz Alleviates Cyclophosphamide-Induced Thymus Ferroptosis in Gosling. Animals 2022, 12, 3394. https://doi.org/10.3390/ani12233394
Zhou X, Cao N, Xu D, Tian Y, Shen X, Jiang D, Huang Y, Li W, Li B. Polysaccharide of Atractylodes macrocephala Koidz Alleviates Cyclophosphamide-Induced Thymus Ferroptosis in Gosling. Animals. 2022; 12(23):3394. https://doi.org/10.3390/ani12233394
Chicago/Turabian StyleZhou, Xiangying, Nan Cao, Danning Xu, Yunbo Tian, Xu Shen, Danli Jiang, Yunmao Huang, Wanyan Li, and Bingxin Li. 2022. "Polysaccharide of Atractylodes macrocephala Koidz Alleviates Cyclophosphamide-Induced Thymus Ferroptosis in Gosling" Animals 12, no. 23: 3394. https://doi.org/10.3390/ani12233394
APA StyleZhou, X., Cao, N., Xu, D., Tian, Y., Shen, X., Jiang, D., Huang, Y., Li, W., & Li, B. (2022). Polysaccharide of Atractylodes macrocephala Koidz Alleviates Cyclophosphamide-Induced Thymus Ferroptosis in Gosling. Animals, 12(23), 3394. https://doi.org/10.3390/ani12233394