
Influence of Zinc and Manganese Nanoparticles on Selected Parameters of Turkey Spermatozoa Stored in a Liquid State at 4 °C

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Nanoparticle Suspensions
2.2. Determination of Sperm Concentration
2.3. Determination of Sperm Motility in the CASA System
2.4. Determination of Plasma Membrane Integrity (SYBR-14/PI)
2.5. Determination of Mitochondrial Membrane Potential (JC-1/PI)
2.6. Determination of Nitric Oxide Production
2.7. Determination of Superoxide Dismutase Activity
2.8. Total Protein Content
2.9. Statistical Analysis
3. Results
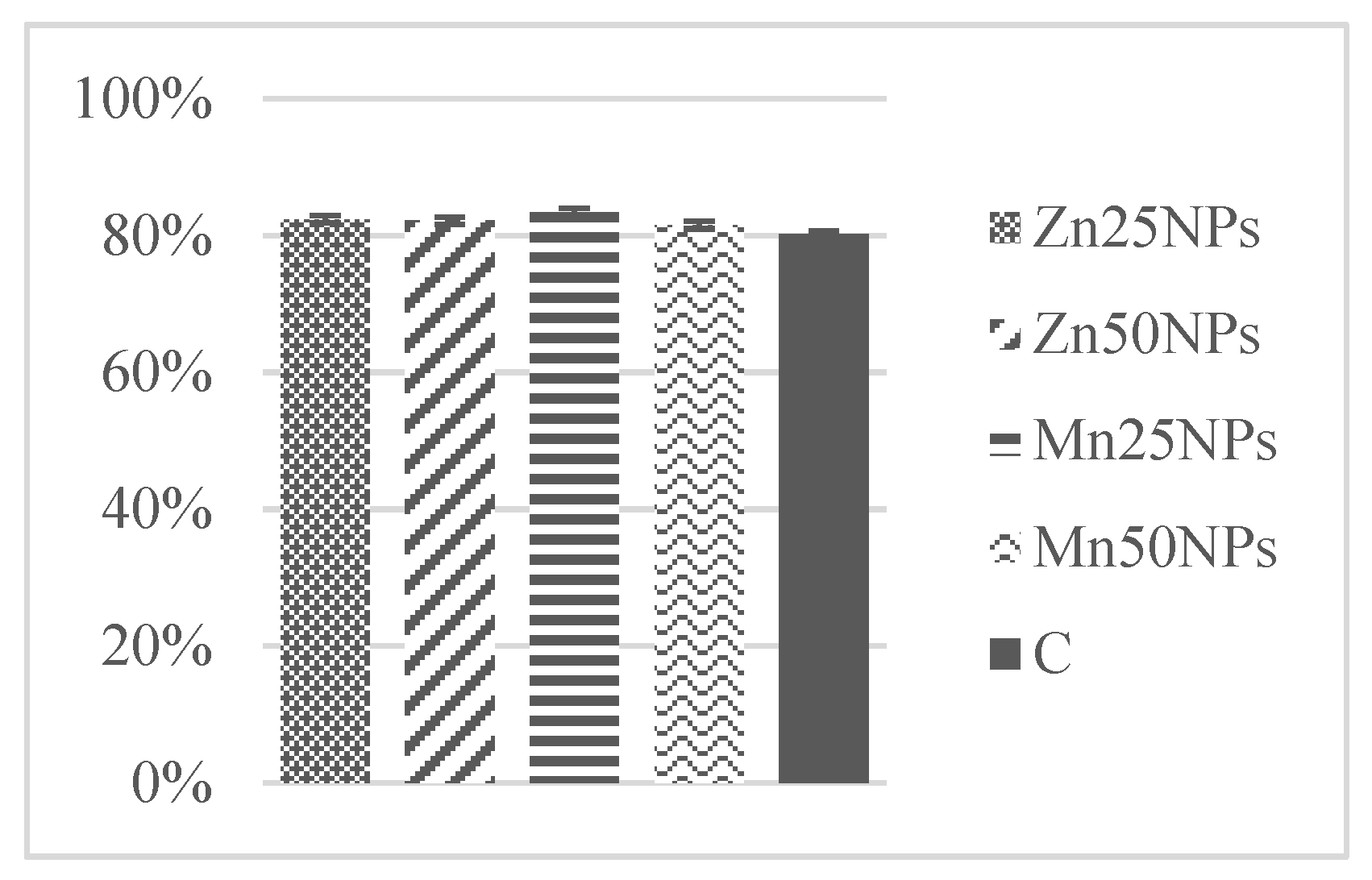
3.1. Sperm Concentration
3.2. Sperm Motility Analysis in the CASA System
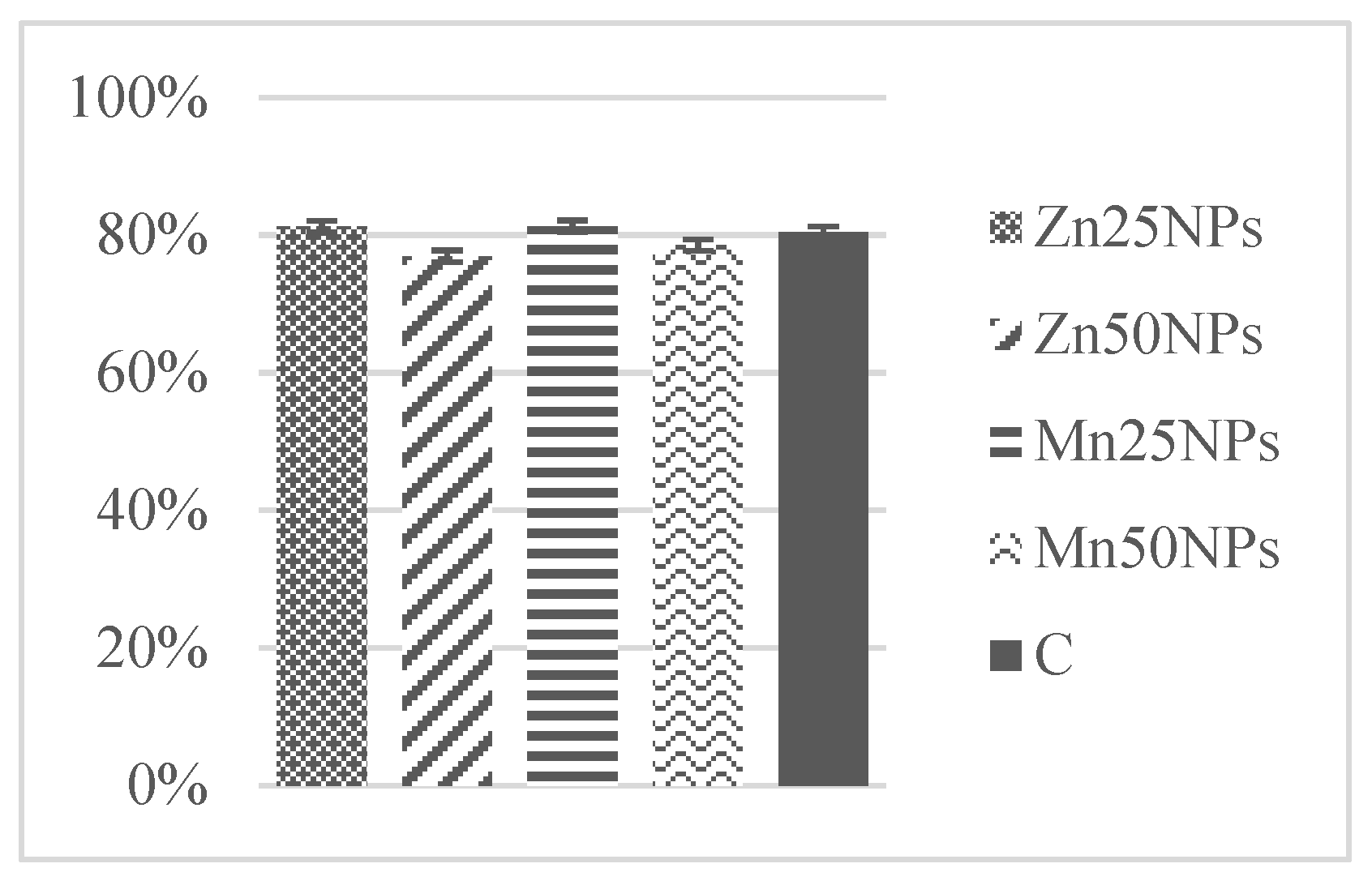
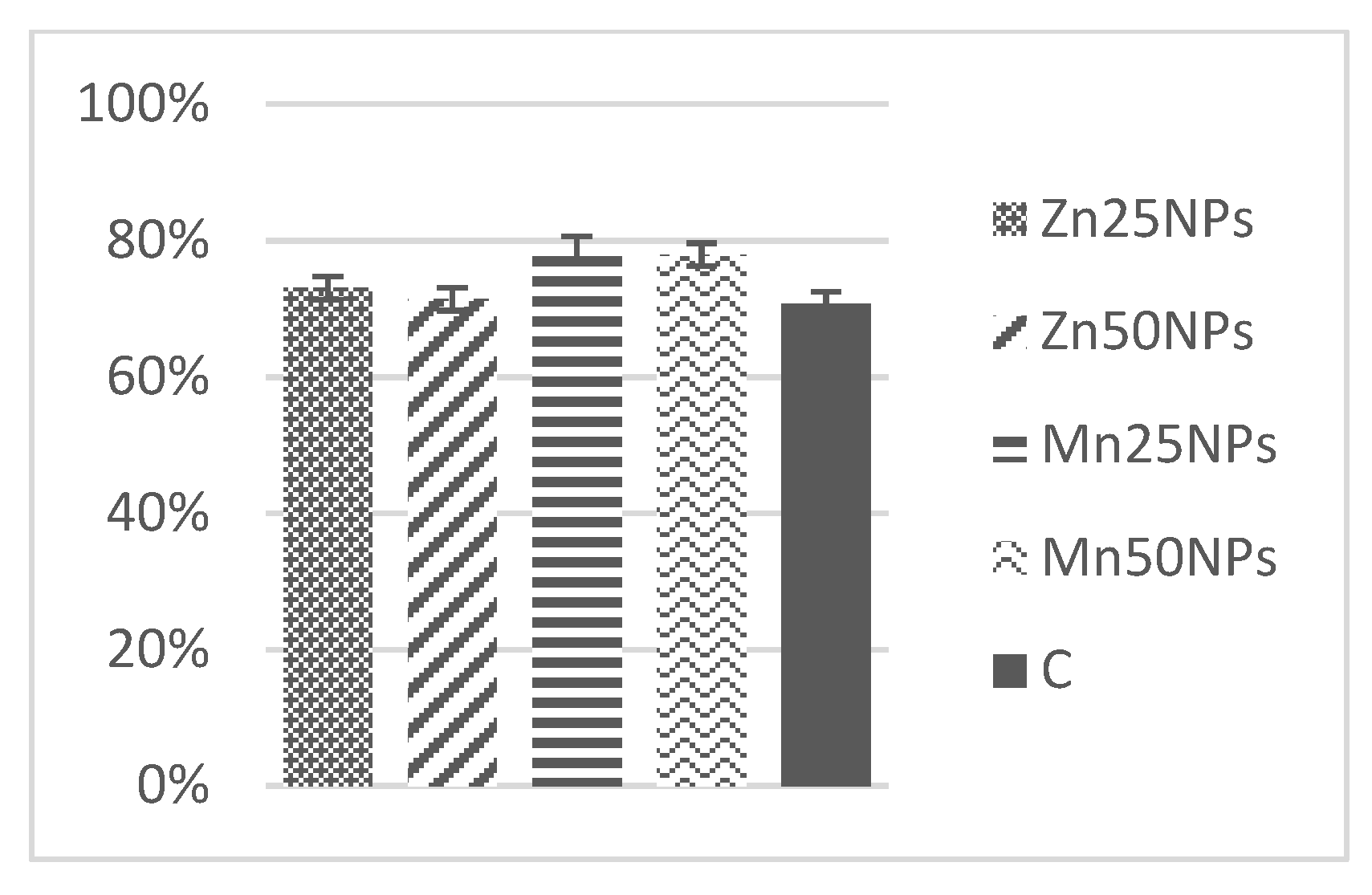
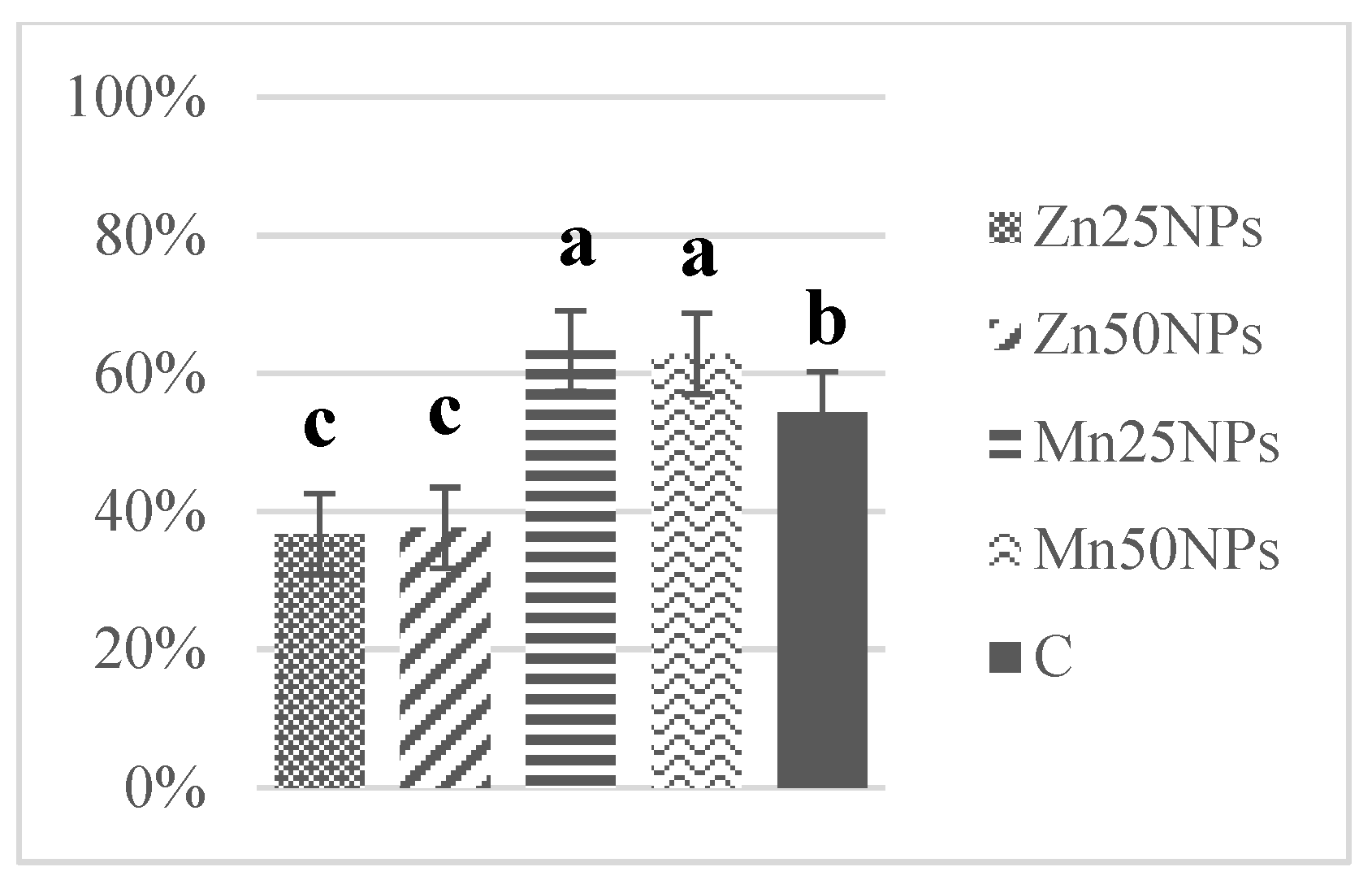
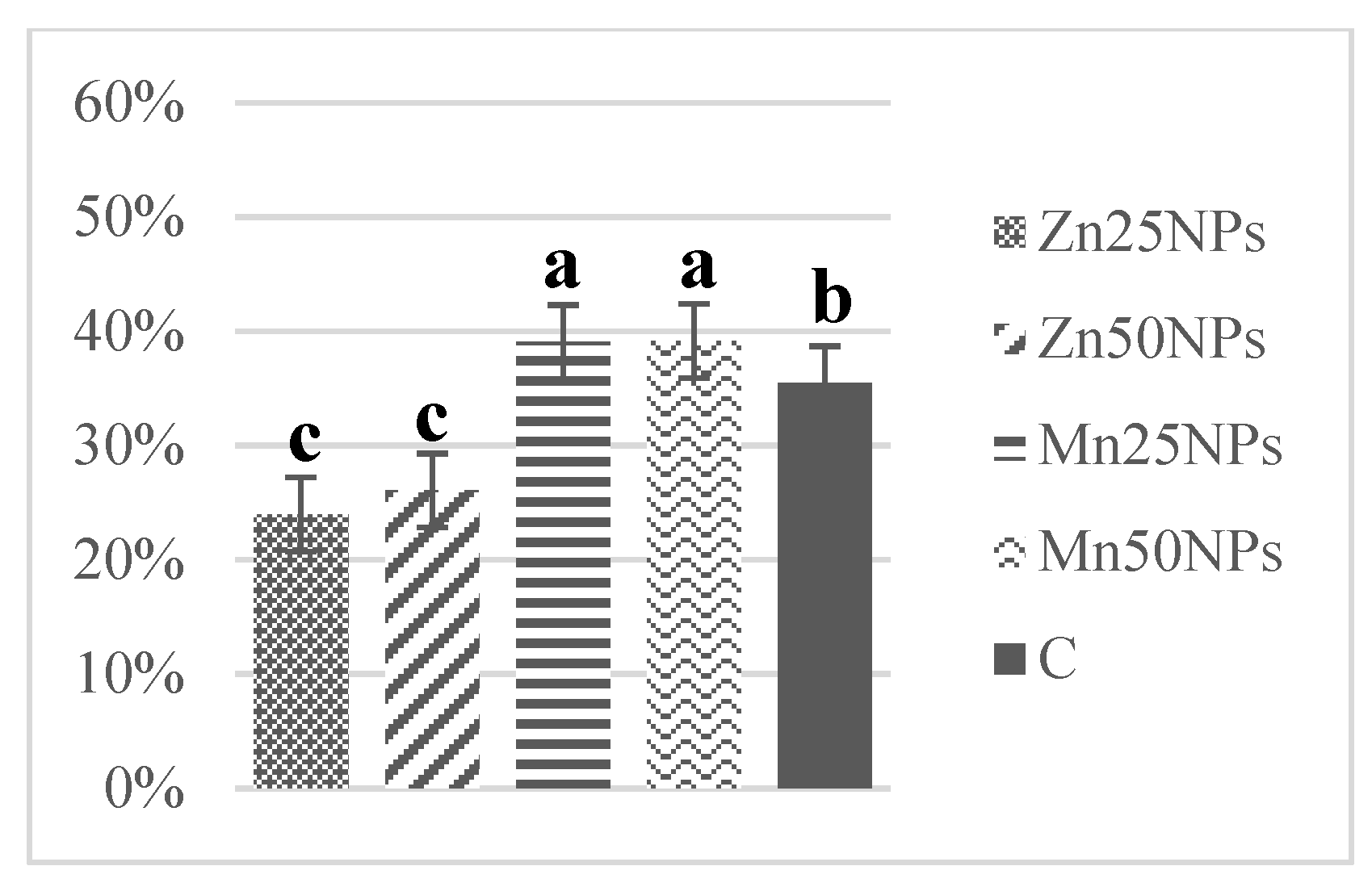
3.3. Plasma Membrane Integrity (SYBR-14/PI)
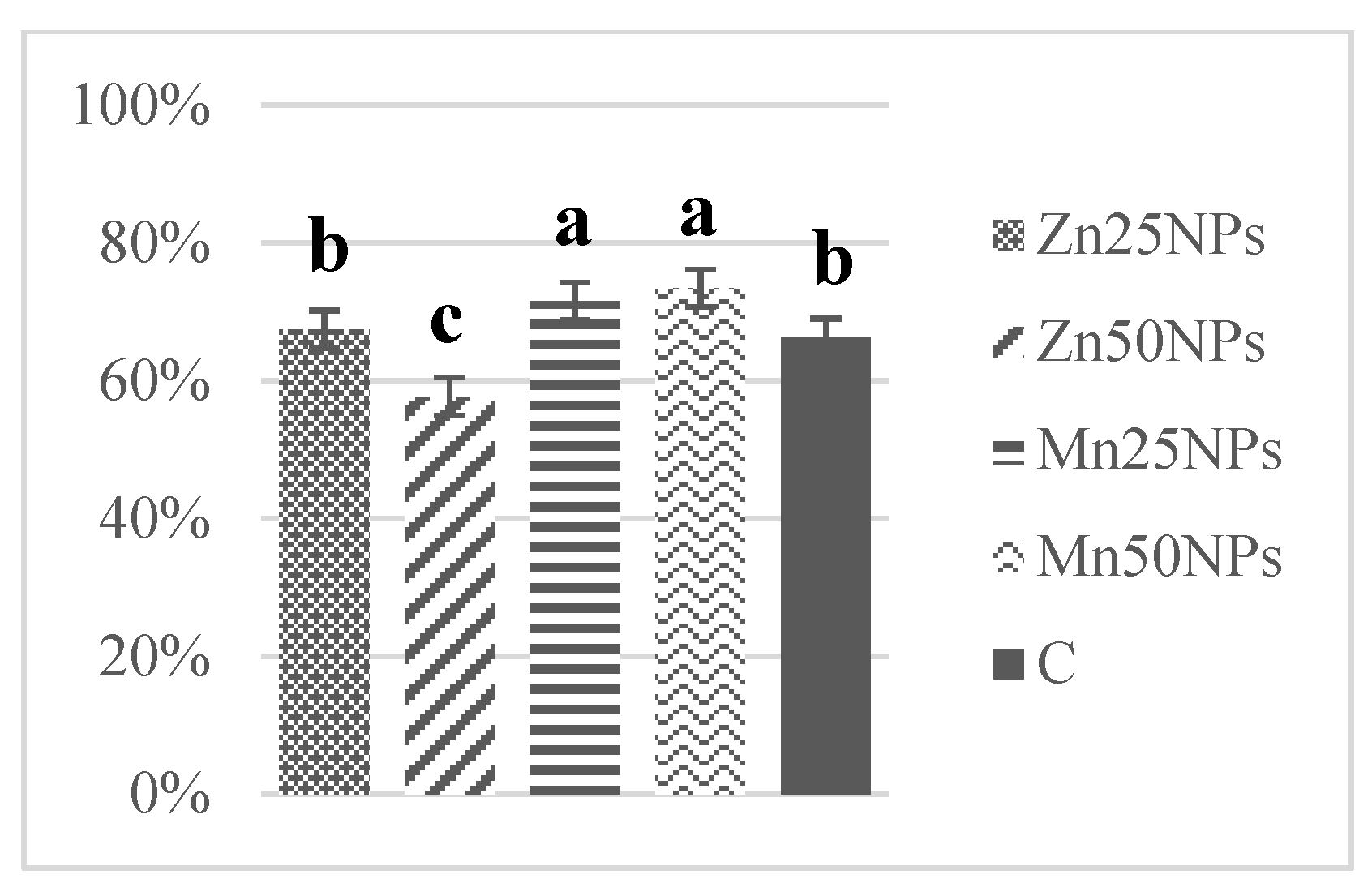
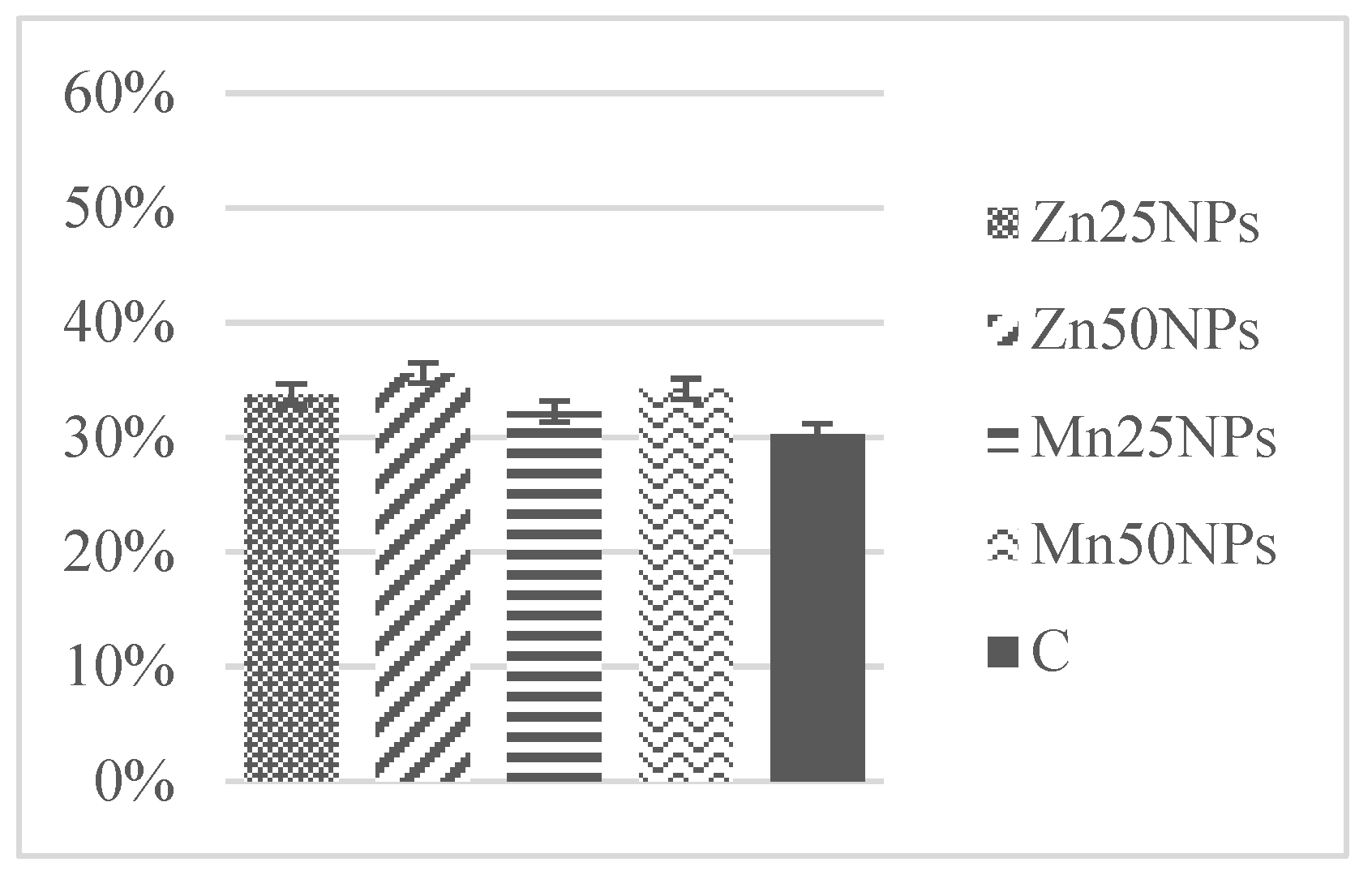
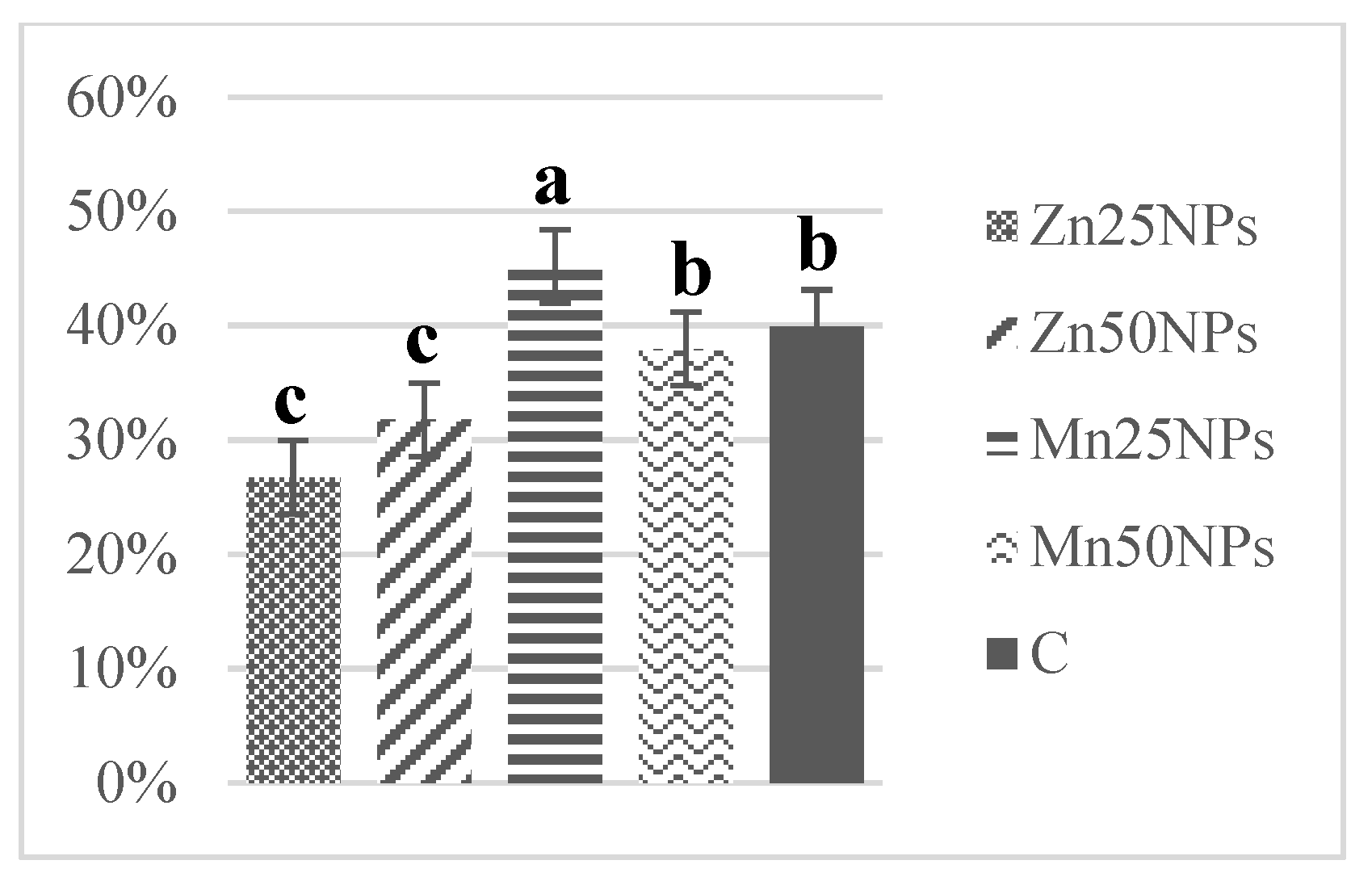
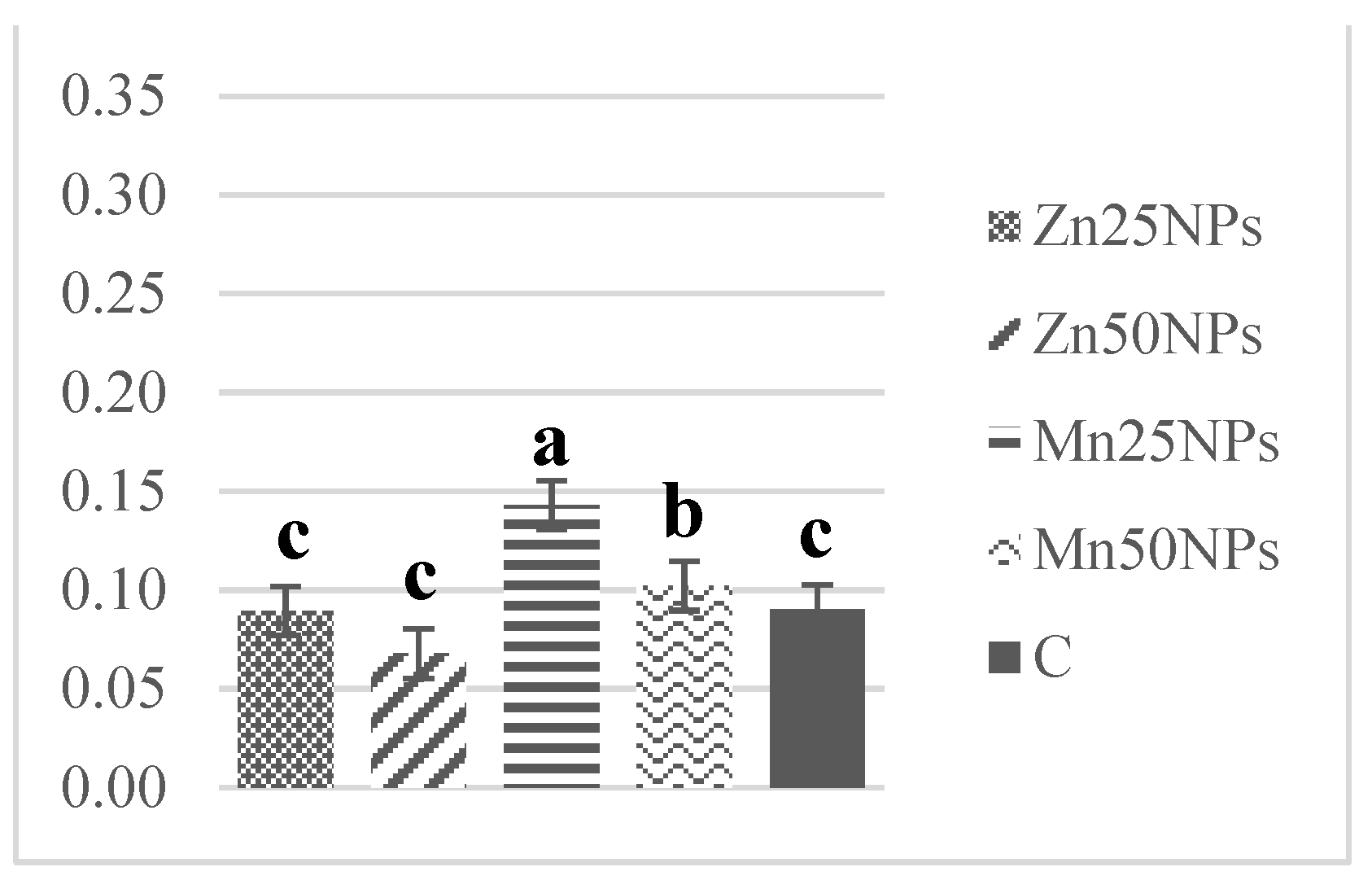
3.4. Mitochondrial Membrane Potential (JC-1/PI)
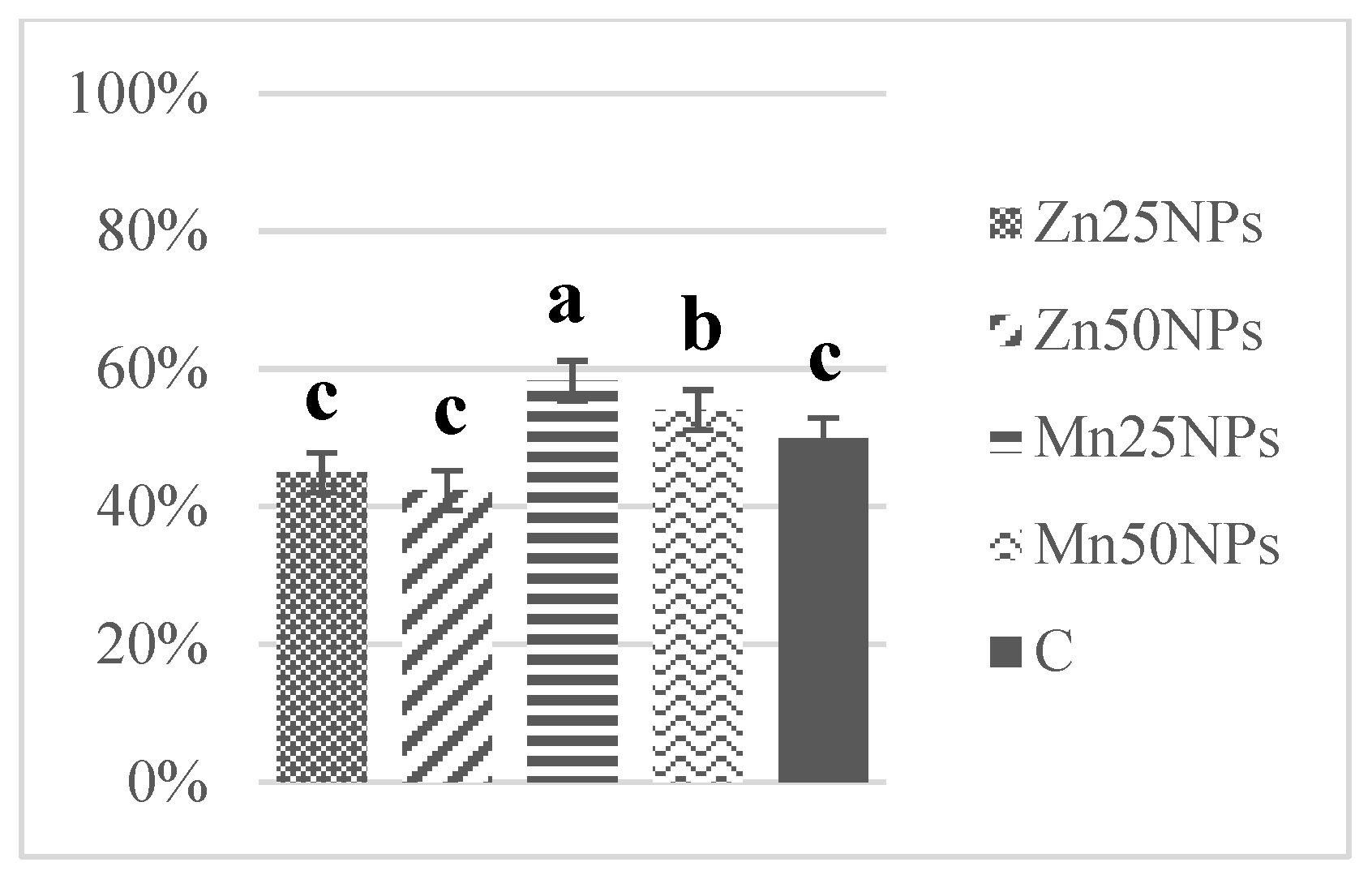
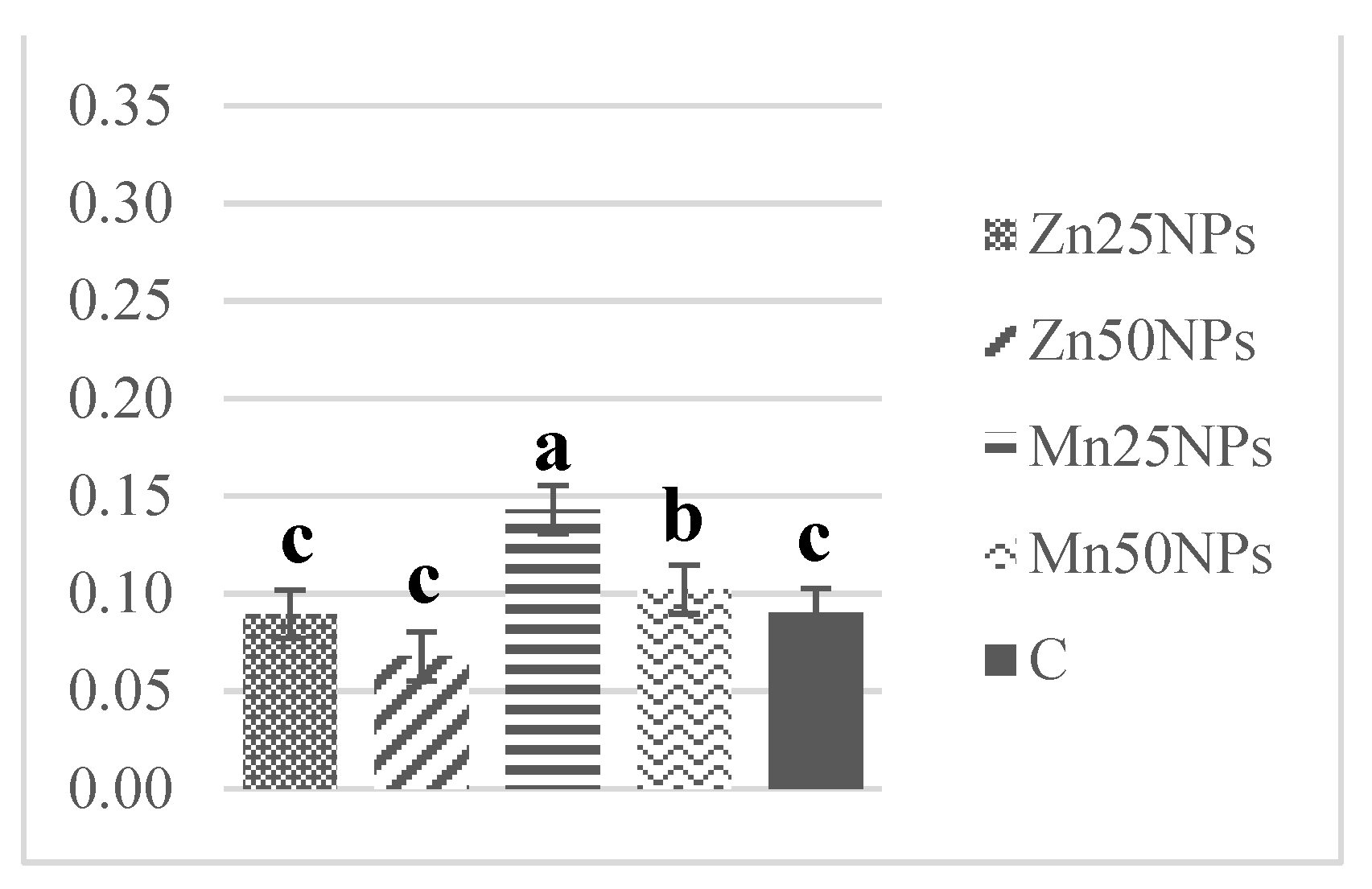
3.5. Nitric Oxide Production
3.6. Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, E.K.; Li, J. Current and future prospects for nanotechnology in animal production. J. Anim. Sci. Biotechnol. 2017, 8, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, M.; Fielding, G.; Bandyopadhyay, A.; Susmita, B. Effects of Zinc and Strontium Substitution in Tricalcium Phosphate on Osteoclast Differentiation and Resorption. Biomater. Sci. 2013, 1, 10. [Google Scholar] [CrossRef]
- Mahboub, H.H.; Shahin, K.; Zaglool, A.W.; Roushdy, E.M.; Ahmed, S.A.A. Efficacy of nano zinc oxide dietary supplements on growth performance, immunomodulation and disease resistance of African catfish Clarias gariepinus. Dis. Aquat. Organ. 2020, 142, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Eggert-Kruse, W.; Zwick, E.M.; Batschulat, K.; Rohr, G.; Armbruster, P.; Petzoldt, D.; Strowitzki, T. Are zinc levels in seminal plasma associated with seminal leukocytes and other determinants of semen quality? Fertil. Steril. 2002, 77, 260–269. [Google Scholar] [CrossRef]
- Isaac, M.; Raibut, E.; Cepeda, C.; Roux, A.; Boturyn, D.; Eliseeva, S.V.; Petoud, S.; Sénèque, O. Luminescent Zinc Fingers: Zn-Responsive Neodymium Near-Infrared Emission in Water. Chemistry 2017, 23, 10992–10996. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Bornhorst, J.; Aschner, M. Manganese metabolism in humans. Front. Biosci. (Landmark Ed.) 2018, 23, 1655–1679. [Google Scholar] [CrossRef] [Green Version]
- Bansal, A.K.; Bilaspuri, G.S. Effect of ferrous sulphate and ascorbic acid on motility, viability and lipid peroxidation of crossbred cattle bull spermatozoa. Animal 2008, 2, 100–104. [Google Scholar] [CrossRef] [Green Version]
- Bansal, A.K.; Bilaspuri, G.S. Oxidative stress alters membrane sulfhydryl status, lipid and phospholipid contents of crossbred cattle bull spermatozoa. Anim. Reprod. Sci. 2008, 104, 398–404. [Google Scholar] [CrossRef]
- Bansal, A.K. Manganese: A potent antioxidant in semen. Iran. J. Appl. Anim. Sci. 2013, 3, 217–221. [Google Scholar]
- Basioura, A.; Tsakmakidis, I.A.; Martinez, E.A.; Roca, J.; Li, J.; Molina, M.F.; Theodoridis, A.; Boscos, C.M.; Parilla, I. Effect of astaxanthin in extenders on sperm quality and functional variables of frozen-thawed boar semen. Anim. Reprod. Sci. 2020, 218, 106478. [Google Scholar] [CrossRef]
- Garner, D.L.; Johnson, L.A. Viability assessment of mammalian sperm using SYBR-14 and propidium iodide. Biol. Reprod. 1995, 53, 276–284. [Google Scholar] [CrossRef]
- Fraser, L.; Lecewicz, M.; Strzeżek, J. Fluorometric assessments of viability and mitochondrial status of boar spermatozoa following liquid storage. Pol. J. Vet. Sci. 2002, 5, 85–92. [Google Scholar] [PubMed]
- Thomas, C.A.; Garner, D.L.; DeJarnette, J.M.; Marshall, C.E. Effect of cryopreservation of bovine sperm organelle function and viability as determined by flow cytometry. Biol. Reprod. 1998, 58, 786–793. [Google Scholar] [CrossRef] [Green Version]
- Lampiao, F.; Strijdom, H.; Du Plessis, S.S. Direct nitric oxide measurement in human spermatozoa: Flow cytometric analysis using the fluorescent probe, diaminofluorescein. Int. J. Andro. 2006, 29, 564–567. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Iaffaldano, N.; Di Iorio, N.; Miranda, M.; Zaniboni, L.; Manshisi, A.; Cerolini, S. Cryopreserving turkey semen in straws and nitrogen vapour using DMSO or DMA: Effects of cryoprotectant concentration, freezing rate and thawing rate on post-thaw semen quality. Br. Poult. Sci. 2016, 57, 264–270. [Google Scholar] [CrossRef]
- Rosato, M.P.; Centoducati, G.; Santacroce, M.P.; Iaffaldano, N. Effects of lycopene on in vitro quality and lipid peroxidation in refrigerated and cryopreserved turkey spermatozoa. Br. Poult. Sci. 2012, 53, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Falchi, L.; Khalil, W.A.; Hassan, M.; Marei, W.F. A Perspectives of nanotechnology in male fertility and sperm function. Int. J. Vet. Sci. Med. 2018, 6, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Stanley, S. Biological nanoparticles and their influence on organisms. Curr. Opin. Biotechnol. 2014, 28, 69–74. [Google Scholar] [CrossRef]
- Mirnamniha, M.; Faroughi, F.; Tahmasbpour, E.; Ebrahimi, P.; Harchegani, A.B. An overview on role of some trace elements in human reproductive health, sperm function and fertilization process. Rev. Environ. Health 2019, 34, 339–348. [Google Scholar] [CrossRef]
- Ahmed, W.M.; El-Tohamy, M.M. Zinc profile in blood and semen of breeding buffalo-bull with particular emphasis on age variation and semen characteristics. In Proceedings of the 5th World Buffalo Congress, Royal Palace, Caserta, Italy, 13–16 October 1997. [Google Scholar]
- Halo, M., Jr.; Bułka, K.; Antos, P.A.; Greń, A.; Slanina, T.; Ondruškae, L.; Tokárová, K.; Massányi, M.; Formicki, G.; Halo, M.; et al. The effect of ZnO nanoparticles on rabbit spermatozoa motility and viability parameters in vitro. Saudi. J. Biol. Sci. 2021. [Google Scholar] [CrossRef]
- Jahanbin, R.; Yazdanshenas, P.; Rahimi, M.; Hajarizadeh, A.; Tvrda, E.; Nazari, S.A.; Mohammadi-Sangcheshmeh, A.; Ghanem, N. In Vivo and In Vitro Evaluation of Bull Semen Processed with Zinc (Zn) Nanoparticles. Biol. Trace Elem. Res. 2021, 199, 126–135. [Google Scholar] [CrossRef]
- Shahin, M.A.; Khalil, W.A.; Saadeldin, I.M.; Swelum, A.A.; El-Harairy, M.A. Comparison between the Effects of Adding Vitamins, Trace Elements, and Nanoparticles to SHOTOR Extender on the Cryopreservation of Dromedary Camel Epididymal Spermatozoa. Animals 2020, 10, 78. [Google Scholar] [CrossRef] [Green Version]
- Zhandi, M.; Talebnia-Chalanbar, A.; Towhidi, A.; Sharafi, M.; Yousefi, A.R.; Hussaini, S.M.H. The effect of zinc oxide on rooster semen cryopreservation. Br. Poult. Sci. 2020, 61, 188–194. [Google Scholar] [CrossRef]
- Chełmońska, B.; Kassner, J. Biologia Rozrodu Zwierząt 2. Biologiczne Uwarunkowania Wartosci Rozrodowej Samca; UWM: Olsztyn, Poland, 2007; pp. 231–260. [Google Scholar]
- Bakst, M.R. Zinc reduces turkey sperm oxygen uptake in vitro. Poult. Sci. 1985, 64, 564–566. [Google Scholar] [CrossRef] [PubMed]
- Bakst, M.R.; Richards, M.P. Concentrations of selected cations in turkey serum and oviductal mucosae. Poult. Sci. 1985, 64, 555–563. [Google Scholar] [CrossRef]
- Zdrojewicz, Z.; Wiśniewska, A. Rola cynku w seksualności mężczyzn. Adv. Clin. Exp. Med. 2005, 14, 1295–1300. [Google Scholar]
- Braun, T. The effect of divalent cations on bovine spermatozoal adenylate cyclase activity. J. Cycl. Nucleotide Res. 1975, 51, 271–281. [Google Scholar]
- Ognik, K.; Krauze, M. The potential for using enzymatic assays to assess the health of turkeys. Poult. Sci. J. 2016, 72, 535–550. [Google Scholar] [CrossRef]
- Bansal, A.K.; Bilaspuri, G.S. Impacts of Oxidative Stress and Antioxidants on Semen Functions. Vet. Med. Int 2010, 2010, 686137. [Google Scholar] [CrossRef] [Green Version]
- Bansal, A.K. Effects of Antioxidants on Crossbred Cattle Bull Spermatozoa Underoxidative Stress. Ph.D. Thesis, Panjab Agricultural University, Ludhiana, India, 2006. [Google Scholar]
- Bansal, A.K.; Cheema, R.S.; Gandotra, V.K. Antioxidant effect of Mn2+ on capacitation and acrosome reaction of fresh and chilled cattle bull semen. Vet. Sci. Dev. 2011, 1, 69–72. [Google Scholar] [CrossRef]
- Cheema, R.S.; Bansal, A.K.; Bilaspuri, G.S. Manganese provides antioxidant protection for sperm cryopreservation that may offer new consideration for clinical fertility. Oxid. Med. Cell. Longev. 2009, 2, 152–159. [Google Scholar] [CrossRef] [Green Version]
- Lapointe, S.; Ahmad, I.; Buhr, M.M.; Sirard, M.A. Modulation of postthaw motility, survival, calcium uptake, and fertility of bovine sperm by magnesium and manganese. J. Dairy Sci. 1996, 79, 2163–2169. [Google Scholar] [CrossRef]
- Eidan, S.M.; Abdulkareem, T.A.; Sultan, O. Influence of Adding Manganese to Tris Extender on some Post-Cryopreservation Semen Attributes of Holstein Bulls. Int. J. App.l Agric. Sci. 2015, 1, 26–30. [Google Scholar] [CrossRef] [Green Version]
- Miriyala, S.; Spasojevic, I.; Tovmasyan, A.; Salvemini, D.; Vujaskovic, Z.; St Clair, D.; Batinic-Haberle, I. Manganese superoxide dismutase, MnSOD and its mimics. Biochim. Biophys. Acta 2012, 1822, 794–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donoghue, A.M.; Sonstegard, T.S.; King, L.M.; Smith, E.J.; Burt, D.W. Turkey sperm mobility influences paternity in the context of competitive fertilization. Biol. Reprod. 1999, 61, 422–427. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; He, Q.; Yan, X.; Cai, Y.; Chen, J. Effect of exogenous nitric oxide on sperm motility in vitro. Biol. Res. 2014, 47, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, S.; Miura, C.; Kikuchi, K.; Celino, F.T.; Agusa, T.; Tanabe, S.; Miura, T. Zinc is an essential trace element for spermatogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 10859–10864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thurston, R.J. Storage of poultry semen above freezing for 24 to 48 h. In Proceedings of the First International Symposium on the Artificial Insemination of Poultry, Savoy, IL, USA, 17–19 June 1994; Bakst, M.R., Wishart, G.J., Eds.; Poultry Science Association: Champaign, IL, USA, 1995; pp. 107–122. [Google Scholar]
- Donoghue, A.M.; Wishart, G.J. Storage of poultry semen. Anim. Reprod Sci. 2000, 62, 213–232. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (h) | Sample | TMOT [%] | PMOT [%] | VAP [µm/s] | VSL [µm/s] | VCL [µm/s] | ALH [µm] | BCF [Hz] | STR [%] | LIN [%} |
|---|---|---|---|---|---|---|---|---|---|---|
| 2 | Zn25 | 32% ± 6% | 27% ± 4% | 72.41 ± 0.89 | 59.39 ± 1.21 | 112.94 ± 2.47 | 3.91 ± 0.23 | 18.67 ± 1.58 | 81% ± 1% | 57% ± 2% |
| Zn50 | 35% ± 6% | 31% ± 4% | 73.41 ± 0.70 | 62.64 ± 1.34 | 111.56 ± 3.07 | 3.86 ± 0.19 | 20.39 ± 1.56 | 84% ± 2% | 60% ± 2% | |
| Mn25 | 44% ± 6% | 31% ± 4% | 73.40 ± 0.89 | 59.12 ± 2.09 | 116.17 ± 2.69 | 3.73 ± 0.24 | 20.83 ± 1.24 | 80% ± 2% | 56% ± 2% | |
| Mn50 | 43% ± 6% | 31% ± 4% | 72.91 ± 1.19 | 59.59 ± 1.57 | 113.61 ± 3.60 | 3.76 ± 0.20 | 21.99 ± 1.52 | 82% ± 2% | 56% ± 4% | |
| C | 47% ± 5% | 31% ± 3% | 73.62 ± 0.86 | 61.90 ± 1.64 | 113.85 ± 2.36 | 3.94 ± 0.14 | 18.35 ± 1.28 | 83% ± 1% | 63% ± 2% | |
| 24 | Zn25 | 20% ± 5% | 14% ± 3% | 48.70 ± 8.39 b | 41.98 ± 7.31 c | 71.87 ± 12.46 b | 2.67 ± 0.41 b | 12.77 ± 2.42 c | 57% ± 10% c | 42% ± 7% c |
| Zn50 | 20% ± 5% | 17% ± 4% | 52.94 ± 8.56 b | 47.90 ± 7.82 b | 76.61 ± 12.64 b | 2.54 ± 0.45 b | 12.46 ± 2.71 c | 63% ± 10% b | 46% ± 8% b | |
| Mn25 | 35% ± 5% | 22% ± 3% | 75.09 ± 1.49 a | 63.31 ± 1.94 a | 112.40 ± 2.88 a | 3.74 ± 0.17 a | 20.49 ± 1.48 a | 84% ± 2% a | 61% ± 2% a | |
| Mn50 | 36% ± 6% | 23% ± 4% | 72.50 ± 0.92 a | 60.41 ± 1.08 a | 111.71 ± 2.91 a | 4.19 ± 0.26 a | 18.09 ± 1.34 b | 83% ± 1% a | 58% ± 2% a | |
| C | 38% ± 6% | 25% ± 3% | 73.68 ± 0.72 a | 61.57 ± 1.27 a | 111.33 ± 2.83 a | 3.71 ± 0.16 a | 18.23 ± 1.35 b | 83% ± 2% a | 60% ± 2% a | |
| 48 | Zn25 | 16% ± 5% b | 12% ± 3% b | 43.27 ± 9.51 b | 37.62 ± 8.37 b | 63.70 ± 14.11 b | 2.22 ± 0.50 b | 11.44 ± 2.76 b | 52% ± 11% b | 37% ± 8% b |
| Zn50 | 10% ± 3% b | 8% ± 3% b | 27.18 ± 9.11 c | 24.07 ± 8.10 c | 38.02 ± 12.73 c | 1.24 ± 0.42 c | 6.56 ± 2.35 b | 33% ± 11% c | 24% ± 8% b | |
| Mn25 | 41% ± 6% a | 30% ± 4% a | 74.93 ± 1.23 a | 64.80 ± 1.65 a | 110.42 ± 2.33 a | 3.57 ± 0.16 a | 18.15 ± 1.28 a | 86% ± 1% a | 63% ± 2% a | |
| Mn50 | 42% ± 6% a | 29% ± 4% a | 73.61 ± 0.77 a | 61.68 ± 1.07 a | 108.19 ± 2.63 a | 3.52 ± 0.12 a | 19.11 ± 1.10 a | 83% ± 1% a | 61% ± 2% a | |
| C | 44% ± 4% a | 31% ± 3% a | 74.59 ± 1.31 a | 64.55 ± 2.15 a | 106.98 ± 1.97 a | 3.41 ± 0.14 a | 17.35 ± 1.44 a | 86% ± 2% a | 64% ± 2% a |
| Time [h] | Total Sperm Motility | Membrane Integrity | Mitochondrial Potential | Nitric Oxide | SOD Activity |
|---|---|---|---|---|---|
| 2 | Zn25NPs | −0.039 | 0.071 | 0.759 * | 0.122 |
| Zn50NPs | −0.252 | 0.213 | 0.746 * | −0.003 | |
| Mn25NPs | 0.193 | −0.004 | 0.826 * | 0.205 | |
| Mn50NPs | 0.008 | 0.286 | 0.633 * | 0.200 | |
| C | −0.435 | 0.407 | 0.174 | 0.301 | |
| 24 | Zn25NPs | −0.032 | 0.576 * | 0.865 * | 0.457 |
| Zn50NPs | 0.085 | 0.738 * | 0.726 * | 0.476 | |
| Mn25NPs | −0.242 | 0.636 * | 0.578 * | 0.666 * | |
| Mn50NPs | 0.080 | 0.786 * | 0.264 | 0.498 | |
| C | −0.111 | 0.442 | 0.403 | 0.504 | |
| 48 | Zn25NPs | −0.039 | 0.092 | 0.591 * | 0.220 |
| Zn50NPs | 0.588 * | −0.180 | 0.618 * | 0.406 | |
| Mn25NPs | 0.614 * | 0.572 * | −0.307 | 0.653 * | |
| Mn50NPs | 0.476 | 0.275 | −0.317 | 0.360 | |
| C | 0.122 | 0.528 * | 0.083 | 0.533 * |
| Time [h] | Membrane Integrity | Total Sperm Motility | Mitochondrial Potential | Nitric Oxide | SOD Activity |
|---|---|---|---|---|---|
| 2 | Zn25NPs | −0.039 | −0.227 | 0.058 | −0.306 |
| Zn50NPs | −0.252 | 0.005 | −0.056 | −0.137 | |
| Mn25NPs | 0.193 | 0.096 | 0.210 | 0.144 | |
| Mn50NPs | 0.008 | −0.300 | 0.312 | −0.384 | |
| C | −0.435 | −0.148 | −0.702 * | −0.036 | |
| 24 | Zn25NPs | −0.032 | 0.169 | −0.088 | −0.106 |
| Zn50NPs | 0.085 | 0.437 | −0.040 | 0.157 | |
| Mn25NPs | −0.242 | −0.036 | −0.333 | −0.108 | |
| Mn50NPs | 0.080 | 0.140 | 0.217 | 0.032 | |
| C | −0.111 | −0.009 | −0.338 | −0.152 | |
| 48 | Zn25NPs | −0.039 | −0.348 | 0.166 | 0.167 |
| Zn50NPs | 0.588 * | −0.161 | 0.229 | 0.205 | |
| Mn25NPs | 0.614 * | 0.045 | 0.136 | 0.318 | |
| Mn50NPs | 0.476 | 0.227 | 0.132 | 0.066 | |
| C | 0.122 | −0.244 | 0.399 | −0.404 |
| Time [h] | Mitochondrial Potential | Total Sperm Motility | Membrane Integrity | Nitric Oxide | SOD Activity |
|---|---|---|---|---|---|
| 2 | Zn25NPs | 0.071 | −0.227 | −0.051 | 0.386 |
| Zn50NPs | 0.213 | 0.005 | −0.098 | 0.677 * | |
| Mn25NPs | −0.004 | 0.096 | −0.084 | 0.521 * | |
| Mn50NPs | 0.286 | −0.300 | −0.024 | 0.631 * | |
| C | 0.407 | −0.148 | −0.306 | 0.711 * | |
| 24 | Zn25NPs | 0.576 * | 0.169 | 0.402 | 0.402 |
| Zn50NPs | 0.738 * | 0.437 | 0.381 | 0.651 * | |
| Mn25NPs | 0.636 * | −0.036 | 0.285 | 0.783 * | |
| Mn50NPs | 0.786 * | 0.140 | −0.093 | 0.595 * | |
| C | 0.442 | −0.009 | −0.413 | 0.706 * | |
| 48 | Zn25NPs | 0.092 | −0.348 | −0.463 | 0.195 |
| Zn50NPs | −0.180 | −0.161 | −0.189 | 0.220 | |
| Mn25NPs | 0.572 * | 0.045 | −0.579 * | 0.805 * | |
| Mn50NPs | 0.275 | 0.227 | −0.720 * | 0.704 * | |
| C | 0.528 * | −0.244 | −0.298 | 0.743 * |
| Time [h] | SOD Activity | Total Sperm Motility | Membrane Integrity | Mitochondrial Potential | Nitric Oxide |
|---|---|---|---|---|---|
| 2 | Zn25NPs | 0.122 | −0.306 | 0.386 | −0.042 |
| Zn50NPs | −0.003 | −0.137 | 0.677 * | −0.210 | |
| Mn25NPs | 0.205 | 0.144 | 0.521 * | 0.233 | |
| Mn50NPs | 0.196 | −0.384 | 0.631 * | −0.259 | |
| C | 0.301 | −0.036 | 0.711 * | −0.233 | |
| 24 | Zn25NPs | 0.457 | −0.106 | 0.402 | 0.218 |
| Zn50NPs | 0.476 | 0.158 | 0.651 * | 0.195 | |
| Mn25NPs | 0.666 * | −0.108 | 0.683 * | 0.049 | |
| Mn50NPs | 0.498 | 0.033 | 0.595 * | −0.289 | |
| C | 0.504 | −0.152 | 0.706 * | −0.191 | |
| 48 | Zn25NPs | 0.220 | 0.167 | 0.195 | 0.053 |
| Zn50NPs | 0.406 | 0.205 | 0.220 | 0.217 | |
| Mn25NPs | 0.653 * | 0.318 | 0.805 * | −0.567 * | |
| Mn50NPs | 0.360 | 0.066 | 0.704 * | −0.834 * | |
| C | 0.533 * | −0.404 | 0.743 * | −0.273 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orzołek, A.; Rafalska, K.T.; Otowska, W.A.; Kordan, W.; Korzekwa, A.J.; Kozłowski, K. Influence of Zinc and Manganese Nanoparticles on Selected Parameters of Turkey Spermatozoa Stored in a Liquid State at 4 °C. Animals 2021, 11, 3289. https://doi.org/10.3390/ani11113289
Orzołek A, Rafalska KT, Otowska WA, Kordan W, Korzekwa AJ, Kozłowski K. Influence of Zinc and Manganese Nanoparticles on Selected Parameters of Turkey Spermatozoa Stored in a Liquid State at 4 °C. Animals. 2021; 11(11):3289. https://doi.org/10.3390/ani11113289
Chicago/Turabian StyleOrzołek, Aleksandra, Katarzyna T. Rafalska, Wiktoria A. Otowska, Władysław Kordan, Anna J. Korzekwa, and Krzysztof Kozłowski. 2021. "Influence of Zinc and Manganese Nanoparticles on Selected Parameters of Turkey Spermatozoa Stored in a Liquid State at 4 °C" Animals 11, no. 11: 3289. https://doi.org/10.3390/ani11113289
APA StyleOrzołek, A., Rafalska, K. T., Otowska, W. A., Kordan, W., Korzekwa, A. J., & Kozłowski, K. (2021). Influence of Zinc and Manganese Nanoparticles on Selected Parameters of Turkey Spermatozoa Stored in a Liquid State at 4 °C. Animals, 11(11), 3289. https://doi.org/10.3390/ani11113289