
Canine and Feline Epididymal Semen—A Plentiful Source of Gametes


 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
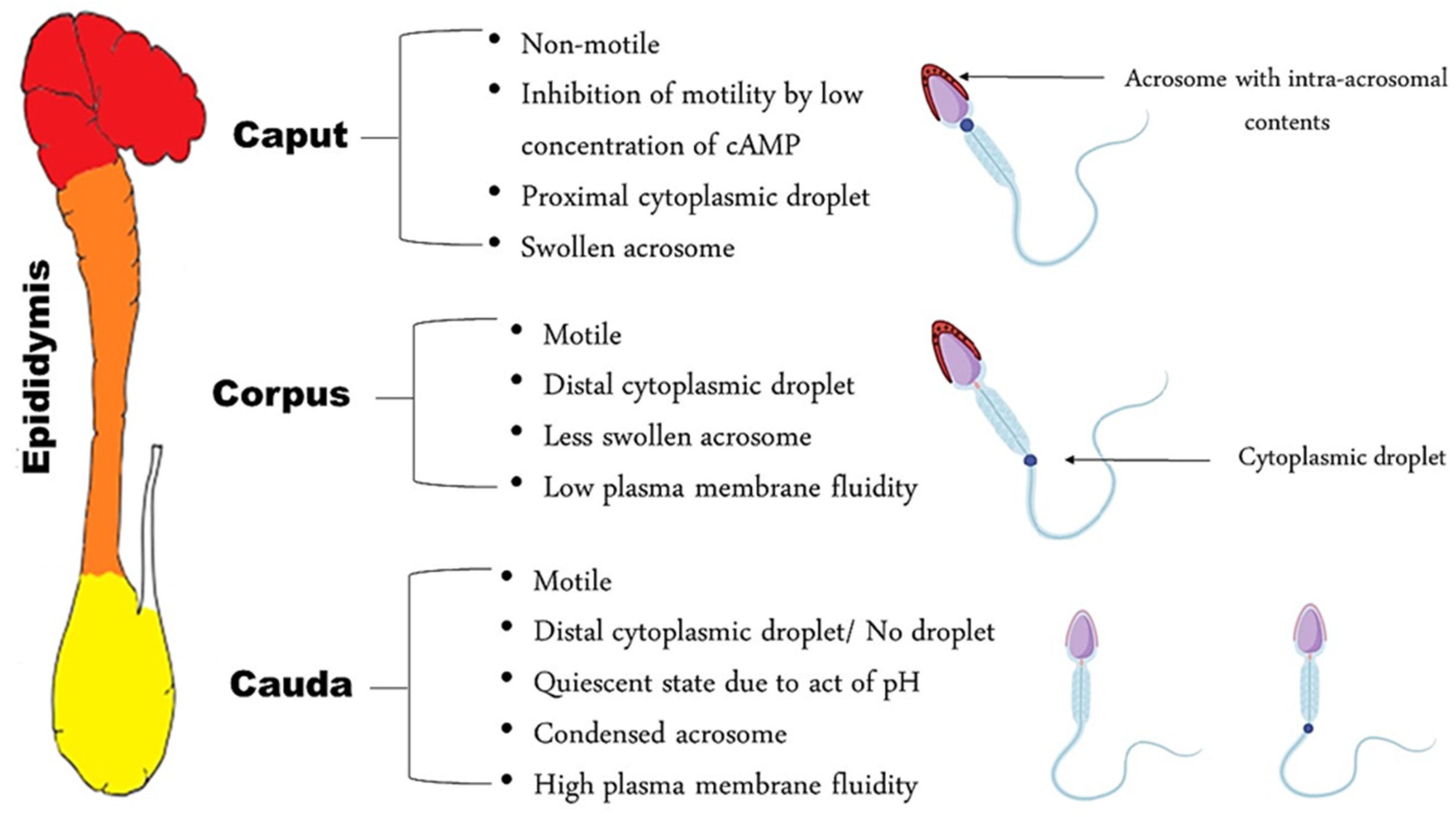
2. Morphological Changes in the Spermatozoa during Epididymal Transit
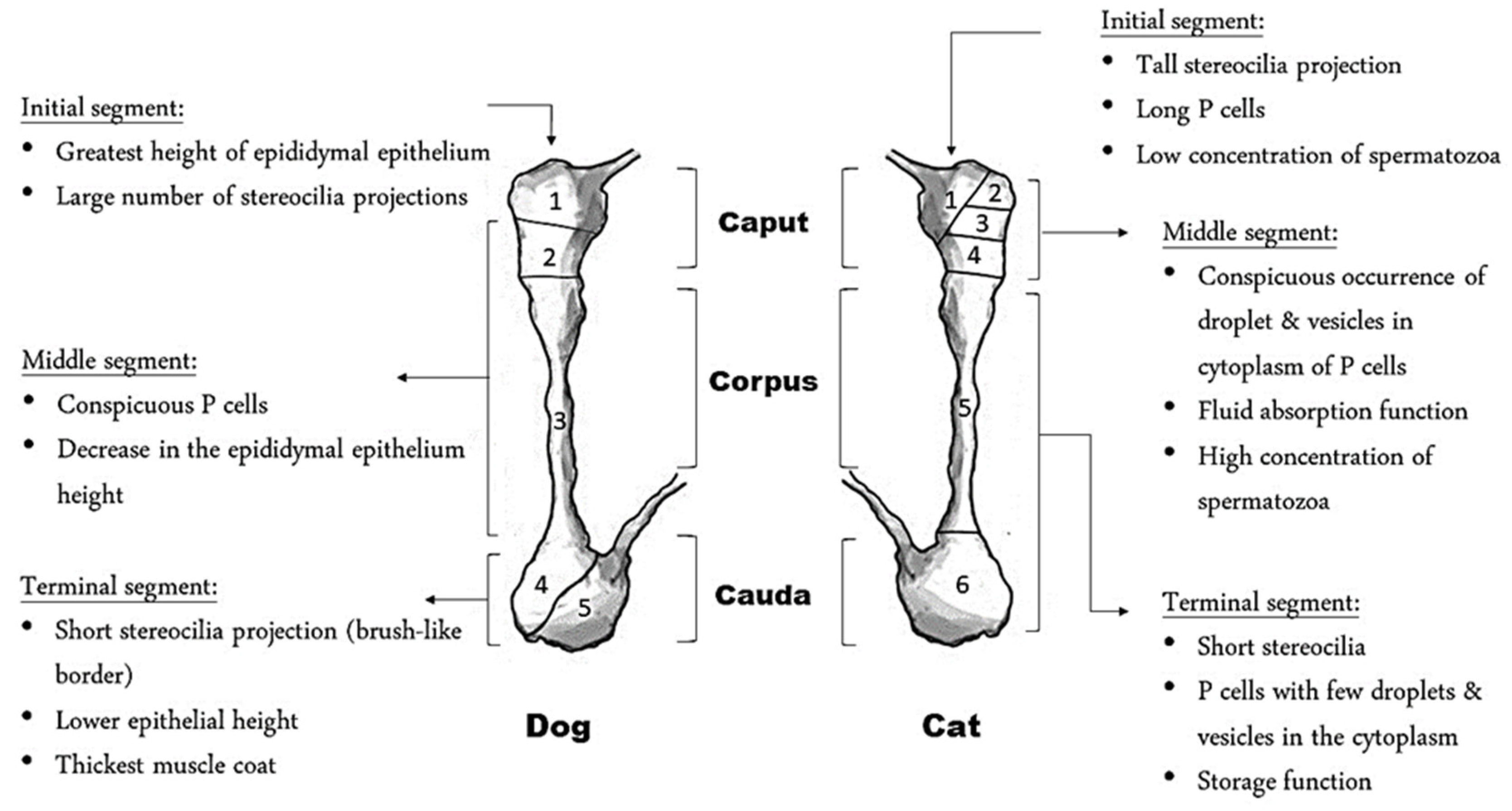
3. Anatomical and Functional Characteristics of the Epididymis
4. Collection of Epididymal Semen in Cats and Dogs
5. Evaluation of Epididymal Sperm Quality
6. Freezability of Epididymal Semen
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Domain, G.; Wydooghe, E. Semen donation and establishment of an open canine semen bank: A novel tool to prevent inbreeding in pedigree dogs. Vlaams Diergeneeskd. Tijdschr. 2019, 88, 55–61. [Google Scholar] [CrossRef]
- Filliers, M.; Rijsselaere, T.; Bossaert, P.; Zambelli, D.; Anastasi, P.; Hoogewijs, M.; Van Soom, A. In vitro evaluation of fresh sperm quality in tomcats: A comparison of two collection techniques. Theriogenology 2010, 74, 31–39. [Google Scholar] [CrossRef]
- Varesi, S.; Vernocchi, V.; Faustini, M.; Luvoni, G.C. Morphological and acrosomal changes of canine spermatozoa during epididymal transit. Acta Vet. Scand. 2013, 55, 17. [Google Scholar] [CrossRef]
- Arrotéia, K.F.; Garcia, P.V. The epididymis: Embryology, structure, function and its role in fertilization and infertility. In Embryology Updates and Highlights on Classic Topics; IntechOpen: London, UK, 2012. [Google Scholar]
- Cooper, T.G. The epididymis, cytoplasmic droplets and male fertility. Asian J. Androl. 2011, 13, 130. [Google Scholar] [CrossRef]
- Angrimani, D.S.R.; Nichi, M. Fatty acid content in epididymal fluid and spermatozoa during sperm maturation in dogs. J. Anim. Sci. Biotechnol. 2017, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Axnér, E. Sperm maturation in the domestic cat. Theriogenology 2006, 66, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Lange-Consiglio, A.; Antonucci, N.; Manes, S.; Corradetti, B.; Cremonesi, F.; Bizzaro, D. Morphometric characteristics and chromatin integrity of spermatozoa in three Italian dog breeds. J. Small Anim. Pract. 2010, 51, 624–627. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.B.; Vandaele, L.; Rijsselaere, T.; Maes, D.; Hoogewijs, M.; Frijters, A.; Noordman, J.; Granados, A.; Dernelle, E.; Shamsuddin, M.; et al. Scrotal insulation and its relationship to abnormal morphology, chromatin protamination and nuclear shape of spermatozoa in Holstein-Friesian and Belgian Blue bulls. Theriogenology 2011, 76, 1246–1257. [Google Scholar] [CrossRef] [PubMed]
- Vernocchi, V.; Morselli, M.G.; Lange-Consiglio, A.; Faustini, M.; Luvoni, G.C. DNA fragmentation and sperm head morphometry in cat epididymal spermatozoa. Theriogenology 2014, 82, 982–987. [Google Scholar] [CrossRef]
- Yoshinaga, K.; Toshimori, K. Organization and modifications of sperm acrosomal molecules during spermatogenesis and epididymal maturation. Microsc. Res. Tech. 2003, 61, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Filho, R.R.D.R.; Angrimani, D.S.R.; Brito, M.M.; Nichi, M.; Vannucchi, C.I.; Lucio, C. Susceptibility of epididymal sperm against reactive oxygen species in dogs. Anim. Biotechnol. 2019, 32, 92–99. [Google Scholar] [CrossRef]
- Kunkitti, P.; Bergqvist, A.-S.; Sjunnesson, Y.; Axnér, E. The ability of feline spermatozoa in different epididymal regions to undergo capacitation and acrosome reaction. Anim. Reprod. Sci. 2015, 161, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Ford, W. Glycolysis and sperm motility: Does a spoonful of sugar help the flagellum go round? Hum. Reprod. Update 2006, 12, 269–274. [Google Scholar] [CrossRef] [PubMed]
- de Sousa Barbosa, B.; Rodrigues Silva, H.V. Morphological and morphometric characterization of domestic cat epididymal sperm. Reprod. Domest. Anim. 2019, 54, 1630–1636. [Google Scholar] [CrossRef] [PubMed]
- Angrimani, D.S.R.; Losano, J.D.A. Role of residual cytoplasm on oxidative status during sperm maturation in dogs. Anim. Reprod. Sci. 2014, 151, 256–261. [Google Scholar] [CrossRef]
- Chłopik, A.; Wysokińska, A. Canine spermatozoa—what do we know about their morphology and physiology? An overview. Reprod. Domest. Anim. 2020, 55, 113–126. [Google Scholar] [CrossRef]
- Kunkitti, P.; Chatdarong, K. Osmotic tolerance of feline epididymal spermatozoa. Anim. Reprod. Sci. 2017, 185, 148–153. [Google Scholar] [CrossRef]
- Toyonaga, M.; Morita, M.; Hori, T.; Tsutsui, T. Distribution of glycoproteins on feline testicular sperm, epididymal sperm and ejaculated sperm. J. Vet.-Med. Sci. 2011, 73, 827–829. [Google Scholar] [CrossRef]
- Vernocchi, V.; Morselli, M.G. Sperm ubiquitination in epididymal feline semen. Theriogenology 2014, 82, 636–642. [Google Scholar] [CrossRef]
- England, G. Dog Breeding, Whelping and Puppy Care; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Greer, M.L. Canine Reproduction and Neonatology; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Sullivan, R.; Mieusset, R. The human epididymis: Its function in sperm maturation. Hum. Reprod. Update 2016, 22, 574–587. [Google Scholar] [CrossRef]
- Yonezawa, A.; Kawamura, S.; Ando, R.; Tadano, T.; Nobunaga, T.; Kimura, Y. Diminution of ejaculatory capacity induced by frequent ejaculation in dogs: Prevention and reversal by yohimbine. Andrologia 1991, 23, 71–74. [Google Scholar] [CrossRef]
- Nicander, L.; Glover, T.D. Regional histology and fine structue of the epididymal duct in the golden hamster (Mesocricetus auratus). J. Anat. 1973, 114, 347–364. [Google Scholar]
- Johnson, L.; Welsh, T.H., Jr. Anatomy and Physiology of the Male Reproductive System and Potential Targets of Toxicants; Elsevier: Amsterdam, The Netherlands, 2010. [Google Scholar]
- Caballero, J.; Frenette, G.; Sullivan, R. Post testicular sperm maturational changes in the bull: Important role of the epididymosomes and prostasomes. Vet. Med. Int. 2011, 2011, 757194. [Google Scholar] [CrossRef]
- Rowlison, T.; Ottinger, M.A.; Comizzoli, P. Key factors enhancing sperm fertilizing ability are transferred from the epididymis to the spermatozoa via epididymosomes in the domestic cat model. J. Assist. Reprod. Genet. 2018, 35, 221–228. [Google Scholar] [CrossRef]
- Sullivan, R.; Frenette, G.; Girouard, J. Epididymosomes are involved in the acquisition of new sperm proteins during epididymal transit. Asian J. Androl. 2007, 9, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, R.; Saez, F. Epididymosomes, prostasomes, and liposomes: Their roles in mammalian male reproductive physiology. Reproduction 2013, 146, R21–R35. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; De Iuliis, G.N. Characteristics of the epididymal luminal environment responsible for sperm maturation and storage. Front. Endocrinol. 2018, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.W.; Asa, C.S.; Wang, C.; Vansandt, L.; Bauman, K.; Callahan, M.; Jens, J.K.; Ellinwood, N.M. Effect of semen collection method on sperm motility of gray wolves (Canis lupus) and domestic dogs (C. l. familiaris). Theriogenology 2011, 76, 975–980. [Google Scholar] [CrossRef]
- Simons, N.; Temmerman, R. Collection and Cryopreservation of Epididymal Spermatozoa in Dogs. Master Thesis, Ghent University, Ghent, Belgium, 2018. [Google Scholar]
- Varesi, S.; Vernocchi, V. Quality of canine spermatozoa retrieved by percutaneous epididymal sperm aspiration. J. Small Anim. Pract. 2013, 54, 87–91. [Google Scholar] [CrossRef]
- Woldringh, G.; Horvers, M. Follow-up of children born after icsi with epididymal spermatozoa. Hum. Reprod. 2011, 26, 1759–1767. [Google Scholar] [CrossRef] [PubMed]
- Saade, R.D.; Neves, P.A. Quantitative (stereological) and qualitative study of rat epididymis after vasectomy and percutaneous epididymal sperm aspiration. J. Urol. 2008, 179, 381–384. [Google Scholar] [CrossRef]
- Attia, K.; Zaki, A. Anti-sperm antibodies and seminal characteristics after testicular biopsy or epididymal aspiration in dogs. Theriogenology 2000, 53, 1355–1363. [Google Scholar] [CrossRef]
- Angrimani, D.S.R.; Losano, J.D.A. Cytoplasmic droplet acting as a mitochondrial modulator during sperm maturation in dogs. Anim. Reprod. Sci. 2017, 181, 50–56. [Google Scholar] [CrossRef]
- Prinosilova, P.; Rybar, R.; Zajicova, A.; Hlavicova, J. DNA integrity in fresh, chilled and frozen-thawed Canine Spermatozoa. Czech Acad. Agric. Sci. 2012, 57, 133–142. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Sartorius, G.; Pletscher, F.; De Geyter, M.; Zhang, H.; De Geyter, C. Isolation of spermatozoa with low levels of fragmented DNA with the use of flow cytometry and sorting. Fertil. Steril. 2013, 100, 686–694.e4. [Google Scholar] [CrossRef]
- Kumaresan, A.; Das Gupta, M.; Datta, T.K.; Morrell, J.M. Sperm DNA Integrity and Male Fertility in Farm Animals: A Review. Front. Vet. Sci. 2020, 7, 321. [Google Scholar] [CrossRef]
- Evenson, D.P.; Larson, K.L.; Jost, L.K. Sperm chromatin structure assay: Its clinical use for detecting sperm DNA fragmentation in male infertility and comparisons with other techniques. J. Androl. 2002, 23, 25–43. [Google Scholar] [CrossRef]
- Niżański, W.; Partyka, A. Flow cytometric, computer assisted and traditional sperm analysis in fertile and subfertile dogs. Sci. Bull. Lviv. Natl. Univ. Vet. Med. Biotechnol. Named SZ Gzhytsky 2011, 13(4-1(50)), 537–538. [Google Scholar]
- Kunkitti, P.; Sjödahl, A.; Bergqvist, A.; Johannisson, A.; Axnér, E. Comparison of DNA Fragmentation Assay in Frozen-Thawed Cat Epididymal Sperm. Reprod. Domest. Anim. 2016, 51, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Van Soom, A.; Rijsselaere, T.; Filliers, M. Cats and dogs: Two neglected species in this era of embryo production in vitro? Reprod. Domest. Anim. 2014, 49, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Hori, T.; Ichikawa, M.; Kawakami, E.; Tsutsui, T. Artificial Insemination of Frozen Epididymal Sperm in Beagle Dogs. J. Vet. Med. Sci. 2004, 66, 37–41. [Google Scholar] [CrossRef][Green Version]
- Hori, T.; Hagiuda, K.; Kawakami, E.; Tsutsui, T. Unilateral intrauterine insemination with prostatic fluid-sensitized frozen caudal epididymal sperm in beagle dogs. Theriogenology 2005, 63, 1573–1583. [Google Scholar] [CrossRef]
- Klinc, P.; Majdic, G.; Sterbenc, N.; Cebulj-Kadunc, N.; Butinar, J.; Kosec, M. Establishment of a Pregnancy Following Intravaginal Insemination with Epididymal Semen from a Dog Castrated due to Benign Prostatic Hyperplasia. Reprod. Domest. Anim. 2005, 40, 559–561. [Google Scholar] [CrossRef] [PubMed]
- Wydooghe, E.; Snoeck, F.; Van Soom, A. Establishment of live birth following intravaginal artificial insemination with chilled epididymal dog semen collected postmortem: Case report. In Proceedings of the 8th International Symposium on Canine and Feline Reproduction ISCFR, Paris, France, 25 June 2016. [Google Scholar]
- Pope, C. Embryo technology in conservation efforts for endangered felids. Theriogenology 2000, 53, 163–174. [Google Scholar] [CrossRef]
- Tsutsui, T.; Wada, M.; Anzai, M.; Hori, T. Artificial Insemination with Frozen Epididymal Sperm in Cats. J. Vet. Med. Sci. 2003, 65, 397–399. [Google Scholar] [CrossRef] [PubMed]
- Kunkitti, P.; Axnér, E.; Bergqvist, A.-S.; Sjunnesson, Y. In vitro fertilization using frozen-thawed feline epididymal spermatozoa from corpus and cauda regions. Theriogenology 2016, 86, 1403–1408. [Google Scholar] [CrossRef] [PubMed]
- Bogliolo, L.; Leoni, G.; Ledda, S.; Naitana, S.; Zedda, M.T.; Carluccio, A.; Pau, S. Intracytoplasmic sperm injection of in vitro matured oocytes of domestic cats with frozen-thawed epididymal spermatozoa. Theriogenology 2001, 56, 955–967. [Google Scholar] [CrossRef]
- Said, T.; Agarwal, A.; Sharma, R.; Thomas, A.J.; Sikka, S.C. Impact of sperm morphology on DNA damage caused by oxidative stress induced by β-nicotinamide adenine dinucleotide phosphate. Fertil. Steril. 2005, 83, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Khodamoradi, K.; Kuchakulla, M.; Narasimman, M.; Khosravizadeh, Z.; Ali, A.; Brackett, N.; Ibrahim, E.; Ramasamy, R. Laboratory and clinical management of leukocytospermia and hematospermia: A review. Ther. Adv. Reprod. Health 2020, 14. [Google Scholar] [CrossRef]
- Rijsselaere, T.; Soom, V.A. Effect of blood admixture on in vitro survival of chilled and frozen–thawed canine spermatozoa. Theriogenology 2004, 61, 1589–1602. [Google Scholar] [CrossRef]
- Peña, A.; Barrio, M. Infertility in a dog due to proximal cytoplasmic droplets in the ejaculate: Investigation of the significance for sperm functionality in vitro. Reprod. Domest. Anim. 2007, 42, 471–478. [Google Scholar] [CrossRef]
- Phillips, T.; Dhaliwal, G. Efficacy of four density gradient separation media to remove erythrocytes and nonviable sperm from canine semen. Theriogenology 2012, 77, 39–45. [Google Scholar] [CrossRef]
- Soygur, B.; Celik, S. Effect of erythrocyte-sperm separation medium on nuclear, acrosomal, and membrane maturity parameters in human sperm. J. Assist. Reprod. Genet. 2018, 35, 491–501. [Google Scholar] [CrossRef]
- Nepal, S.; Feng, H.; Gale, B.K. Optimization of a microfluidic spiral channel used to separate sperm from blood cells. Biomicrofluidics 2020, 14, 064103. [Google Scholar] [CrossRef]
- Martins, M.; Justino, R. Comparison of two different extenders for cryopreservation of epididymal dog sperm. Reprod. Domest. Anim. 2012, 47, 293–294. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, D.; Leahy, R. Cryopreservation of epididymal dog sperm. Anim. Reprod. Sci. 2001, 67, 101–111. [Google Scholar] [CrossRef]
- Peña, F.; Núñez-Martínez, I.; Morán, J. Semen technologies in dog breeding: An update. Reprod. Domest. Anim. 2006, 41, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, T.J.; Morrell, J.M. Artificial Insemination, in Veterinary Reproduction and Obstetrics, 10th ed.; Noakes, D.E.J., Parkinson, T.J., England, G.C.W., Eds.; Bailliere Tindall: Paris, France, 2019; pp. 746–777. [Google Scholar]
- Pan, C.; Wu, Y. Effects of seminal plasma concentration on sperm motility and plasma and acrosome membrane integrity in chilled canine spermatozoa. Pol. J. Vet. Sci. 2018, 21, 133–138. [Google Scholar] [PubMed]
- Korochkina, E.; Johannisson, A. Effect of prostatic fluid on the quality of fresh and frozen-thawed canine epididymal spermatozoa. Theriogenology 2014, 82, 1206–1211. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Different Features | Dog | Cat | References |
|---|---|---|---|
| Total sperm length | 62.7 µm | 58.6 µm | [14,15] |
| Acquisition of motility | In corpus | In caput-corpus junction | [7,16] |
| Motility mechanism | Supported by glycolysis | Supported by endogenous lipids | [7,17] |
| Migration of cytoplasmic droplet to end of midpiece | In corpus | In caput- corpus junction | [3,7] |
| Common Features | References |
|---|---|
| Fluid leaving the testis is reabsorbed in efferent duct and caput of the epididymis | [7,17] |
| Osmotic pressure within the epididymis increases caudally | [17,18] |
| Sperm plasma membrane is composed of large amount of phospholipids, cholesterol and glycoproteins | [6,19] |
| Cholesterol stiffens the sperm plasma membrane and increases its permeability | [6,7] |
| Glycoproteins coating the plasma membrane change in the same manner during epididymal transit | [19] |
| Percentage of spermatozoa with swollen acrosome and incomplete condensed chromatin decreases as sperm migrates distally along the epididymis | [3,15] |
| Presence of selective abnormal spermatozoa removal (phagocytosis) | [7,17] |
| Retained cytoplasmic droplets will be detached at or post-ejaculation | [20] |
| Spermatozoa contain only protamine P1 which renders chromatin relatively stable | [20] |
| Method of ART | Region of Epididymal Sperm | Sperm Dose | Deposition Site | Conception Rate | Cleavage Rate | Reference |
|---|---|---|---|---|---|---|
| AI | cauda | 5 × 107 | unilateral intrauterine | 27.30% | - | [51] |
| AI | cauda | 4 × 107 | unilateral intrauterine | 28.60% | - | [19] |
| AI | cauda | 1 × 107 | Intratubal | 80% | - | [19] |
| (24 hr post hCG injection) | ||||||
| Intratubal | 20% | - | ||||
| (30 hr post hCG injection) | ||||||
| IVF | cauda | 5 × 105 | - | - | 33.33% | [52] |
| IVF | corpus | 5 × 105 | - | - | 32.03% | [52] |
| ICSI | cauda | single spermatozoa | - | - | 82.20% | [53] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali Hassan, H.; Domain, G.; Luvoni, G.C.; Chaaya, R.; Van Soom, A.; Wydooghe, E. Canine and Feline Epididymal Semen—A Plentiful Source of Gametes. Animals 2021, 11, 2961. https://doi.org/10.3390/ani11102961
Ali Hassan H, Domain G, Luvoni GC, Chaaya R, Van Soom A, Wydooghe E. Canine and Feline Epididymal Semen—A Plentiful Source of Gametes. Animals. 2021; 11(10):2961. https://doi.org/10.3390/ani11102961
Chicago/Turabian StyleAli Hassan, Hiba, Guillaume Domain, Gaia Cecilia Luvoni, Rana Chaaya, Ann Van Soom, and Eline Wydooghe. 2021. "Canine and Feline Epididymal Semen—A Plentiful Source of Gametes" Animals 11, no. 10: 2961. https://doi.org/10.3390/ani11102961
APA StyleAli Hassan, H., Domain, G., Luvoni, G. C., Chaaya, R., Van Soom, A., & Wydooghe, E. (2021). Canine and Feline Epididymal Semen—A Plentiful Source of Gametes. Animals, 11(10), 2961. https://doi.org/10.3390/ani11102961