
Is There a “Right” Side of Communicating Friendship? Lateralization of Social Interactions in Zoo Barbary macaques (Macaca sylvanus)


 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Housing
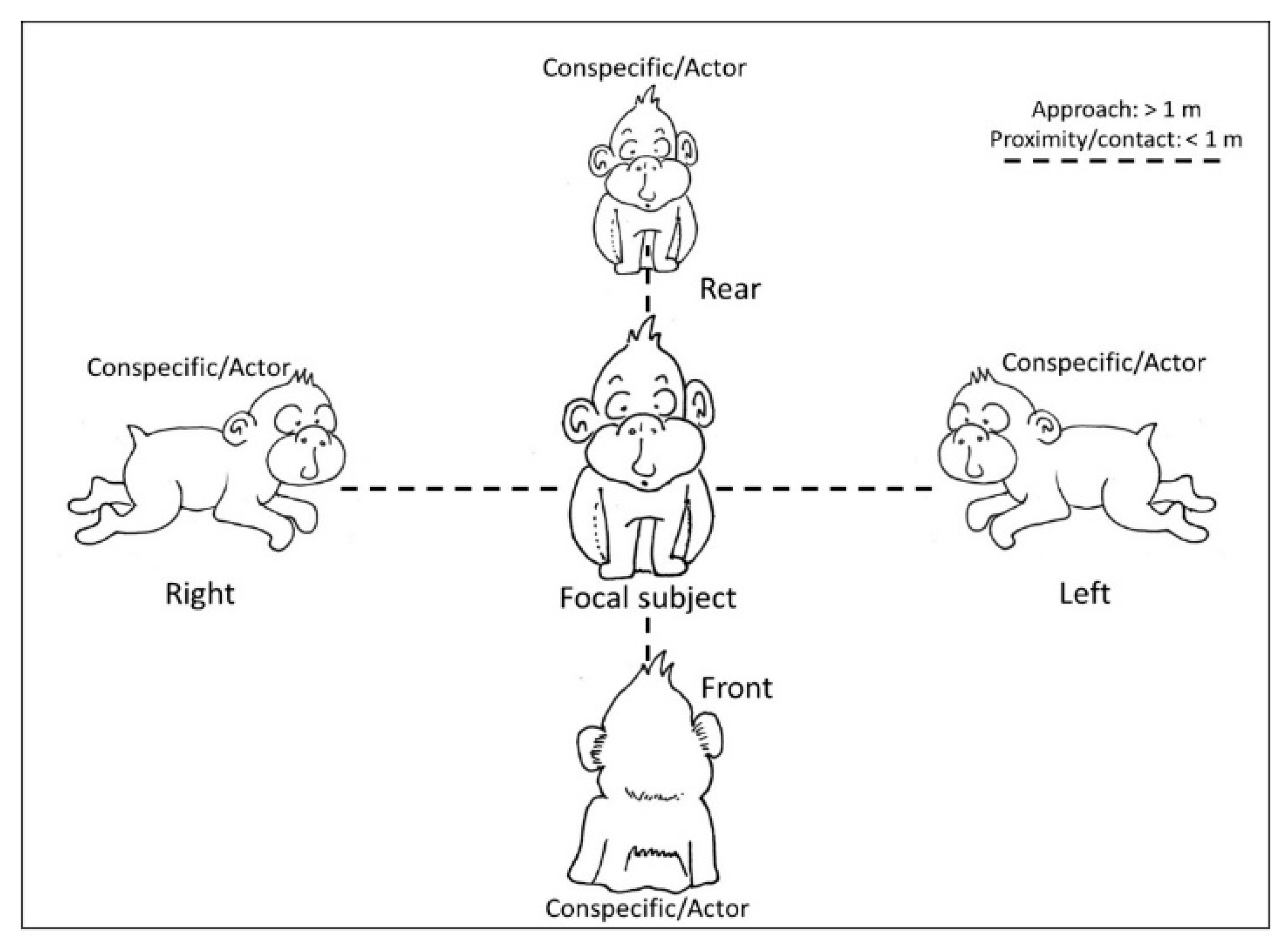
2.2. Data Collection
2.3. Data Analysis
3. Results
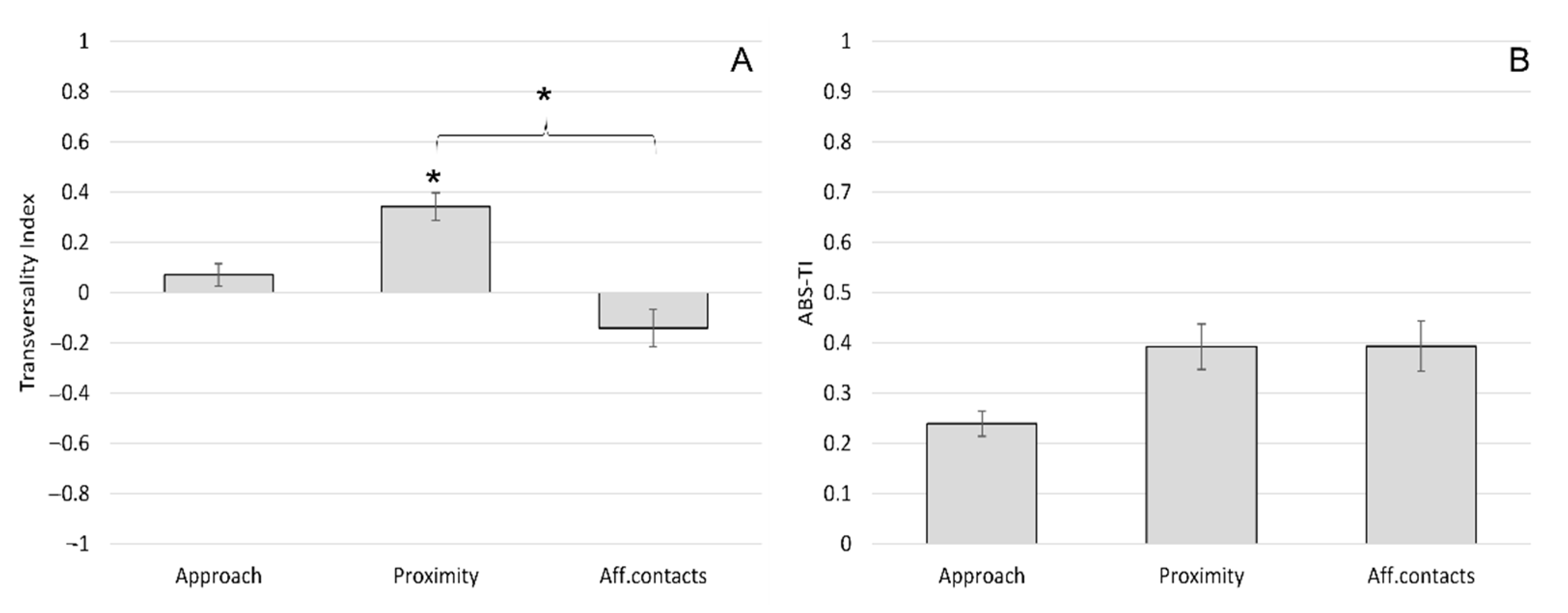
3.1. Side Preferences during Social Interactions
3.2. Sagittal Preferences during Social Interactions
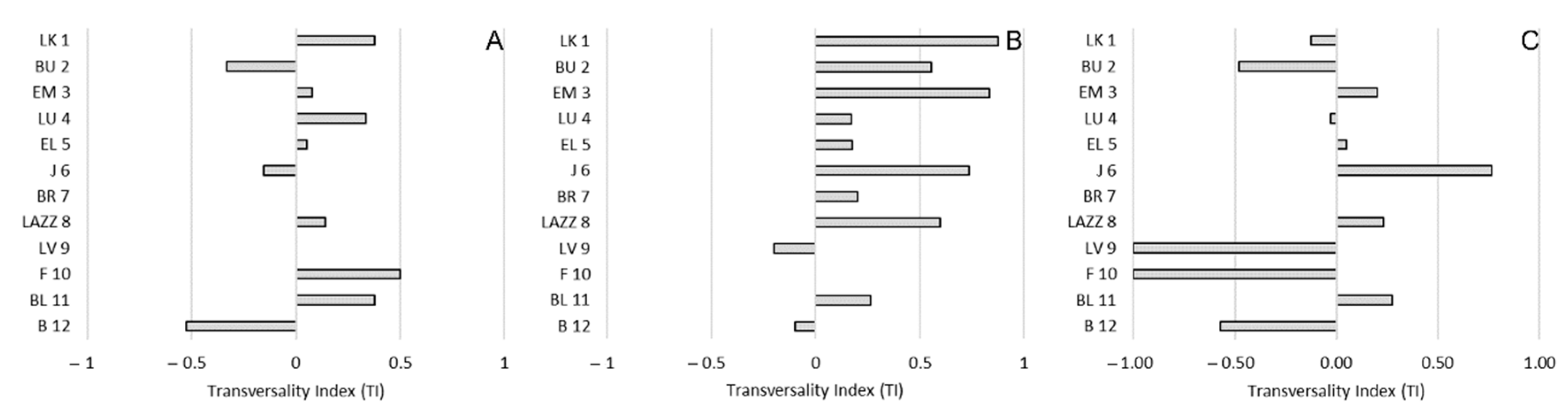
3.3. Side/sagittal Preference and Social Rank
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacNeilage, P.F.; Rogers, L.J.; Vallortigara, G. Origins of the left & right brain. Sci. Am. 2009, 301, 60–67. [Google Scholar] [CrossRef]
- Ghirlanda, S.; Vallortigara, G. The evolution of brain lateralization: A game-theoretical analysis of population structure. Proc. R. Soc. B Boil. Sci. 2004, 271, 853–857. [Google Scholar] [CrossRef] [Green Version]
- Rogers, L.J.; Vallortigara, G.; Andrew, R.J. Divided Brains: The Biology and Behaviour of Brain Asymmetries; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Rogers, L.J.; Zucca, P.; Vallortigara, G. Advantages of having a lateralized brain. Proc. R. Soc. B Boil. Sci. 2004, 271, S420–S422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salva, O.R.; Regolin, L.; Vallortigara, G. Inversion of contrast polarity abolishes spontaneous preferences for face-like stimuli in newborn chicks. Behav. Brain Res. 2012, 228, 133–143. [Google Scholar] [CrossRef]
- Robins, A.; Lippolis, G.; Bisazza, A.; Vallortigara, G.; Rogers, L.J. Lateralized agonistic responses and hindlimb use in toads. Anim. Behav. 1998, 56, 875–881. [Google Scholar] [CrossRef] [Green Version]
- Pellitteri-Rosa, D.; Gazzola, A. Context-dependent behavioural lateralization in the European pond turtle Emys orbicularis (Testudines, Emydidae). J. Exp. Biol. 2018, 221, jeb.186775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deckel, W.A. Laterality of aggressive responses in Anolis. J. Exp. Zool. 1995, 272, 194–200. [Google Scholar] [CrossRef]
- Hews, D.K.; Worthington, R.A. Fighting from the right side of the brain: Left visual field preference during aggression in free-ranging male tree lizards (Urosaurus ornatus). Brain Behav. Evol. 2001, 58, 356–361. [Google Scholar] [CrossRef]
- Nagy, M.; Ákos, Z.; Biro, D.; Vicsek, T. Hierarchical group dynamics in pigeon flocks. Nature 2010, 464, 890–893. [Google Scholar] [CrossRef]
- Karenina, K.; Giljov, A.; Ingram, J.; Rowntree, V.J.; Malashichev, Y. Lateralization of mother–infant interactions in a diverse range of mammal species. Nat. Ecol. Evol. 2017, 1, 30. [Google Scholar] [CrossRef]
- Giljov, A.; Karenina, K.; Malashichev, Y. Facing each other: Mammal mothers and infants prefer the position favouring right hemisphere processing. Biol. Lett. 2018, 14, 20170707. [Google Scholar] [CrossRef] [PubMed]
- Hook, M.; Rogers, L. Lateralized use of the mouth in production of vocalizations by marmosets. Neuropsychologia 1998, 36, 1265–1273. [Google Scholar] [CrossRef]
- De Latude, M.; Demange, M.; Bec, P.; Blois-Heulin, C. Visual laterality responses to different emotive stimuli by red-capped mangabeys, Cercocebus torquatus torquatus. Anim. Cogn. 2008, 12, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Wallez, C.; Vauclair, J. Right hemisphere dominance for emotion processing in baboons. Brain Cogn. 2011, 75, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Carriba, S.; Loeches, A.; Morcillo, A.; Hopkins, W.D. Functional asymmetry of emotions in primates: New findings in chimpanzees. Brain Res. Bull. 2002, 57, 561–564. [Google Scholar] [CrossRef]
- Roberts, A.I.; Murray, L.; Roberts, S.G.B. Complex sociality of wild chimpanzees can emerge from laterality of manual gestures. Hum. Nat. 2019, 30, 299–325. [Google Scholar] [CrossRef] [Green Version]
- Karenina, K.; Giljov, A. Mother and offspring lateralized social behavior across mammalian species. Prog. Brain Res. 2018, 238, 115–141. [Google Scholar] [CrossRef]
- Regaiolli, B.; Spiezio, C.; Hopkins, W.D. Asymmetries in mother-infant behaviour in Barbary macaques (Macaca sylvanus). PeerJ 2018, 6, e4736. [Google Scholar] [CrossRef] [Green Version]
- Casperd, J.M.; Dunbar, R. Asymmetries in the visual processing of emotional cues during agonistic interactions by gelada baboons. Behav. Process. 1996, 37, 57–65. [Google Scholar] [CrossRef]
- Baraud, I.; Buytet, B.; Bec, P.; Blois-Heulin, C. Social laterality and ‘transversality’ in two species of mangabeys: Influence of rank and implication for hemispheric specialization. Behav. Brain Res. 2009, 198, 449–458. [Google Scholar] [CrossRef]
- Quaresmini, C.; Forrester, G.S.; Spiezio, C.; Vallortigara, G. Social environment elicits lateralized behaviors in gorillas (Gorilla gorilla gorilla) and chimpanzees (Pan troglodytes). J. Comp. Psychol. 2014, 128, 276–284. [Google Scholar] [CrossRef] [Green Version]
- Picaud, S.; Dalkara, D.; Marazova, K.; Goureau, O.; Roska, B.; Sahel, J.-A. The primate model for understanding and restoring vision. Proc. Natl. Acad. Sci. USA 2019, 116, 26280–26287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stidwill, D.; Optom, F.C. Normal Binocular Vision: Theory, Investigation and Practical Aspects; Fletcher, R., Optom, F.C., Orth, D., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2010. [Google Scholar]
- Altmann, J. Observational study of behaviour: Sampling methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thierry, B.; Bynum, E.L.; Baker, S.; Kinnaird, M.F.; Matsumura, S.; Muroyama, Y.; O’Brien, T.G.; Petit, O.; Watanabe, K. The social repertoire of Sulawesi macaques. Primate Res. 2000, 16, 203–226. [Google Scholar] [CrossRef] [Green Version]
- Deag, J.M. A study of the Social Behaviour and Ecology of the Wild Barbary Macaque, Macaca sylvanus; University of Bristol: Bristol, UK, 1974. [Google Scholar]
- Sandri, C.; Regaiolli, B.; Vespiniani, A.; Spiezio, C. New food provision strategy for a colony of Barbary macaques (Macaca sylvanus): Effects on social hierarchy? Integr. Food Nutr. Metab. 2017, 4, 1–8. [Google Scholar] [CrossRef]
- Norscia, I.; Palagi, E. The socio-matrix reloaded: From hierarchy to dominance profile in wild lemurs. PeerJ 2015, 3, e729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang, A.; Deshpande, S.; Sumana, A.; Gadagkar, R. Choosing an appropriate index to construct dominance hierarchies in animal societies: A comparison of three indices. Anim. Behav. 2010, 79, 631–636. [Google Scholar] [CrossRef]
- Clutton-Brock, T.; Albon, S.; Gibson, R.; Guinness, F. The logical stag: Adaptive aspects of fighting in red deer (Cervus elaphus L.). Anim. Behav. 1979, 27, 211–225. [Google Scholar] [CrossRef]
- Michel, G.F.; Sheu, C.-F.; Brumley, M.R. Evidence of a right-shift factor affecting infant hand-use preferences from 7 to 11 months of age as revealed by latent class analysis. Dev. Psychobiol. 2002, 40, 1–13. [Google Scholar] [CrossRef]
- McGrew, W.C.; Marchant, L.F. Using the tools at hand: Manual laterality and elementary technology in Cebus spp. and Pan spp. Int. J. Primatol. 1997, 18, 787–810. [Google Scholar] [CrossRef]
- Boeving, E.R.; Belnap, S.C.; Nelson, E.L. Embraces are lateralized in spider monkeys (Ateles fusciceps rufiventris). Am. J. Primatol. 2017, 79, e22654. [Google Scholar] [CrossRef]
- Demaree, H.; Everhart, D.; Youngstrom, E.; Harrison, D.W. Brain Lateralization of emotional processing: Historical roots and a future incorporating “dominance”. Behav. Cogn. Neurosci. Rev. 2005, 4, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Kuhl, J.; Kazén, M. Motivation, affect, and hemispheric asymmetry: Power versus affiliation. J. Pers. Soc. Psychol. 2008, 95, 456–469. [Google Scholar] [CrossRef] [Green Version]
- Hecht, D. Cerebral lateralization of pro- and anti-social tendencies. Exp. Neurobiol. 2014, 23, 1–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, L.J.; Vallortigara, G. When and why did brains break symmetry? Symmetry 2015, 7, 2181–2194. [Google Scholar] [CrossRef] [Green Version]
- Boccia, M. Comparison of the physical characteristics of grooming in two species of macaques (Macaca nemestrina and M. radiata). J. Comp. Psychol. 1989, 103, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Allanic, M.; Hayashi, M.; Furuichi, T.; Matsuzawa, T. Social influences on grooming site preferences in wild bonobos (Pan paniscus) at Wamba, DRC. Primates 2020, 61, 213–223. [Google Scholar] [CrossRef]
- Thierry, M.; Kaumanns, B.; Singh, W. Macaque Societies: A Model for the Study of Social Organization; Cambridge University Press: Cambridge, UK, 2004. [Google Scholar]
- Thierry, B. Unity in diversity: Lessons from macaque societies. Evol. Anthr. Issues News Rev. 2007, 16, 224–238. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Approach | Proximity | Affiliative Contacts | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Subject | Sex | Age | NSI | SI | z-Score | NTI | TI | z-Score | NSI | SI | z-Score | NTI | TI | z-Score | NSI | SI | z-Score | NTI | TI | z-Score |
| Belinda (B) | F | 12 | 44 | 0.05 | 0.15 | 21 | −0.52 | −2.18 * | 49 | −0.06 | −0.29 | 20 | −0.10 | −0.22 | 21 | 0.24 | 0.87 | 14 | −0.57 | −1.87 |
| Belle (BL) | F | 4 | 66 | −0.24 | −1.85 | 29 | 0.38 | 1.86 | 82 | −0.22 | −1.88 | 19 | 0.26 | 0.92 | 42 | −0.48 | −2.93 * | 33 | 0.27 | 1.39 |
| Berta (BR) | F | 6 | 47 | 0.02 | 0.00 | 6 | 0.00 | # | 53 | 0.06 | 0.27 | 5 | 0.20 | # | 31 | −0.23 | −1.08 | 2 | 0.00 | # |
| Buddha (BU) | F | 13 | 17 | −0.18 | −0.49 | 18 | −0.33 | −1.18 | 16 | −0.25 | −0.75 | 9 | 0.56 | # | 26 | −0.23 | −0.98 | 27 | −0.48 | −2.31 * |
| Elly (EL) | F | 5 | 27 | −0.11 | −0.38 | 19 | 0.05 | 0.00 | 38 | −0.11 | −0.49 | 17 | 0.18 | 0.49 | 36 | −0.39 | −2.17 * | 21 | 0.05 | 0 |
| Emma (EM) | F | 5 | 22 | 0.09 | 0.21 | 13 | 0.08 | 0.00 | 20 | 0.00 | 0.00 | 12 | 0.83 | 2.6 * | 32 | 0.13 | 0.53 | 30 | 0.20 | 0.91 |
| Fanny (F) | F | 8 | 54 | 0.00 | 0.00 | 12 | 0.50 | 1.44 | 54 | −0.11 | −0.68 | 14 | 0.00 | 0.00 | 15 | −0.47 | −1.55 | 2 | −1.00 | # |
| Jack (J) | M | 4 | 37 | 0.30 | 1.64 | 26 | −0.15 | −0.59 | 31 | 0.23 | 1.08 | 23 | 0.74 | 3.34 * | 9 | 0.78 | # | 17 | 0.76 | 2.91 * |
| Labbraviola (LV) | F | 10 | 35 | 0.09 | 0.34 | 20 | 0.00 | 0.00 | 49 | 0.06 | 0.29 | 15 | −0.20 | −0.52 | 10 | 0.00 | 0.00 | 3 | −1.00 | # |
| Luke (LAZZ) | M | 4 | 24 | −0.25 | −1.02 | 14 | 0.14 | 0.27 | 29 | −0.17 | −0.74 | 10 | 0.60 | 1.58 | 5 | 0.20 | # | 13 | 0.23 | 0.55 |
| Lucky (LK) | M | 14 | 42 | 0.05 | 0.15 | 29 | 0.38 | 1.86 | 47 | −0.06 | −0.29 | 16 | 0.88 | 3.25 * | 25 | 0.04 | 0.00 | 32 | −0.13 | −0.53 |
| Lucrezia (LU) | F | 9 | 26 | −0.38 | −1.77 | 21 | 0.33 | 1.31 | 30 | −0.07 | −0.18 | 41 | 0.17 | 0.94 | 22 | −0.45 | −1.92 | 35 | −0.03 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldachini, M.; Regaiolli, B.; Llorente, M.; Riba, D.; Spiezio, C. Is There a “Right” Side of Communicating Friendship? Lateralization of Social Interactions in Zoo Barbary macaques (Macaca sylvanus). Animals 2021, 11, 3288. https://doi.org/10.3390/ani11113288
Baldachini M, Regaiolli B, Llorente M, Riba D, Spiezio C. Is There a “Right” Side of Communicating Friendship? Lateralization of Social Interactions in Zoo Barbary macaques (Macaca sylvanus). Animals. 2021; 11(11):3288. https://doi.org/10.3390/ani11113288
Chicago/Turabian StyleBaldachini, Marzia, Barbara Regaiolli, Miquel Llorente, David Riba, and Caterina Spiezio. 2021. "Is There a “Right” Side of Communicating Friendship? Lateralization of Social Interactions in Zoo Barbary macaques (Macaca sylvanus)" Animals 11, no. 11: 3288. https://doi.org/10.3390/ani11113288
APA StyleBaldachini, M., Regaiolli, B., Llorente, M., Riba, D., & Spiezio, C. (2021). Is There a “Right” Side of Communicating Friendship? Lateralization of Social Interactions in Zoo Barbary macaques (Macaca sylvanus). Animals, 11(11), 3288. https://doi.org/10.3390/ani11113288