
Different Immune Responses of the Lymphoid Organ in Shrimp at Early Challenge Stage of Vibrio parahaemolyticus and WSSV


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Immune Challenge and Shrimp Tissues Preparation
2.2. RNA Extraction and Illumina Sequencing
2.3. De Novo Assembly and Transcriptome Analysis
2.4. Differential Expression Analysis and Functional Characterization
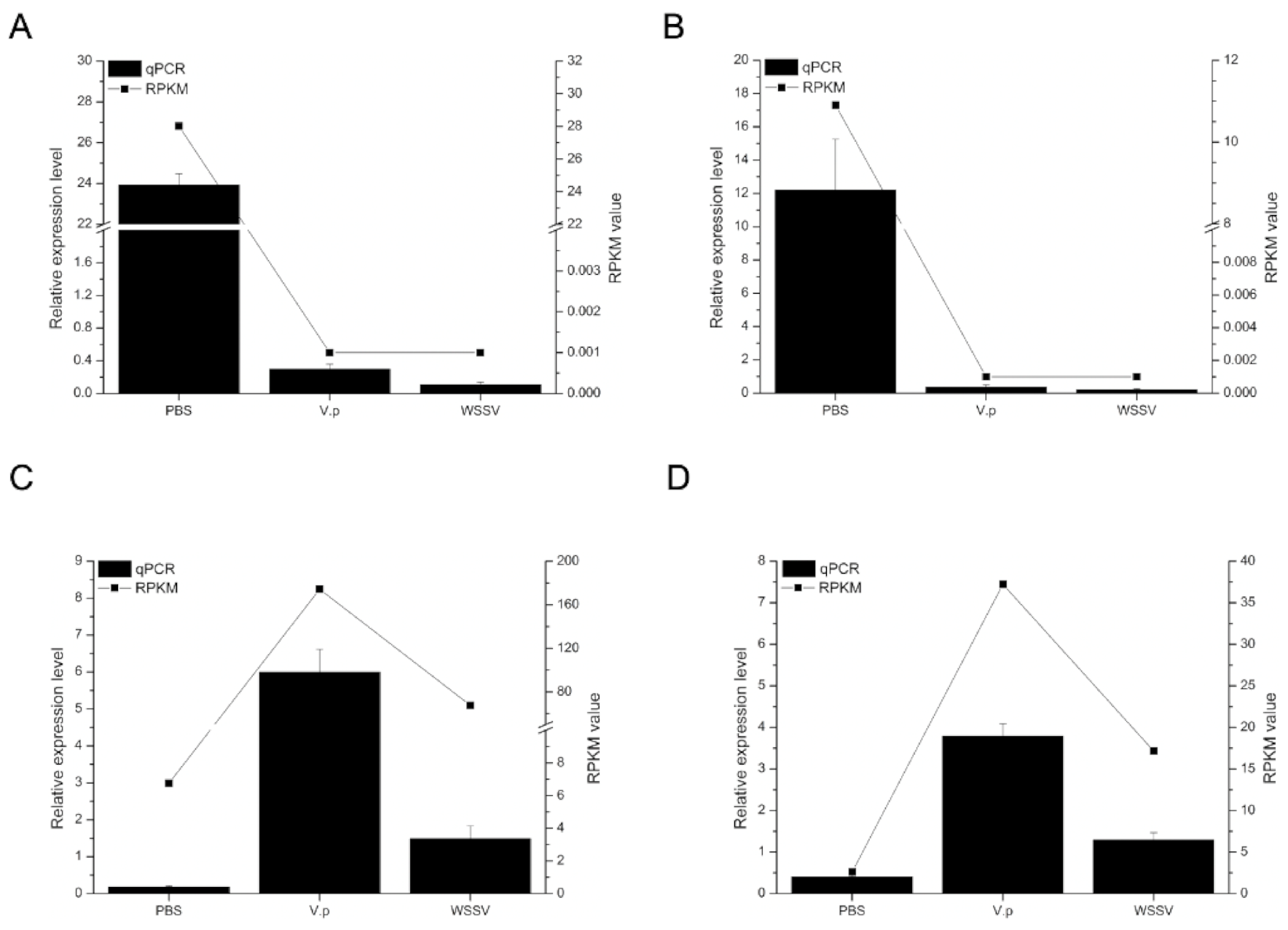
2.5. Data Reliability Confirmed by qRT-PCR Assay
3. Results and Discussion
3.1. The Basic Information of the Transcriptome
3.2. Identification and Verification of DEGs
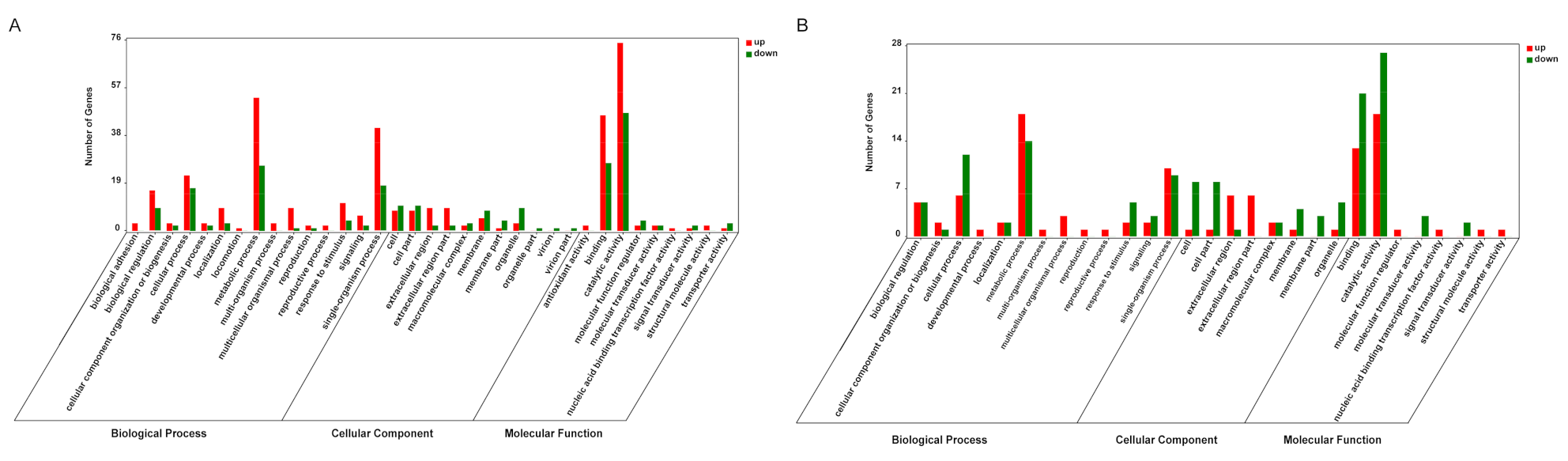
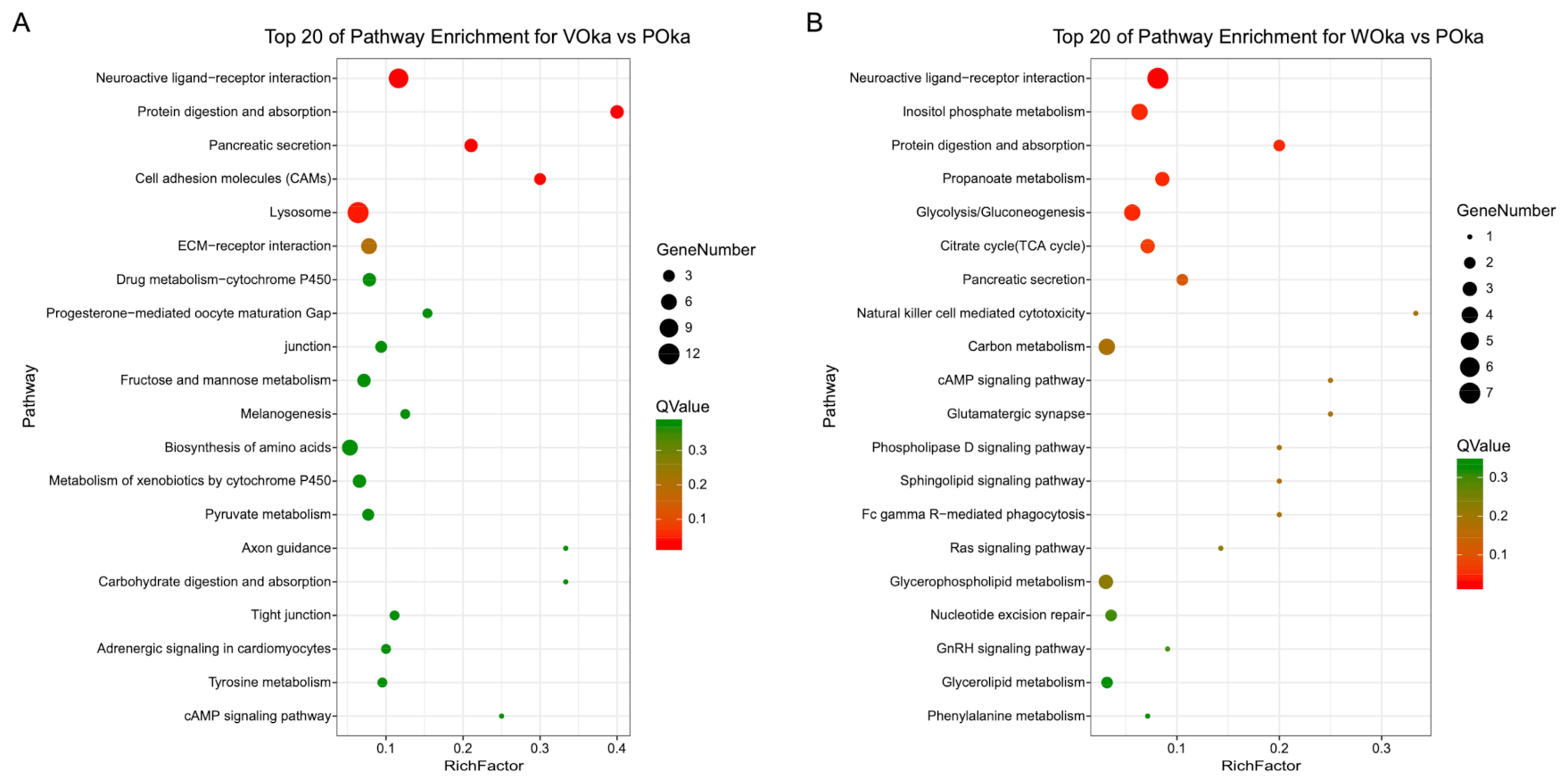
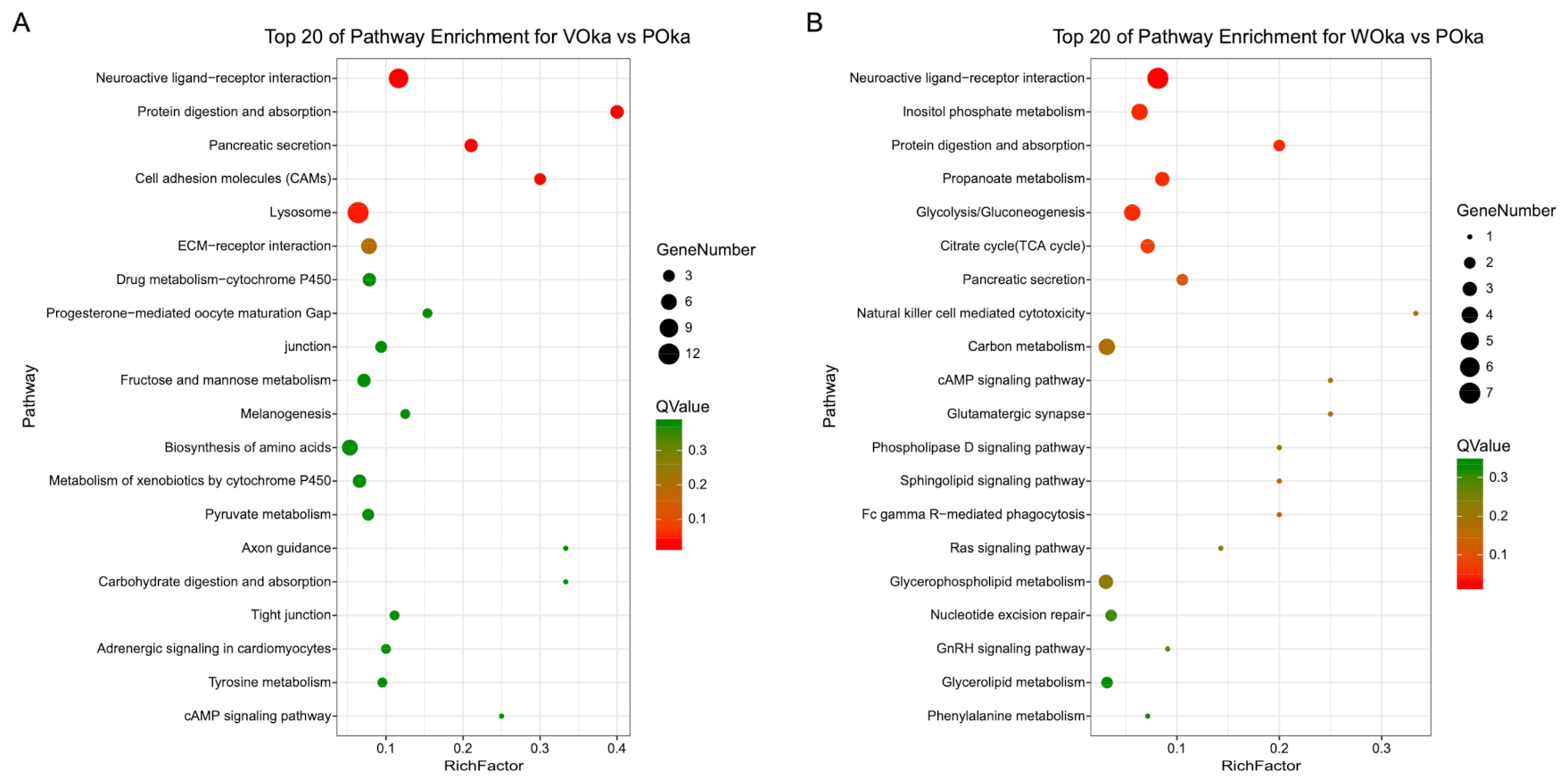
3.3. Functional Characterization of DEGs
3.4. In-Depth Analysis of Immune-Related DEGs
3.4.1. PRRs
3.4.2. ProPO Activating System
3.4.3. Signaling Pathways
3.4.4. Phagocytosis
3.4.5. Chitin-Binding Proteins (CBPs)
3.4.6. Immune Effectors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALFs | Anti-Lipopolysaccharide Factors |
| CBPs | Chitin-Binding Proteins |
| DEGs | Differentially Expressed Genes |
| HMC | Hemocyanin |
| PRRs | Pattern Recognition Receptors |
| RPKM | Reads per Kilobase per Million Reads |
| VMO-I | Vitelline Membrane Outer Layer Protein I |
References
- Ghaffari, N.; Sánchez-Flores, A.; Doan, R.; Garcia-Orozco, K.D.; Chen, P.L.; Ochoa-Leyva, A.; López-Zavala, A.A.; Carrasco, J.S.; Hong, C.; Brieba, L.G.; et al. Novel transcriptome assembly and improved annotation of the whiteleg shrimp (Litopenaeus vannamei), a dominant crustacean in global seafood mariculture. Sci. Rep. 2014, 4, 7081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thitamadee, S.; Prachumwat, A.; Srisala, J.; Jaroenlak, P.; Salachan, P.V.; Sritunyalucksana, K.; Flegel, T.W.; Itsathitphaisarn, O. Review of current disease threats for cultivated penaeid shrimp in Asia. Aquaculture 2016, 452, 69–87. [Google Scholar] [CrossRef]
- Citarasu, T.; Sivaram, V.; Immanuel, G.; Rout, N.; Murugan, V. Influence of selected Indian immunostimulant herbs against white spot syndrome virus (WSSV) infection in black tiger shrimp, Penaeus monodon with reference to haematological, biochemical and immunological changes. Fish Shellfish Immunol. 2006, 21, 372–384. [Google Scholar] [CrossRef]
- Tran, L.; Nunan, L.; Redman, R.M.; Mohney, L.L.; Pantoja, C.R.; Fitzsimmons, K.; Lightner, D.V. Determination of the infectious nature of the agent of acute hepatopancreatic necrosis syndrome affecting penaeid shrimp. Dis. Aquat. Org. 2013, 105, 45–55. [Google Scholar] [CrossRef]
- Li, F.; Xiang, J. Recent advances in researches on the innate immunity of shrimp in China. Dev. Comp. Immunol. 2013, 39, 11–26. [Google Scholar] [CrossRef]
- Söderhäll, I. Crustacean hematopoiesis. Dev. Comp. Immunol. 2016, 58, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zheng, S.-C.; Li, Y.-L.; Li, J.; Liu, H.-P. Hemocyte-Mediated Phagocytosis in Crustaceans. Front. Immunol. 2020, 11, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oka, M. Studies on Penaeus orientalis Kishinouye-VIII structure of the newly found lymphoid organ. Bull. Jpn. Soc. Sci. Fish. 1969, 35, 245–250. [Google Scholar] [CrossRef]
- Lightner, D.V.; Hedrick, R.P.; Fryer, J.L.; Chen, S.N.; Liao, I.C.; Kou, G.H. A survey of cultured penaeid shrimp in Taiwan for viral and other important diseases. Fish. Pathol. 1987, 22, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Bonami, J.R.; Lightner, D.V.; Redman, R.M.; Poulos, B.T. Partial Characterization of a Togavirus (Lovv) Associated with Histopathological Changes of the Lymphoid Organ of Penaeid Shrimps. Dis. Aquat. Organ. 1992, 14, 145–152. [Google Scholar] [CrossRef]
- Rusaini; Owens, L. Insight into the lymphoid organ of penaeid prawns: A review. Fish Shellfish Immunol. 2010, 29, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Wu, L.; Jin, M.; Li, T.; Hui, K.; Ren, Q. Transcriptome profiling of the Macrobrachium rosenbergii lymphoid organ under the white spot syndrome virus challenge. Fish Shellfish Immunol. 2017, 67, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.-Q. Comparative transcriptome analysis reveals three potential antiviral signaling pathways in lymph organ tissue of the red swamp crayfish, Procambarus clarkii. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Van de Braak, C.; Botterblom, M.; Taverne, N.; Van Muiswinkel, W.; Rombout, J.; Van der Knaap, W. The roles of haemocytes and the lymphoid organ in the clearance of injected Vibrio bacteria in Penaeus monodon shrimp. Fish Shellfish Immunol. 2002, 13, 293–309. [Google Scholar] [CrossRef]
- Burgents, J.E.; Burnett, L.E.; Stabb, E.V.; Burnett, K.G. Localization and bacteriostasis of Vibrio introduced into the Pacific white shrimp, Litopenaeus vannamei. Dev. Comp. Immunol. 2005, 29, 681–691. [Google Scholar] [CrossRef]
- Anggraeni, M.S.; Owens, L. The haemocytic origin of lymphoid organ spheroid cells in the penaeid prawn Penaeus monodon. Dis. Aquat. Org. 2000, 40, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Guo, S.; Li, F.; Xiang, J. Characterization and function analysis of an anti-lipopolysaccharide factor (ALF) from the Chinese shrimp Fenneropenaeus chinensis. Dev. Comp. Immunol. 2014, 46, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, F.; Chi, Y.; Xiang, J. Enhanced resistance of marine shrimp Exopalamon carincauda Holthuis to WSSV by injecting live VP28-recombinant bacteria. Acta Oceanol. Sin. 2013, 32, 52–58. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef] [Green Version]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B.; et al. TIGR Gene Indices clustering tools (TGICL): A software system for fast clustering of large EST datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conesa, A.; Götz, S.; Garcia-Gomez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, I.-T.; Aoki, T.; Huang, Y.-T.; Hirono, I.; Chen, T.-C.; Huang, J.-Y.; Chang, G.-D.; Lo, C.-F.; Wang, H.-C. White Spot Syndrome Virus Induces Metabolic Changes Resembling the Warburg Effect in Shrimp Hemocytes in the Early Stage of Infection. J. Virol. 2011, 85, 12919–12928. [Google Scholar] [CrossRef] [Green Version]
- Mathew, S.; Nair, A.K.K.; Anandan, R.; Gopalan, P.; Nair, N.V.; Devadasan, K. Biochemical studies on changes associated with enzymes of glucose metabolism in white spot syndrome virus (WSSV) infected with Penaeus monodon (Fabricius). Afr. J. Biotechnol. 2007, 6. [Google Scholar] [CrossRef]
- Su, M.-A.; Huang, Y.-T.; Chen, I.-T.; Lee, D.-Y.; Hsieh, Y.-C.; Li, C.-Y.; Ng, T.H.; Liang, S.-Y.; Lin, S.-Y.; Huang, S.-W. An invertebrate Warburg effect: A shrimp virus achieves successful replication by altering the host metabolome via the PI3K-Akt-mTOR pathway. PLOS Pathog. 2014, 10, e1004196. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Zhang, X. Host defense against DNA virus infection in shrimp is mediated by the siRNA pathway. Eur. J. Immunol. 2013, 43, 137–146. [Google Scholar] [CrossRef]
- Li, F.; Xiang, J. Signaling pathways regulating innate immune responses in shrimp. Fish Shellfish Immunol. 2013, 34, 973–980. [Google Scholar] [CrossRef]
- Labreuche, Y.; Warr, G.W. Insights into the antiviral functions of the RNAi machinery in penaeid shrimp. Fish Shellfish Immunol. 2013, 34, 1002–1010. [Google Scholar] [CrossRef] [Green Version]
- Walsh, M.C.; Lee, J.; Choi, Y. Tumor necrosis factor receptor- associated factor 6 (TRAF6) regulation of development, function, and homeostasis of the immune system. Immunol. Rev. 2015, 266, 72–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.H.; Wan, D.H.; Gu, Z.H.; Deng, X.X.; Weng, S.P.; Yu, X.Q.; He, J.G. Litopenaeus vannamei tumor necrosis factor receptor-associated factor 6 (TRAF6) responds to Vibrio alginolyticus and white spot syndrome virus (WSSV) infection and activates antimicrobial peptide genes. Dev. Comp. Immunol. 2011, 35, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Huang, Y.; Wang, B.; Jian, J.; Xu, Y. Tumor necrosis factor receptor-associated factor 6 (TRAF6) participates in peroxinectin gene expression in Fenneropenaeus penicillatus. Fish Shellfish Immunol. 2017, 64, 193–201. [Google Scholar] [CrossRef]
- Wang, S.; Song, X.; Zhang, Z.; Li, H.; Lǚ, K.; Yin, B.; He, J.; Li, C. Shrimp with knockdown of LvSOCS2, a negative feedback loop regulator of JAK/STAT pathway in Litopenaeus vannamei, exhibit enhanced resistance against WSSV. Dev. Comp. Immunol. 2016, 65, 289–298. [Google Scholar] [CrossRef]
- Sun, J.-J.; Lan, J.-F.; Xu, J.-D.; Niu, G.-J.; Wang, J.-X. Suppressor of cytokine signaling 2 (SOCS2) negatively regulates the expression of antimicrobial peptides by affecting the Stat transcriptional activity in shrimp Marsupenaeus japonicus. Fish Shellfish Immunol. 2016, 56, 473–482. [Google Scholar] [CrossRef]
- Shen, Z.; Jacobs-Lorena, M. Evolution of Chitin-Binding Proteins in Invertebrates. J. Mol. Evol. 1999, 48, 341–347. [Google Scholar] [CrossRef]
- Yang, F.; Li, S.; Li, F.; Xiang, J. A cuticle protein from the Pacific white shrimp Litopenaeus vannamei involved in WSSV infection. Dev. Comp. Immunol. 2018, 81, 303–311. [Google Scholar] [CrossRef]
- Wang, L.; Li, F.; Wang, B.; Xiang, J. A new shrimp peritrophin-like gene from Exopalaemon carinicauda involved in white spot syndrome virus (WSSV) infection. Fish Shellfish Immunol. 2013, 35, 840–846. [Google Scholar] [CrossRef]
- Yang, F.; Li, X.; Li, S.; Xiang, J.; Li, F. A novel cuticle protein involved in WSSV infection to the Pacific white shrimp Litopenaeus vannamei. Dev. Comp. Immunol. 2020, 102, 103491. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.D.; Barracco, M.A. Antimicrobial peptides in crustaceans. Invertebr. Surviv. 2010, 7, 262–284. [Google Scholar]
- Kido, S.; Morimoto, A.; Kim, F.; Doi, Y. Isolation of a novel protein from the outer layer of the vitelline membrane. Biochem. J. 1992, 286, 17–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngernsoungnern, P.; Ngernsoungnern, A.; Chaiseha, Y.; Sretarugsa, P. Role of vitelline envelope during fertilization in the black tiger shrimp, Penaeus monodon. Acta Histochem. 2012, 114, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Pascucci, T.; Perrino, J.; Mahowald, A.; Waring, G. Eggshell Assembly inDrosophila:Processing and Localization of Vitelline Membrane and Chorion Proteins. Dev. Biol. 1996, 177, 590–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, K.F.; Acorn, A.R.; Greenwood, S.J. A transcriptomic analysis of American lobster (Homarus americanus) immune response during infection with the bumper car parasite Anophryoides haemophila. Dev. Comp. Immunol. 2013, 40, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Back, J.F.; Bain, J.M.; Vadehra, D.; Burley, R. Proteins of the outer layer of the vitelline membrane of hen’s eggs. Biochim. et Biophys. Acta (BBA)—Protein Struct. Mol. Enzym. 1982, 705, 12–19. [Google Scholar] [CrossRef]
- Sun, Y.; Li, F.; Xiang, J. Analysis on the dynamic changes of the amount of WSSV in Chinese shrimp Fenneropenaeus chinensis during infection. Aquaculture 2013, 376-379, 124–132. [Google Scholar] [CrossRef]
- Wang, X.-W.; Wang, J.-X. Pattern recognition receptors acting in innate immune system of shrimp against pathogen infections. Fish Shellfish Immunol. 2013, 34, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Albores, F.; Yepiz-Plascencia, G. Beta glucan binding protein and its role in shrimp immune response. Aquaculture 2000, 191, 13–21. [Google Scholar] [CrossRef]
- Wang, X.-W.; Wang, J.-X. Diversity and multiple functions of lectins in shrimp immunity. Dev. Comp. Immunol. 2013, 39, 27–38. [Google Scholar] [CrossRef]
- Junkunlo, K.; Prachumwat, A.; Tangprasittipap, A.; Senapin, S.; Borwornpinyo, S.; Flegel, T.W.; Sritunyalucksana, K. A novel lectin domain-containing protein (LvCTLD) associated with response of the whiteleg shrimp Penaeus (Litopenaeus) vannamei to yellow head virus (YHV). Dev. Comp. Immunol. 2012, 37, 334–341. [Google Scholar] [CrossRef]
- Zhao, Z.-Y.; Yin, Z.-X.; Xu, X.-P.; Weng, S.-P.; Rao, X.-Y.; Dai, Z.-X.; Luo, Y.-W.; Yang, G.; Li, Z.-S.; Guan, H.-J.; et al. A Novel C-Type Lectin from the Shrimp Litopenaeus vannamei Possesses Anti-White Spot Syndrome Virus Activity. J. Virol. 2009, 83, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Lage, C.; Bok Luel, L.; Kenneth, S.D.L. The proPO-system: Pros and cons for its role in invertebrate immunity. Trends. Immunol. 2008, 29, 263–271. [Google Scholar]
- Amparyup, P.; Charoensapsri, W.; Tassanakajon, A. Prophenoloxidase system and its role in shrimp immune responses against major pathogens. Fish Shellfish Immunol. 2013, 34, 990–1001. [Google Scholar] [CrossRef] [PubMed]
- González-Santoyo, I.; Córdoba-Aguilar, A. Phenoloxidase: A key component of the insect immune system. Entomol. Exp. Appl. 2012, 142, 1–16. [Google Scholar] [CrossRef]
- Le Clec’h, W.; Anderson, T.J.C.; Chevalier, F.D. Characterization of hemolymph phenoloxidase activity in two Biomphalaria snail species and impact of Schistosoma mansoni infection. Parasites Vectors 2016, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dittmer, N.T.; Kanost, M.R. Insect multicopper oxidases: Diversity, properties, and physiological roles. Insect Biochem. Mol. Biol. 2010, 40, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Chan, S.; Li, C.; Zhang, S. Identification and characterization of a laccase from Litopenaeus vannamei involved in anti-bacterial host defense. Fish Shellfish Immunol. 2017, 66, 1–10. [Google Scholar] [CrossRef]
- Coates, C.J.; Nairn, J. Diverse immune functions of hemocyanins. Dev. Comp. Immunol. 2014, 45, 43–55. [Google Scholar] [CrossRef]
- Flannagan, R.S.; Jaumouillé, V.; Grinstein, S. The cell biology of phagocytosis. Annu. Rev. Pathol. 2012, 7, 61–98. [Google Scholar] [CrossRef]
- Li, W.-W.; Jin, X.-K.; He, L.; Jiang, H.; Gong, Y.-N.; Xie, Y.-N.; Wang, Q. Molecular cloning, characterization, expression and activity analysis of cathepsin L in Chinese mitten crab, Eriocheir sinensis. Fish Shellfish Immunol. 2010, 29, 1010–1018. [Google Scholar] [CrossRef]
- Mohamed, M.M.; Sloane, B.F. Cysteine cathepsins: Multifunctional enzymes in cancer. Nat. Rev. Cancer. 2006, 6, 764–775. [Google Scholar] [CrossRef]
- Liaudet-Coopman, E.; Beaujouin, M.; Derocq, D.; Garcia, M.; Glondu-Lassis, M.; Laurent-Matha, V.; Prébois, C.; Rochefort, H.; Vignon, F. Cathepsin D: Newly discovered functions of a long-standing aspartic protease in cancer and apoptosis. Cancer Lett. 2006, 237, 167–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Q.; Zhang, X.-W.; Sun, Y.-D.; Sun, S.-S.; Zhou, J.; Wang, Z.-H.; Zhao, X.-F.; Wang, J.-X. Two cysteine proteinases respond to bacterial and WSSV challenge in Chinese white shrimp Fenneropenaeus chinensis. Fish Shellfish Immunol. 2010, 29, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Chen, L.; Zhang, F.; Wu, P.; Li, E.; Qin, J. Molecular characterization of cathepsin L cDNA and its expression during oogenesis and embryogenesis in the oriental river prawn Macrobrachium nipponense (Palaemonidae). Genet. Mol. Res 2013, 12, 5215–5225. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Jiang, S.; Huang, J.; Wang, W.; Zhang, D.; Wu, Q.; Yang, K. Molecular cloning and mRNA expression of cathepsin C gene in black tiger shrimp (Penaeus monodon). Comp. Biochem. Physiol. Part. A Mol. Integr. Physiol. 2008, 150, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.S.; Lin, S.T.; Wu, Z.H.; Jian, J.C. Molecular cloning and mRNA expression of cathepsin C in white shrimp, Litopenaeus vannamei. Aquac. Res. 2011, 42, 1569–1576. [Google Scholar]
- Yu, X.-M.; Chen, J.-L.; Abbas, M.N.; Gul, I.; Kausar, S.; Dai, L.-S. Characterization of the cathepsin D in Procambarus clarkii and its biological role in innate immune responses. Dev. Comp. Immunol. 2020, 111, 103766. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Raw Reads | Adapter (%) | Low Quality (%) | Clean Reads (%) | GC | Clean Bases (bp) | Q20 (%) |
|---|---|---|---|---|---|---|---|
| POka-1 | 50,743,734 | 39,416 (0.08%) | 1,532,076 (3.02%) | 49,170,842 (96.90%) | 45.41% | 7,351,935,942 | 7,049,298,471 (95.88%) |
| POka-2 | 45,500,944 | 32,062 (0.07%) | 1,515,748 (3.33%) | 43,952,044 (96.60%) | 45.24% | 6,571,862,271 | 6,288,283,729 (95.68%) |
| POka-3 | 56,141,350 | 44,672 (0.08%) | 1,919,290 (3.42%) | 54,175,832 (96.50%) | 46.58% | 8,097,101,318 | 7,747,188,736 (95.68%) |
| VOka-1 | 45,722,036 | 64,476 (0.14%) | 1,640,504 (3.59%) | 44,016,176 (96.27%) | 48.29% | 6,573,342,054 | 6,284,318,401 (95.60%) |
| VOka-2 | 61,168,912 | 81,778 (0.13%) | 2,388,808 (3.91%) | 58,697,110 (95.96%) | 49.48% | 8,769,829,446 | 8,372,020,539 (95.46%) |
| VOka-3 | 50,048,002 | 77,522 (0.15%) | 1,890,092 (3.78%) | 48,079,500 (96.07%) | 49.02% | 7,180,638,897 | 6,859,211,988 (95.52%) |
| WOka-1 | 59,908,592 | 95,602 (0.16%) | 2,260,780 (3.77%) | 57,551,410 (96.07%) | 50.18% | 8,599,297,116 | 8,212,992,611 (95.51%) |
| WOka-2 | 41,272,526 | 55,908 (0.14%) | 1,723,184 (4.18%) | 39,492,746 (95.69%) | 48.27% | 5,896,503,470 | 5,621,053,593 (95.33%) |
| WOka-3 | 41,909,626 | 76,846 (0.18%) | 1,458,246 (3.48%) | 40,373,928 (96.34%) | 50.29% | 6,028,800,808 | 5,764,943,773 (95.62%) |
| Classification | Gene ID | Nr-Annotation | log2 Ratio (VOka/POka) | log2 Ratio (WOka/POka) |
|---|---|---|---|---|
| BGBP | Unigene0006001 | beta-1,3-glucan-binding protein precursor | 9.2689 | not significant |
| Unigene0006002 | beta-1,3-glucan-binding protein precursor | 7.2856 | not significant | |
| Unigene0011623 | beta-1,3-glucan-binding protein precursor | 5.8020 | not significant | |
| Unigene0006003 | beta-1,3-glucan binding protein | 7.1715 | not significant | |
| Unigene0022216 | beta-1,3-glucan binding protein | 13.0141 | not significant | |
| Unigene0020907 | Beta-1,3-glucan-binding protein | 9.9063 | not significant | |
| LGBP | Unigene0011641 | lipopolysaccharide and beta-1,3-glucan binding protein | 5.6731 | not significant |
| CTL | Unigene0021953 | antiviral protein | 7.1551 | not significant |
| Unigene0007029 | mannose-binding protein | 4.1113 | not significant | |
| Unigene0011922 | lectin D, partial | 7.0167 | not significant | |
| Unigene0001862 | lectin B isoform 3, partial | 7.1265 | not significant | |
| Unigene0009722 | lectin B isoform 3, partial | 7.4837 | not significant | |
| Unigene0007283 | C-type lectin 5 | 7.8437 | not significant | |
| Unigene0025672 | C-type lectin 4 | 6.1137 | not significant | |
| Unigene0026420 | C-type lectin 2 | 4.8577 | not significant | |
| Unigene0005566 | C-type lectin 1 | 8.0877 | not significant | |
| Unigene0036730 | C-type lectin 1 | 6.4531 | not significant | |
| Unigene0056556 | C-type lectin 1 | 8.6933 | not significant | |
| Unigene0008649 | C-type lectin | 3.8902 | not significant | |
| Unigene0042720 | C-type lectin | 6.5687 | not significant | |
| Unigene0004121 | C-type lectin | 11.8192 | not significant | |
| Unigene0040089 | C-type lectin | 5.4285 | not significant | |
| Unigene0042727 | C-type lectin | 7.9989 | not significant | |
| Unigene0028929 | C-type lectin | 3.8404 | 2.7238 | |
| Unigene0029885 | C-type lectin | 8.7081 | 4.7517 | |
| others | Unigene0045581 | Protein NLRC5 | 2.6992 | not significant |
| Unigene0037611 | protein NLRC5-like | 3.1469 | not significant | |
| Unigene0012727 | fibrinogen-like protein | 4.5223 | not significant | |
| Unigene0022681 | FREP2 | not significant | −3.8725 | |
| Unigene0038302 | leucine rich repeat only protein | 7.9370 | not significant |
| Classification | Gene ID | Nr-Annotation | log2 Ratio (VOka/POka) | log2 Ratio (WOka/POka) |
|---|---|---|---|---|
| SP | Unigene0006596 | preprochymotrypsin 1 | 7.5397 | not significant |
| Unigene0008624 | Chymotrypsin BII | 6.7811 | not significant | |
| Unigene0040871 | Chymotrypsin BII | 6.7003 | not significant | |
| Unigene0032512 | chymotrypsin-2-like | 6.4923 | not significant | |
| Unigene0032601 | trypsin | 6.1049 | not significant | |
| Unigene0006379 | trypsin | 6.3933 | not significant | |
| Unigene0010203 | trypsin-1 | −5.3595 | not significant | |
| Unigene0007993 | trypsin-1 | −4.9586 | not significant | |
| Unigene0031678 | venom protease-like | 4.0204 | not significant | |
| Unigene0007586 | CUB-serine protease | 4.0236 | not significant | |
| Unigene0024520 | clip domain serine proteinase 2 | 2.8457 | not significant | |
| Unigene0033178 | serine protease | 3.8166 | not significant | |
| Unigene0009569 | transmembrane protease serine 12-like | 4.9554 | not significant | |
| Unigene0013356 | coagulation factor IX-like | −3.2687 | not significant | |
| Unigene0008625 | preprochymotrypsin 1 | 7.3505 | 4.2663 | |
| Unigene0040873 | preprochymotrypsin 1 | 6.9669 | 3.8781 | |
| Unigene0023371 | Trypsin-7 | −5.6098 | −5.9965 | |
| Unigene0006380 | trypsin, partial | 6.6686 | 4.7227 | |
| Unigene0042231 | trypsin, partial | 6.4265 | 4.6668 | |
| Unigene0006516 | trypsin | 8.8686 | 8.7520 | |
| Unigene0007302 | serine protease SP24D | not significant | −14.7897 | |
| Unigene0028519 | clip domain serine proteinase 1 | not significant | −3.5529 | |
| SPI | Unigene0004226 | inter-alpha-trypsin inhibitor heavy chain H3 | 7.8262 | not significant |
| Unigene0037147 | Kazal-type serine proteinase inhibitor 4 | 7.4926 | not significant | |
| Unigene0030648 | Kazal-type protease inhibitor | 5.3919 | not significant | |
| Unigene0012105 | serine proteinase inhibitor-3 | −6.3212 | not significant | |
| Unigene0040667 | serine proteinase inhibitor 8 | 3.5000 | not significant | |
| Unigene0040665 | serine proteinase inhibitor | −3.7258 | not significant | |
| Unigene0031232 | Kunitz-type serine protease inhibitor | 4.1047 | 3.1427 | |
| Unigene0033607 | kazal type protease inhibitor | 3.1518 | 2.6125 | |
| Unigene0040669 | serine proteinase inhibitor 8 | 3.8173 | 3.3256 | |
| Unigene0006314 | serine proteinase inhibitor | 3.8517 | 3.4715 | |
| Unigene0028753 | serine proteinase inhibitor | 4.8338 | 5.5177 | |
| Unigene0040666 | serine proteinase inhibitor | 4.1175 | 3.7476 | |
| proPO | Unigene0006825 | laccase-1-like isoform X1 | 5.3797 | not significant |
| Unigene0040447 | Laccase 2 | 8.2782 | not significant | |
| Unigene0043665 | hemocyanin, partial | 8.9674 | not significant | |
| Unigene0006113 | hemocyanin subunit L4, partial | 7.4781 | not significant | |
| Unigene0043668 | hemocyanin subunit L4, partial | 6.2013 | not significant | |
| Unigene0008173 | hemocyanin subunit L3, partial | 14.5992 | not significant | |
| Unigene0006336 | hemocyanin subunit L2, partial | 8.0490 | not significant | |
| Unigene0043663 | hemocyanin subunit L2, partial | 10.1345 | not significant | |
| Unigene0006337 | hemocyanin subunit L1, partial | 6.3323 | not significant | |
| Unigene0043667 | hemocyanin subunit L1, partial | 8.4202 | not significant | |
| Unigene0025329 | hemocyanin | 6.0904 | not significant | |
| Unigene0043666 | hemocyanin | 12.5672 | not significant |
| Gene ID | Nr-Annotation | log2 Ratio (VOka/POka) | log2 Ratio (WOka/POka) |
|---|---|---|---|
| Unigene0014407 | invertebrate-type lysozyme protein | −5.7228 | not significant |
| Unigene0015966 | invertebrate-type lysozyme 2 | −6.8231 | not significant |
| Unigene0043096 | beta-hexosaminidase subunit alpha-like | 5.1646 | not significant |
| Unigene0039600 | cathepsin L, partial | 7.9327 | not significant |
| Unigene0004352 | cathepsin L | 8.8075 | not significant |
| Unigene0004353 | cathepsin L | 7.1853 | not significant |
| Unigene0039602 | cathepsin L, partial | 8.0292 | not significant |
| Unigene0044696 | cathepsin C | 7.4405 | not significant |
| Unigene0044700 | cathepsin C | 6.2937 | not significant |
| Unigene0038874 | cathepsin D | 5.8533 | not significant |
| Unigene0043169 | tick legumain | 7.3633 | not significant |
| Unigene0043171 | tick legumain | 8.0169 | not significant |
| Unigene0019904 | destabilase I | 7.2670 | not significant |
| Unigene0008646 | gamma-interferon induced thiol reductase GILT3 | 5.1848 | not significant |
| Classification | Gene ID | Nr-Annotation | log2 Ratio (VOka/POka) | log2 Ratio (WOka/POka) |
|---|---|---|---|---|
| ALF | Unigene0031278 | anti-lipopolysaccharide factor isoform 6 | 2.5922 | not significant |
| Unigene0028455 | anti-lipopolysaccharide factor isoform 5 | 2.5390 | not significant | |
| Crustin | Unigene0013411 | crustin 3 | 9.6385 | 9.3923 |
| Unigene0053122 | crustin Pm5 | −10.2677 | not significant | |
| Unigene0023765 | CruI-1 | not significant | −9.2311 | |
| VMOI | Unigene0004307 | vitelline membrane outer layer protein I like protein | 9.2916 | not significant |
| Unigene0035626 | Vitelline membrane outer layer protein 1 | 7.9527 | not significant | |
| Unigene0004073 | vitelline membrane outer layer 1-like protein | 7.0706 | not significant | |
| Unigene0009081 | vitelline membrane outer layer 1-like protein | 7.8432 | not significant | |
| Unigene0021817 | vitelline membrane outer layer 1-like protein | 7.9325 | not significant | |
| Unigene0021818 | vitelline membrane outer layer 1-like protein | 8.1444 | not significant | |
| Unigene0037137 | vitelline membrane outer layer 1-like protein | 8.0456 | not significant | |
| Unigene0057916 | vitelline membrane outer layer 1-like protein | 7.3888 | not significant |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Li, S.; Li, F. Different Immune Responses of the Lymphoid Organ in Shrimp at Early Challenge Stage of Vibrio parahaemolyticus and WSSV. Animals 2021, 11, 2160. https://doi.org/10.3390/ani11082160
Wang F, Li S, Li F. Different Immune Responses of the Lymphoid Organ in Shrimp at Early Challenge Stage of Vibrio parahaemolyticus and WSSV. Animals. 2021; 11(8):2160. https://doi.org/10.3390/ani11082160
Chicago/Turabian StyleWang, Fuxuan, Shihao Li, and Fuhua Li. 2021. "Different Immune Responses of the Lymphoid Organ in Shrimp at Early Challenge Stage of Vibrio parahaemolyticus and WSSV" Animals 11, no. 8: 2160. https://doi.org/10.3390/ani11082160
APA StyleWang, F., Li, S., & Li, F. (2021). Different Immune Responses of the Lymphoid Organ in Shrimp at Early Challenge Stage of Vibrio parahaemolyticus and WSSV. Animals, 11(8), 2160. https://doi.org/10.3390/ani11082160