Dietary Phytogenic Combination with Hops and a Mixture of a Free Butyrate Acidifier and Gluconic Acid Maintaining the Health Status of the Gut and Performance in Chickens


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
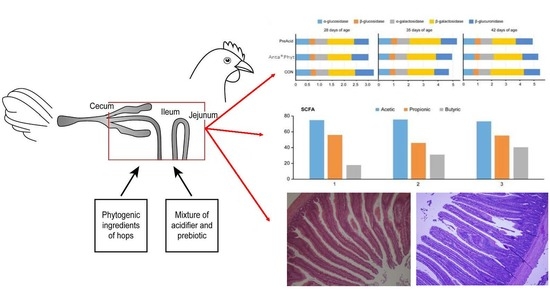
2.1. Birds, Diets, and Experimental Design
2.2. Sampling Procedures
2.3. Analyses of Caecal Microbiota Activity
2.4. Analysis of Gut Morphostructure Indices
2.5. Statistical Analysis
2.6. Ethical Statement
3. Results and Discussion
3.1. Bird Performance
3.2. SCFA Concentrations and Profile
3.3. Bacterial Enzyme Activity
3.4. Gut Morphostructure Indices
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pourabedin, M.; Zhao, X. Prebiotics and gut microbiota in chickens. FEMS Microbiol. Lett. 2015, 362, 122. [Google Scholar] [CrossRef]
- Tsukahara, T.; Koyama, H.; Okada, M.; Ushida, K. Stimulation of Butyrate Production by Gluconic Acid in Batch Culture of Pig Cecal Digesta and Identification of Butyrate-Producing Bacteria. J. Nutr. 2002, 132, 2229–2234. [Google Scholar] [CrossRef]
- Cook, S.I.; Sellin, J.H. Review article: Short chain fatty acids in health and disease. Aliment. Pharmacol. Ther. 1998, 12, 499–507. [Google Scholar] [CrossRef]
- Yeoman, C.J.; Chia, N.; Jeraldo, P.; Sipos, M.; Goldenfeld, N.D.; White, B.A. The microbiome of the chicken gastrointestinal tract. Anim. Health Res. Rev. 2012, 13, 89–99. [Google Scholar] [CrossRef]
- Applegate, T.J.; Klose, V.; Steiner, T.; Ganner, A.; Schatzmayr, G. Probiotics and phytogenics for poultry: Myth or reality? J. Appl. Poult. Res. 2010, 19, 194–210. [Google Scholar] [CrossRef]
- Jamroz, D.; Wertelecki, T.; Houszka, M.; Kamel, C. Influence of diet type on the inclusion of plant origin active substances on morphological and histochemical characteristics of the stomach and jejunum walls in chicken. J. Anim. Physiol. Anim. Nutr. 2006, 90, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi Gheisar, M.M.; Kim, I. Phytobiotics in poultry and swine nutrition—A review. Ital. J. Anim. Sci. 2018, 17, 92–99. [Google Scholar] [CrossRef]
- Alagawany, M.; ElNesr, S.S.; Farag, M.R.; El-Hack, M.E.; Khafaga, A.F.; Taha, A.E.; Tiwari, R.; Yatoo, M.I.; Bhatt, P.; Marappan, G.; et al. Use of licorice (Glycyrrhiza glabra) herb as a feed additive in poultry: Current knowledge and prospects. Animals 2019, 9, 536. [Google Scholar] [CrossRef] [PubMed]
- Cornelison, J.M.; Yan, F.; Watkins, S.E.; Rigby, L.; Segal, J.B.; Waldroup, P.W. Evaluation of hops (Humulus iupulus) as an antimicrobial in broiler diets. Int. J. Poult. Sci. 2006, 5, 134–136. [Google Scholar] [CrossRef]
- Gerhäuser, C. Broad spectrum antiinfective potential of xanthohumol from hop (Humulus lupulus L.) in comparison with activities of other hop constituents and xanthohumol metabolites. Mol. Nutr. Food Res. 2005, 49, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Windisch, W.; Schedle, K.; Plitzner, C.; Kroismayr, A. Use of phytogenic products as feed additives for swine and poultry. J. Anim. Sci. 2008, 86, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Aviagen. Ross 308 Broiler Nutrition Specifications; Aviagen Group: Huntsville, AL, USA, 2014. [Google Scholar]
- Konieczka, P.; Kaczmarek, S.A.; Hejdysz, M.; Kinsner, M.; Szkopek, D.; Smulikowska, S. Effects of faba bean extrusion and phytase supplementation on performance, phosphorus and nitrogen retention and gut microbiota activity in broilers. J. Sci. Food Agric. 2020. [Google Scholar] [CrossRef] [PubMed]
- Konieczka, P.; Mikulski, D.; Ognik, K.; Juśkiewicz, J.; Zduńczyk, Z.; Józefiak, D.; Jankowski, J. Chemically preserved high-moisture corn in the turkey diet does not compromise performance and maintains the functional status of the gut. Anim. Feed Sci. Technol. 2020, 263, 114483. [Google Scholar] [CrossRef]
- Konieczka, P.; Barszcz, M.; Choct, M.; Smulikowska, S. The interactive effect of dietary n-6:n-3 fatty acid ratio and vitamin E level on tissue lipid peroxidation, DNA damage in intestinal epithelial cells, and gut morphology in chickens of different ages. Poult. Sci. 2018, 97, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Guo, Y. Effects of dietary sodium butyrate supplementation on the intestinal morphological structure, absorptive function and gut flora in chickens. Anim. Feed Sci. Technol. 2007, 132, 240–249. [Google Scholar] [CrossRef]
- Jacobs, C.M.; Parsons, C.M. The effect of Grobiotic-P combined with yeast cell wall and gluconic acid on growth performance, nutrient digestibilities, and cecal microbial populations in young chicks. Poult. Sci. 2009, 88, 2360–2367. [Google Scholar] [CrossRef]
- Midilli, M.; Alp, M.; Kocabach, N.; Muglah, O.H.; Turan, N.; Yilmaz, H.; Cakir, S. Effects of dietary probiotic and prebiotic supplementation on growth performance and serum IgG concentration of broilers. S. Afr. J. Anim. Sci. 2008, 38, 21–27. [Google Scholar] [CrossRef]
- Froebel, L.K.; Jalukar, S.; Lavergne, T.A.; Lee, J.T.; Duong, T. Administration of dietary prebiotics improves growth performance and reduces pathogen colonization in broiler chickens. Poult. Sci. 2019, 98, 6668–6676. [Google Scholar] [CrossRef]
- Richards, P.J.; Flaujac Lafontaine, G.M.; Connerton, P.L.; Liang, L.; Asiani, K.; Fish, N.M.; Connerton, I.F. Galacto-Oligosaccharides Modulate the Juvenile Gut Microbiome and Innate Immunity To Improve Broiler Chicken Performance. mSystems 2020, 5, e00827-19. [Google Scholar] [CrossRef]
- Grashorn, M.A. Use of phytobiotics in broiler nutrition–An alternative to infeed antibiotics? J. Anim. Feed Sci. 2010, 19, 338–347. [Google Scholar] [CrossRef]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the gut to the peripheral tissues: The multiple effects of butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ndelekwute, E.K.; Assam, E.D.; Assam, E.M. Apparent nutrient digestibility, gut pH and digesta viscosity of broiler chickens fed acidified water. MOJ Anat. Physiol. 2018, 5, 250–253. [Google Scholar] [CrossRef]
- Ndelekwute, E.K.; Unah, U.L.; Udoh, U.H. Effect of dietary organic acids on nutrient digestibility, faecal moisture, digesta pH and viscosity of broiler chickens. MOJ Anat. Physiol. 2019, 6, 40–43. [Google Scholar] [CrossRef]
- Konieczka, P.; Nowicka, K.; Madar, M.; Taciak, M.; Smulikowska, S. Effects of pea extrusion and enzyme and probiotic supplementation on performance, microbiota activity and biofilm formation in the broiler gastrointestinal tract. Br. Poult. Sci. 2018, 59, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Konieczka, P.; Czerwiński, J.; Jankowiak, J.; Ząbek, K.; Smulikowska, S. Effects of partial replacement of soybean meal with rapeseed meal, narrow-leaved lupin, DDGS, and probiotic supplementation, on performance and gut microbiota activity and diversity in broilers. Ann. Anim. Sci. 2019, 19, 1115–1131. [Google Scholar] [CrossRef]
- Zduńczyk, Z.; Jankowski, J.; Kaczmarek, S.; Juśkiewicz, J. Determinants and effects of postileal fermentation in broilers and turkeys part 1: Gut microbiota composition and its modulation by feed additives. World’s Poult. Sci. J. 2015, 71, 37–48. [Google Scholar] [CrossRef]
- Laudadio, V.; Passantino, L.; Perillo, A.; Lopresti, G.; Passantino, A.; Khan, R.U.; Tufarelli, V. Productive performance and histological features of intestinal mucosa of broiler chickens fed different dietary protein levels. Poult. Sci. 2011, 91, 265–270. [Google Scholar] [CrossRef]
- Hashemi, S.R.; Zulkifli, I.; Davoodi, H.; Hair Bejo, M.; Loh, T.C. Intestinal histomorphology changes and serum biochemistry responses of broiler chickens fed herbal plant (Euphorbia hirta) and mix of acidifier. Iran. J. Appl. Anim. Sci. 2014, 4, 95–103. [Google Scholar]
- Deepa, K.; Purushothaman, M.R.; VasanthaKumar, P.; Sivakumar, K. Butyric acid as an antibiotic substitute for broiler chicken—A review. Adv. Anim. Vet. Sci. 2018, 6, 63–69. [Google Scholar] [CrossRef]
- Emami, N.K.; Daneshmand, A.; Naeini, S.Z.; Graystone, E.N.; Broom, L.J. Effects of commercial organic acid blends on male broilers challenged with E. coli K88: Performance, microbiology, intestinal morphology, and immune response. Poult. Sci. 2017, 96, 3254–3263. [Google Scholar] [CrossRef] [PubMed]
- Miles, R.D.; Butcher, G.D.; Henry, P.R.; Littell, R.C. Effect of antibiotic growth promoters on broiler performance, intestinal growth parameters, and quantitative morphology. Poult. Sci. 2006, 85, 476–485. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Indices | Dietary Treatments 1 | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | Anta®Phyt | PreAcid | |||
| Birds at 28 days of age | |||||
| SCFA concentrations (μmol/g) | |||||
| Acetic | 60.8 ab | 56.4 a | 73.2 b | 2.90 | 0.042 |
| Propionic | 4.47 | 3.42 | 5.04 | 0.353 | 0.165 |
| Iso-butyric | 0.69 | 0.70 | 0.92 | 0.068 | 0.320 |
| Butyric | 15.4 ab | 12.80 a | 19.7 b | 1.160 | 0.040 |
| Iso-valeric | 0.49 | 0.53 | 0.80 | 0.071 | 0.153 |
| Valeric | 1.03 a | 0.97 a | 1.36 b | 0.071 | 0.049 |
| Sum of SCFAs | 82.9 ab | 74.9 a | 101.0 b | 4.113 | 0.022 |
| SCFA profile (% of total SCFAs) | |||||
| Acetic | 74.0 | 75.4 | 72.5 | 0.765 | 0.316 |
| Propionic | 5.46 | 4.70 | 5.04 | 0.363 | 0.719 |
| Butyric | 17.9 | 16.8 | 19.4 | 0.775 | 0.391 |
| Birds at 35 days of age | |||||
| SCFA concentrations (μmol/g) | |||||
| Acetic | 71.4 | 57.9 | 61.6 | 3.524 | 0.277 |
| Propionic | 4.36 | 3.56 | 4.09 | 0.318 | 0.585 |
| Iso-butyric | 0.91 | 0.79 | 1.01 | 0.067 | 0.432 |
| Butyric | 16.8 | 20.4 | 15.9 | 1.057 | 0.186 |
| Iso-valeric | 0.69 | 0.76 | 0.91 | 0.064 | 0.401 |
| Valeric | 1.45 | 1.58 | 1.66 | 0.077 | 0.567 |
| Sum of SCFAs | 95.7 | 84.9 | 85.2 | 4.467 | 0.555 |
| SCFA profile (% of total SCFAs) | |||||
| Acetic | 74.5 c | 68.4 a | 71.4 b | 0.675 | 0.001 |
| Propionic | 4.46 | 4.36 | 4.98 | 0.258 | 0.585 |
| Butyric | 17.7 a | 23.4 b | 18.8 a | 4.767 | 0.006 |
| Birds at 42 days of age | |||||
| SCFA concentrations (μmol/g) | |||||
| Acetic | 54.9 | 55.1 | 47.2 | 2.351 | 0.300 |
| Propionic | 4.80 | 4.30 | 3.72 | 0.285 | 0.297 |
| Iso-butyric | 1.15 | 0.82 | 0.84 | 0.064 | 0.061 |
| Butyric | 12.9 | 12.6 | 11.9 | 0.849 | 0.888 |
| Iso-valeric | 1.05 | 0.70 | 0.72 | 0.084 | 0.166 |
| Valeric | 1.46 | 1.28 | 1.27 | 0.080 | 0.562 |
| Sum of SCFAs | 76.3 | 74.8 | 65.7 | 3.252 | 0.357 |
| SCFA profile (% of total SCFAs) | |||||
| Acetic | 72.0 | 73.8 | 72.2 | 0.593 | 0.426 |
| Propionic | 6.46 | 5.82 | 5.88 | 0.328 | 0.686 |
| Butyric | 16.6 | 16.5 | 17.2 | 0.603 | 0.878 |
| Indices | Dietary Treatments 1 | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | Anta®Phyt | PreAcid | |||
| Duodenum | |||||
| Villus height, µm | 1728.2 | 1646.1 | 1615.8 | 54.04 | 0.692 |
| Crypt depth, µm | 197.9 | 186.3 | 179.2 | 5.82 | 0.431 |
| Villus width, µm | 172.4 | 166.8 | 179.8 | 4.79 | 0.549 |
| Wall thickness, µm | 153.0 | 165.6 | 135.7 | 6.29 | 0.15 |
| Villus area, µm2 | 971.4 | 893.1 | 937.7 | 42.32 | 0.761 |
| Villus height to crypt depth | 9.10 | 9.03 | 9.11 | 0.36 | 0.996 |
| Jejunum | |||||
| Villus height, µm | 1047.8 | 1061.8 | 955.5 | 36.48 | 0.471 |
| Crypt depth, µm | 171.3 | 161.3 | 152.1 | 4.69 | 0.257 |
| Villus width, µm | 155.7 | 137.3 | 144.3 | 5.41 | 0.364 |
| Wall thickness, µm | 126.6 | 134.1 | 116.7 | 4.62 | 0.314 |
| Villus area, µm2 | 532.8 | 471 | 440.6 | 26.48 | 0.37 |
| Villus height to crypt depth | 6.25 | 6.58 | 6.40 | 0.227 | 0.84 |
| Ileum | |||||
| Villus height, µm | 703.7 | 728.9 | 656.9 | 18.7 | 0.279 |
| Crypt depth, µm | 168.3 | 170 | 152.4 | 3.8 | 0.101 |
| Villus width, µm | 178.4 | 166.1 | 174.0 | 5.0 | 0.607 |
| Wall thickness, µm | 138.6 | 143.9 | 130.6 | 4.3 | 0.453 |
| Villus area, µm2 | 425.1 | 401.1 | 381.5 | 16.2 | 0.564 |
| Villus height to crypt depth | 4.24 | 4.28 | 4.33 | 0.1 | 0.934 |
| Indices | Dietary Treatments 1 | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | Anta®Phyt | PreAcid | |||
| Duodenum | |||||
| Villus height, µm | 1889.4 | 1860.9 | 1841.4 | 32.76 | 0.836 |
| Crypt depth, µm | 198.8 a | 196.3 a | 217.4 b | 3.01 | 0.005 |
| Villus width, µm | 162.3 c | 136.5 a | 148.8 b | 2.63 | 0.001 |
| Wall thickness, µm | 179.9 | 182.1 | 190.6 | 3.49 | 0.434 |
| Villus area, µm2 | 982.2 b | 820.9 a | 877.8 a | 23.14 | 0.011 |
| Villus height to crypt depth | 9.50 | 9.52 | 8.49 | 0.21 | 0.051 |
| Jejunum | |||||
| Villus height, µm | 1221.2 | 1264.2 | 1274.4 | 31.11 | 0.774 |
| Crypt depth, µm | 166.4 a | 162.3 a | 178.3 b | 2.06 | 0.001 |
| Villus width, µm | 154.2 b | 130.2 a | 126.6 a | 2.68 | 0.001 |
| Wall thickness, µm | 149.4 a | 158.0 b | 162.8 b | 1.76 | 0.004 |
| Villus area, µm2 | 613.9 | 478.6 | 518.7 | 23.38 | 0.057 |
| Villus height to crypt depth | 7.32 | 7.07 | 7.13 | 0.271 | 0.931 |
| Ileum | |||||
| Villus height, µm | 882.8 | 851.6 | 890.9 | 23 | 0.778 |
| Crypt depth, µm | 171.9 | 164.8 | 172.8 | 2.12 | 0.246 |
| Villus width, µm | 149.3 b | 143.9 ab | 140.9 a | 1.44 | 0.048 |
| Wall thickness, µm | 169.9 | 164.2 | 164.6 | 3.84 | 0.804 |
| Villus area, µm2 | 430.6 | 403.5 | 406.0 | 10.92 | 0.539 |
| Villus height to crypt depth | 5.19 | 5.13 | 5.10 | 0.13 | 0.982 |
| Indices | Dietary Treatments 1 | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | Anta®Phyt | PreAcid | |||
| Duodenum | |||||
| Villus height, µm | 1822.8 | 1738.1 | 1879.7 | 72.47 | 0.731 |
| Crypt depth, µm | 219.8 | 209.9 | 210.6 | 4.44 | 0.617 |
| Villus width, µm | 173.0 b | 146.6 a | 161.5 ab | 3.57 | 0.007 |
| Wall thickness, µm | 192.5 | 209.7 | 189.4 | 7.98 | 0.542 |
| Villus area, µm2 | 1023.6 | 822.3 | 982.8 | 50.76 | 0.236 |
| Villus height to crypt depth | 8.41 | 8.35 | 9.15 | 0.323 | 0.545 |
| Jejunum | |||||
| Villus height, µm | 1296.5 | 1342.6 | 1340.5 | 33.11 | 0.823 |
| Crypt depth, µm | 173.0 a | 189.9 b | 195.0 b | 3.62 | 0.026 |
| Villus width, µm | 147.4 b | 122.6 a | 126.9 a | 3.56 | 0.007 |
| Wall thickness, µm | 180.3 | 161.7 | 161 | 6.88 | 0.442 |
| Villus area, µm2 | 636.3 | 528.5 | 546.8 | 19.44 | 0.053 |
| Villus height to crypt depth | 7.69 | 7.09 | 6.88 | 0.242 | 0.365 |
| Ileum | |||||
| Villus height, µm | 786.2 | 908.1 | 840.1 | 37.68 | 0.423 |
| Crypt depth, µm | 161.6 | 177.4 | 170.9 | 3.89 | 0.242 |
| Villus width, µm | 135.2 | 135.2 | 134.8 | 4.04 | 0.999 |
| Wall thickness, µm | 182.8 | 179.1 | 172.4 | 8.39 | 0.887 |
| Villus area, µm2 | 342.9 | 397.6 | 365.3 | 15.65 | 0.368 |
| Villus height to crypt depth | 4.93 | 5.14 | 4.70 | 0.203 | 0.690 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ząbek, K.; Szkopek, D.; Michalczuk, M.; Konieczka, P. Dietary Phytogenic Combination with Hops and a Mixture of a Free Butyrate Acidifier and Gluconic Acid Maintaining the Health Status of the Gut and Performance in Chickens. Animals 2020, 10, 1335. https://doi.org/10.3390/ani10081335
Ząbek K, Szkopek D, Michalczuk M, Konieczka P. Dietary Phytogenic Combination with Hops and a Mixture of a Free Butyrate Acidifier and Gluconic Acid Maintaining the Health Status of the Gut and Performance in Chickens. Animals. 2020; 10(8):1335. https://doi.org/10.3390/ani10081335
Chicago/Turabian StyleZąbek, Katarzyna, Dominika Szkopek, Monika Michalczuk, and Paweł Konieczka. 2020. "Dietary Phytogenic Combination with Hops and a Mixture of a Free Butyrate Acidifier and Gluconic Acid Maintaining the Health Status of the Gut and Performance in Chickens" Animals 10, no. 8: 1335. https://doi.org/10.3390/ani10081335
APA StyleZąbek, K., Szkopek, D., Michalczuk, M., & Konieczka, P. (2020). Dietary Phytogenic Combination with Hops and a Mixture of a Free Butyrate Acidifier and Gluconic Acid Maintaining the Health Status of the Gut and Performance in Chickens. Animals, 10(8), 1335. https://doi.org/10.3390/ani10081335