

Fecal Microbiome Features Associated with Extended-Spectrum β-Lactamase-Producing Enterobacterales Carriage in Dairy Heifers
 ,
,  , ,
, ,  ,
,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Demographic and Environmental Data Collection
2.2. Fecal Sample Collection
2.3. ESBL-Producing Enterobacterales Screening
2.4. Samples for Microbiome Study
2.5. DNA Extraction and 16S rRNA Gene Sequencing
2.6. Sequence Data Processing
2.7. Taxonomic, Diversity, and Statistical Analysis
3. Results
3.1. General Characterization
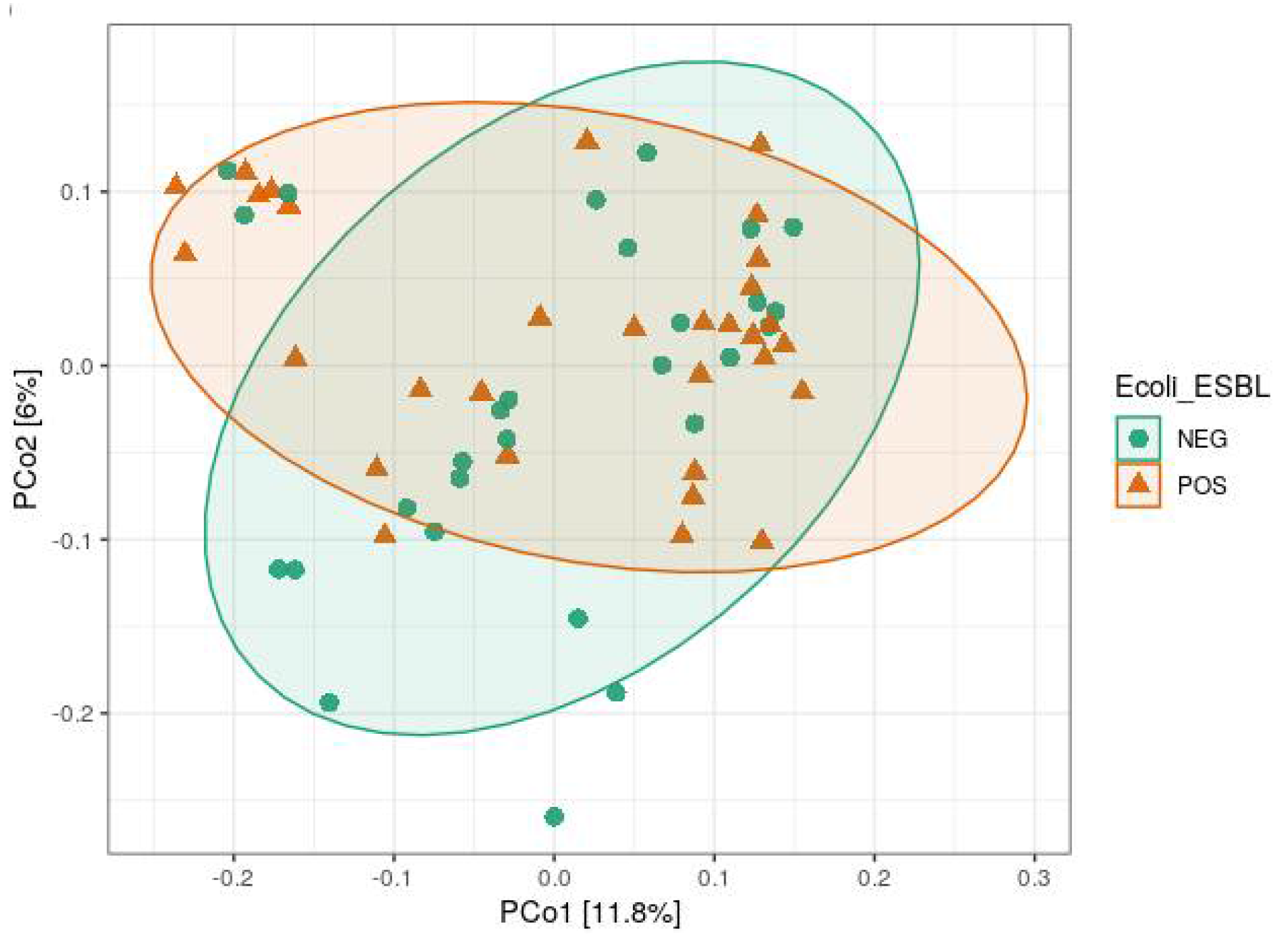
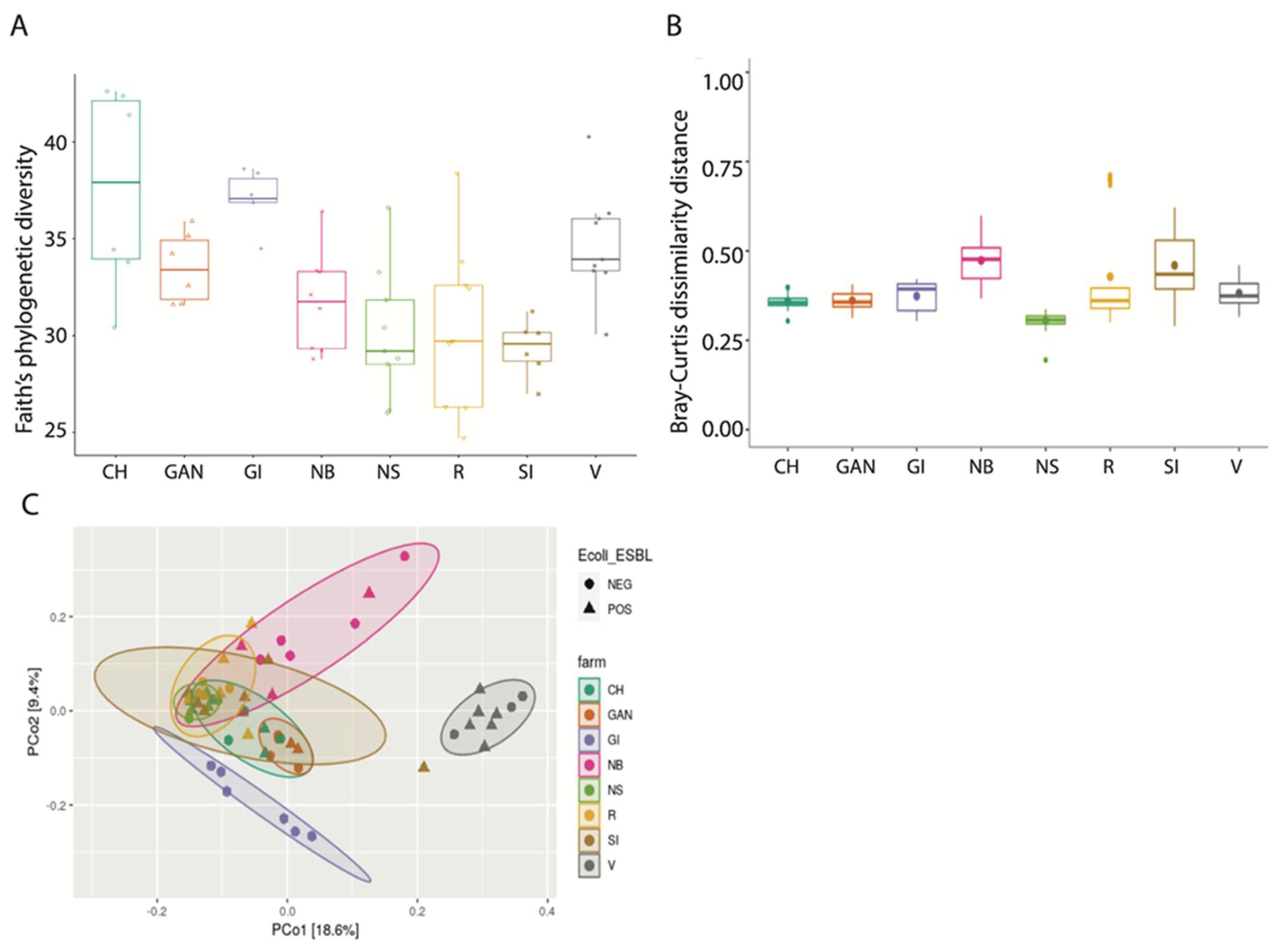
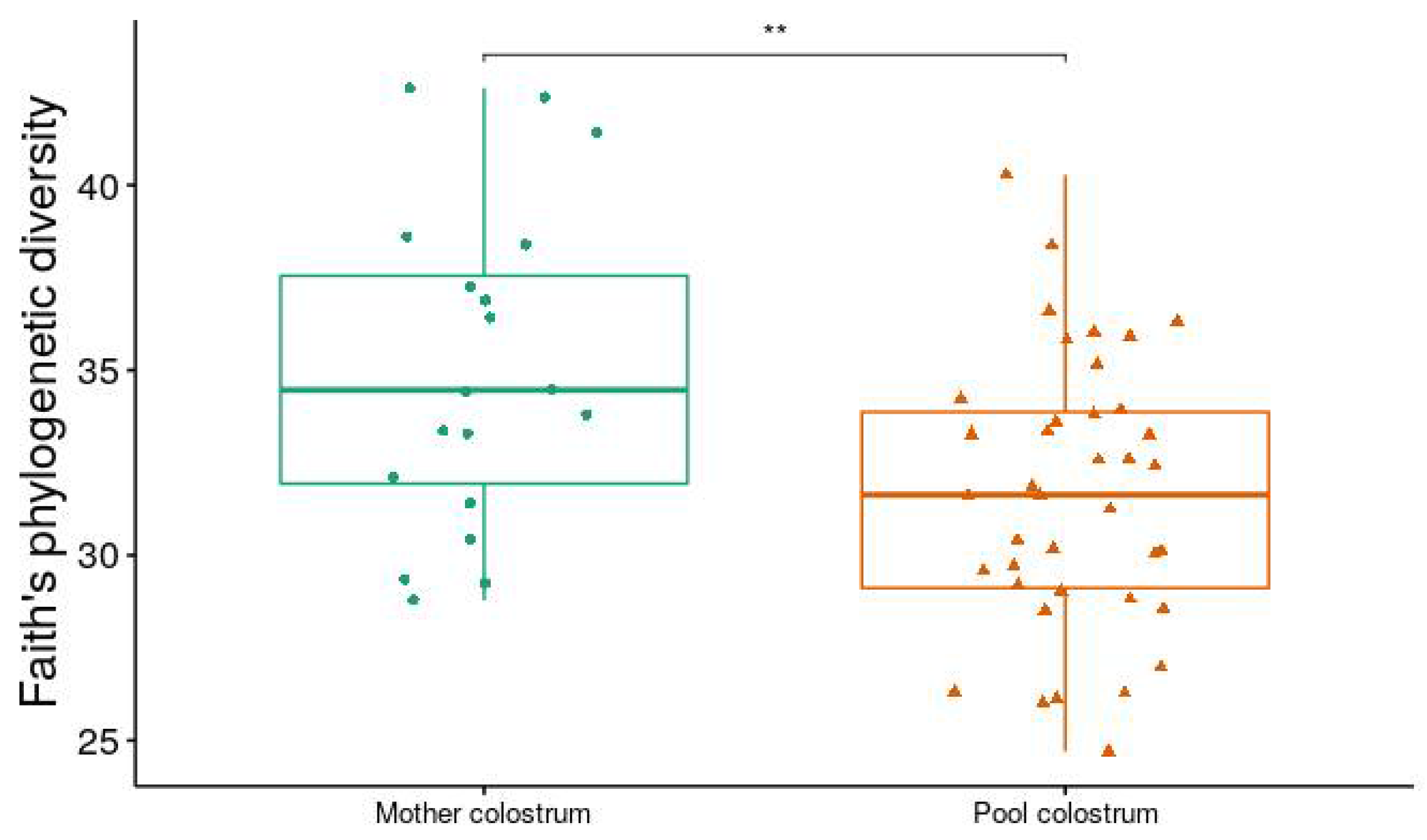
3.2. Alpha and Beta Diversity Analysis
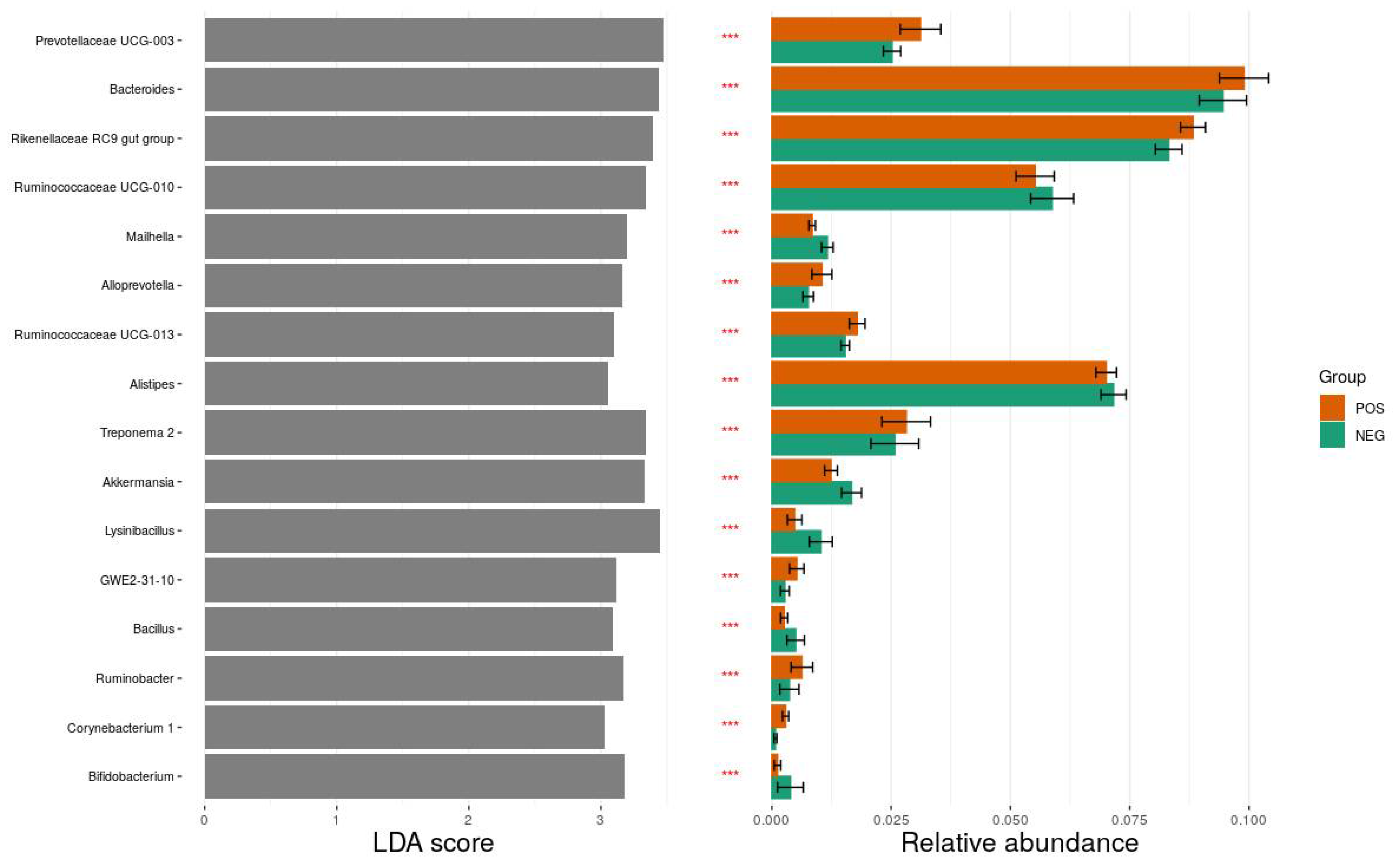
3.3. Taxonomic Composition of Fecal Microbiota between ESBL-PE Carriers and Noncarriers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dantas Palmeira, J.; Ferreira, H.M.N. Extended-Spectrum Beta-Lactamase (ESBL)-Producing Enterobacteriaceae in Cattle Production—A Threat around the World. Heliyon 2020, 6, e03206. [Google Scholar] [CrossRef]
- Dahms, C.; Hübner, N.-O.; Kossow, A.; Mellmann, A.; Dittmann, K.; Kramer, A. Occurrence of ESBL-Producing Escherichia Coli in Livestock and Farm Workers in Mecklenburg-Western Pomerania, Germany. PLoS ONE 2015, 10, e0143326. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Mir, R.A.; Park, S.H.; Kim, D.; Kim, H.-Y.; Boughton, R.K.; Morris, J.G., Jr.; Jeong, K.C. Prevalence of Extended-Spectrum β-Lactamases in the Local Farm Environment and Livestock: Challenges to Mitigate Antimicrobial Resistance. Crit. Rev. Microbiol. 2020, 46, 1–14. [Google Scholar] [CrossRef] [PubMed]
- World Agriculture towards 2030/2050: The 2012 Revision. 154. Available online: https://ageconsearch.umn.edu/record/288998/ (accessed on 15 June 2022).
- Adler, A.; Sturlesi, N.; Fallach, N.; Zilberman-Barzilai, D.; Hussein, O.; Blum, S.E.; Klement, E.; Schwaber, M.J.; Carmeli, Y. Prevalence, Risk Factors, and Transmission Dynamics of Extended-Spectrum-β-Lactamase-Producing Enterobacteriaceae: A National Survey of Cattle Farms in Israel in 2013. J. Clin. Microbiol. 2015, 53, 3515–3521. [Google Scholar] [CrossRef] [PubMed]
- Geser, N.; Stephan, R.; Kuhnert, P.; Zbinden, R.; Kaeppeli, U.; Cernela, N.; Haechler, H. Fecal Carriage of Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae in Swine and Cattle at Slaughter in Switzerland. J. Food Prot. 2011, 74, 446–449. [Google Scholar] [CrossRef]
- Chen, C.-M.; Ke, S.-C.; Li, C.-R.; Wu, Y.-C.; Chen, T.-H.; Lai, C.-H.; Wu, X.-X.; Wu, L.-T. High Diversity of Antimicrobial Resistance Genes, Class 1 Integrons, and Genotypes of Multidrug-Resistant Escherichia Coli in Beef Carcasses. Microb. Drug Resist. 2017, 23, 915–924. [Google Scholar] [CrossRef]
- Mir, R.A.; Weppelmann, T.A.; Teng, L.; Kirpich, A.; Elzo, M.A.; Driver, J.D.; Jeong, K.C. Colonization Dynamics of Cefotaxime Resistant Bacteria in Beef Cattle Raised Without Cephalosporin Antibiotics. Front. Microbiol. 2018, 9, 500. [Google Scholar] [CrossRef]
- Costa, M.C.; Weese, J.S. Understanding the Intestinal Microbiome in Health and Disease. Vet. Clin. N. Am. Equine Pract. 2018, 34, 1–12. [Google Scholar] [CrossRef]
- O’Hara, E.; Neves, A.L.A.; Song, Y.; Guan, L.L. The Role of the Gut Microbiome in Cattle Production and Health: Driver or Passenger? Annu. Rev. Anim. Biosci. 2020, 8, 199–220. [Google Scholar] [CrossRef]
- Kim, M.; Wells, J.E. A Meta-Analysis of Bacterial Diversity in the Feces of Cattle. Curr. Microbiol. 2016, 72, 145–151. [Google Scholar] [CrossRef]
- Gosalbes, M.; Vázquez-Castellanos, J.; Angebault, C.; Woerther, P.-L.; Ruppé, E.; Ferrús, M.; Latorre, A.; Andremont, A.; Moya, A. Carriage of Enterobacteria Producing Extended Spectrum Beta-Lactamases and Composition of the Gut Microbiota in an Amerindian Community. Antimicrob. Agents Chemother. 2015, 60, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Pultz, N.J.; Stiefel, U.; Subramanyan, S.; Helfand, M.S.; Donskey, C.J. Mechanisms by Which Anaerobic Microbiota Inhibit the Establishment in Mice of Intestinal Colonization by Vancomycin-Resistant Enterococcus. J. Infect. Dis. 2005, 191, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.A.; Williams, A.G. Production and Management of Cattle Manure in the UK and Implications for Land Application Practice. Soil Use Manag. 2016, 32, 73–82. [Google Scholar] [CrossRef]
- Piewngam, P.; Quiñones, M.; Thirakittiwatthana, W.; Yungyuen, T.; Otto, M.; Kiratisin, P. Composition of the Intestinal Microbiota in Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae Carriers and Non-Carriers in Thailand. Int. J. Antimicrob. Agents 2019, 53, 435–441. [Google Scholar] [CrossRef]
- Le Bastard, Q.; Chapelet, G.; Birgand, G.; Hillmann, B.M.; Javaudin, F.; Hayatgheib, N.; Bourigault, C.; Bemer, P.; De Decker, L.; Batard, E.; et al. Gut Microbiome Signatures of Nursing Home Residents Carrying Enterobacteria Producing Extended-Spectrum β-Lactamases. Antimicrob. Resist. Infect. Control 2020, 9, 107. [Google Scholar] [CrossRef]
- Murk, J.-L.A.N.; Heddema, E.R.; Hess, D.L.J.; Bogaards, J.A.; Vandenbroucke-Grauls, C.M.J.E.; Debets-Ossenkopp, Y.J. Enrichment Broth Improved Detection of Extended-Spectrum-Beta-Lactamase-Producing Bacteria in Throat and Rectal Surveillance Cultures of Samples from Patients in Intensive Care Units. J. Clin. Microbiol. 2009, 47, 1885–1887. [Google Scholar] [CrossRef]
- Poulou, A.; Grivakou, E.; Vrioni, G.; Koumaki, V.; Pittaras, T.; Pournaras, S.; Tsakris, A. Modified CLSI Extended-Spectrum β-Lactamase (ESBL) Confirmatory Test for Phenotypic Detection of ESBLs among Enterobacteriaceae Producing Various β-Lactamases. J. Clin. Microbiol. 2014, 52, 1483–1489. [Google Scholar] [CrossRef]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF Mass Spectrometry: An Emerging Technology for Microbial Identification and Diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-High-Throughput Microbial Community Analysis on the Illumina HiSeq and MiSeq Platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Liu, C.; Yaoming, C.; Xiangzhen, L.; Minjie, Y. microeco: An R package for data mining in microbial community ecology. FEMS Microbiol. Ecol. 2021, 97, 2. [Google Scholar] [CrossRef] [PubMed]
- Reist, M.; Geser, N.; Hächler, H.; Schärrer, S.; Stephan, R. ESBL-Producing Enterobacteriaceae: Occurrence, Risk Factors for Fecal Carriage and Strain Traits in the Swiss Slaughter Cattle Population Younger than 2 Years Sampled at Abattoir Level. PLoS ONE 2013, 8, e71725. [Google Scholar] [CrossRef] [PubMed]
- Cendron, F.; Niero, G.; Carlino, G.; Penasa, M.; Cassandro, M. Characterizing the Fecal Bacteria and Archaea Community of Heifers and Lactating Cows through 16S RRNA Next-Generation Sequencing. J. Appl. Genet. 2020, 61, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Myer, P.R.; Freetly, H.C.; Wells, J.E.; Smith, T.P.L.; Kuehn, L.A. Analysis of the Gut Bacterial Communities in Beef Cattle and Their Association with Feed Intake, Growth, and Efficiency1,2,3. J. Anim. Sci. 2017, 95, 3215–3224. [Google Scholar] [CrossRef][Green Version]
- Liu, J.; Taft, D.H.; Maldonado-Gomez, M.X.; Johnson, D.; Treiber, M.L.; Lemay, D.G.; DePeters, E.J.; Mills, D.A. The Fecal Resistome of Dairy Cattle Is Associated with Diet during Nursing. Nat. Commun. 2019, 10, 4406. [Google Scholar] [CrossRef]
- Furman, O.; Shenhav, L.; Sasson, G.; Kokou, F.; Honig, H.; Jacoby, S.; Hertz, T.; Cordero, O.X.; Halperin, E.; Mizrahi, I. Stochasticity Constrained by Deterministic Effects of Diet and Age Drive Rumen Microbiome Assembly Dynamics. Nat. Commun. 2020, 11, 1904. [Google Scholar] [CrossRef]
- Firth, C.L.; Kremer, K.; Werner, T.; Käsbohrer, A. The Effects of Feeding Waste Milk Containing Antimicrobial Residues on Dairy Calf Health. Pathogens 2021, 10, 112. [Google Scholar] [CrossRef]
- Xiong, W.; Sun, Y.; Zeng, Z. Antimicrobial Use and Antimicrobial Resistance in Food Animals. Env. Sci. Pollut. Res. 2018, 25, 18377–18384. [Google Scholar] [CrossRef]
- Yang, F.; Klopatek, S.; Oltjen, J.; Yang, X. Comparison of the Effects of Conventional Grain-Fed and Grass-Fed Feeding Systems on the Fecal Microbiota and STEC Population and Prevalence in California Beef Cattle. Ph.D. Dissertation, University of California Davis, Davis, CA, USA, 2020. Available online: https://www.proquest.com/openview/6ad5bfae10f1aa98d6e11394e6f36a97/1.pdf?pqorigsite=gscholar&cbl=18750&disy (accessed on 3 April 2022).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | N Positive (%) | OR (95% CI) | p-Value | ||
|---|---|---|---|---|---|
| Cooling system | Fan | 37 | 16 (43.2%) | 0.59 (0.39–0.88) | 0.01 |
| Nebulizers | 20 | 2 (10%) | 1.29 (0.60–2.79) | 0.506 | |
| Both | 100 | 37 (37%) | ref | ref | |
| Working dogs (yes/no) | No | 20 | 2 (10%) | ref | ref |
| Yes | 137 | 53 (38.7%) | 5.68 (1.27–25.47) | 0.023 | |
| Colostrum feeding of calves | Pooled colostrum | 97 | 48 (49.5%) | 7.42 (3.07–17.94) | <0.001 |
| Individual colostrum | 60 | 7 (11.7%) | ref | ref | |
| Milk feeding of calves | Milk replacement | 80 | 17 (21.3%) | 0.096 (0.04–0.24) | <0.001 |
| Whole milk | 39 | 10 (25.6%) | 0.12 (0.04–0.34) | <0.001 | |
| Both | 38 | 28 (73.7%) | ref | ref | |
| Method for manure cleaning | Tractor | 79 | 32 (40.5%) | 0.85 (0.30–2.39) | 0.76 |
| Automatic shovel | 60 | 15 (25%) | 0.42 (0.14–1.25) | 0.118 | |
| Both | 18 | 8 (44.4%) | ref | ref | |
| Antimicrobial prophylaxis | No | 117 | 33 (28.2%) | ref | ref |
| Yes | 40 | 22 (55%) | 3.11 (1.48–6.53) | 0.003 | |
| ESBL-PE-Negative | ESBL-PE-Positive | ||||||
|---|---|---|---|---|---|---|---|
| Phylum | N | Mean | SD | N | Mean | SD | p-Value |
| Bacteroidetes | 26 | 0.439 | 0.045 | 33 | 0.458 | 0.051 | 0.621 |
| Firmicutes | 26 | 0.397 | 0.04 | 33 | 0.386 | 0.045 | 0.801 |
| Spirochaetes | 26 | 0.029 | 0.03 | 33 | 0.034 | 0.036 | 0.986 |
| Proteobacteria | 26 | 0.03 | 0.013 | 33 | 0.029 | 0.017 | 0.801 |
| Tenericutes | 26 | 0.021 | 0.008 | 33 | 0.021 | 0.008 | 0.977 |
| Verrucomicrobia | 26 | 0.022 | 0.007 | 33 | 0.015 | 0.007 | 0.801 |
| Cyanobacteria | 26 | 0.017 | 0.011 | 33 | 0.013 | 0.007 | 0.621 |
| Kiritimatiellaeota | 26 | 0.014 | 0.006 | 33 | 0.011 | 0.006 | 0.977 |
| Actinobacteria | 26 | 0.012 | 0.01 | 33 | 0.005 | 0.009 | 0.977 |
| Lentisphaerae | 26 | 0.005 | 0.01 | 33 | 0.004 | 0.008 | 0.801 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cohen, A.; Poupko, L.; Craddock, H.A.; Motro, Y.; Khalfin, B.; Zelinger, A.; Tirosh-Levy, S.; Blum, S.E.; Steinman, A.; Moran-Gilad, J. Fecal Microbiome Features Associated with Extended-Spectrum β-Lactamase-Producing Enterobacterales Carriage in Dairy Heifers. Animals 2022, 12, 1738. https://doi.org/10.3390/ani12141738
Cohen A, Poupko L, Craddock HA, Motro Y, Khalfin B, Zelinger A, Tirosh-Levy S, Blum SE, Steinman A, Moran-Gilad J. Fecal Microbiome Features Associated with Extended-Spectrum β-Lactamase-Producing Enterobacterales Carriage in Dairy Heifers. Animals. 2022; 12(14):1738. https://doi.org/10.3390/ani12141738
Chicago/Turabian StyleCohen, Adar, Liat Poupko, Hillary A. Craddock, Yair Motro, Boris Khalfin, Amit Zelinger, Sharon Tirosh-Levy, Shlomo E. Blum, Amir Steinman, and Jacob Moran-Gilad. 2022. "Fecal Microbiome Features Associated with Extended-Spectrum β-Lactamase-Producing Enterobacterales Carriage in Dairy Heifers" Animals 12, no. 14: 1738. https://doi.org/10.3390/ani12141738
APA StyleCohen, A., Poupko, L., Craddock, H. A., Motro, Y., Khalfin, B., Zelinger, A., Tirosh-Levy, S., Blum, S. E., Steinman, A., & Moran-Gilad, J. (2022). Fecal Microbiome Features Associated with Extended-Spectrum β-Lactamase-Producing Enterobacterales Carriage in Dairy Heifers. Animals, 12(14), 1738. https://doi.org/10.3390/ani12141738