Form of Supplemental Selenium in Vitamin-Mineral Premixes Differentially Affects Early Luteal and Gestational Concentrations of Progesterone, and Postpartum Concentrations of Prolactin in Beef Cows

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Procedure
2.2. Experimental Regimen
2.2.1. Effect of Form of Se on Early Luteal Phase Concentrations of P4
2.2.2. Effect of Form of Se on Concentrations of P4 during Gestation
2.2.3. Effect of Form of Se on Concentrations of PRL during Lactation
2.3. Se and Hormone Analyses
2.4. Statistical Analysis
3. Results
3.1. Concentrations of Se in Whole Blood
3.2. Concentrations of P4 during the Early Luteal Phase
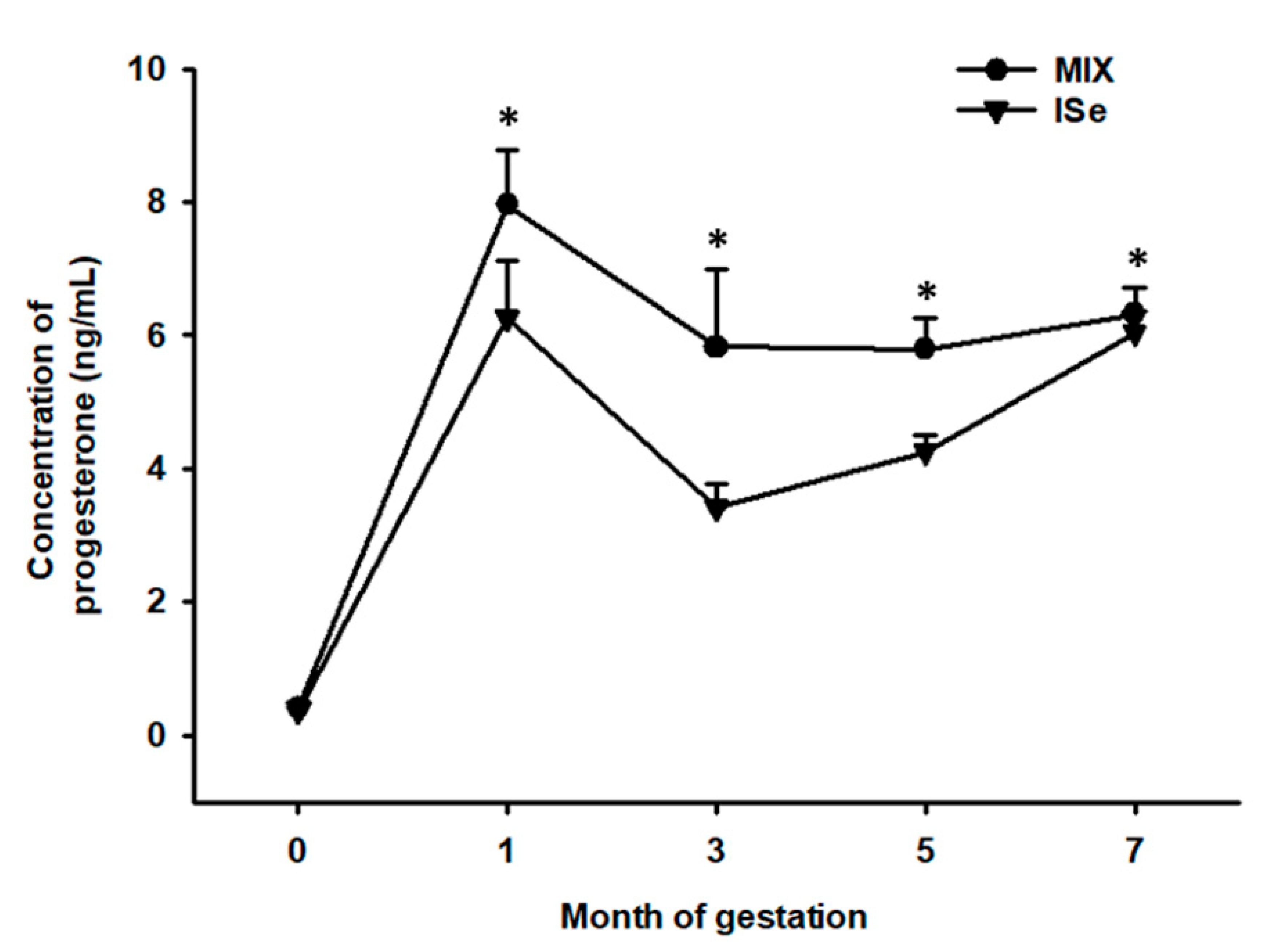
3.3. Concentrations of P4 during Gestation
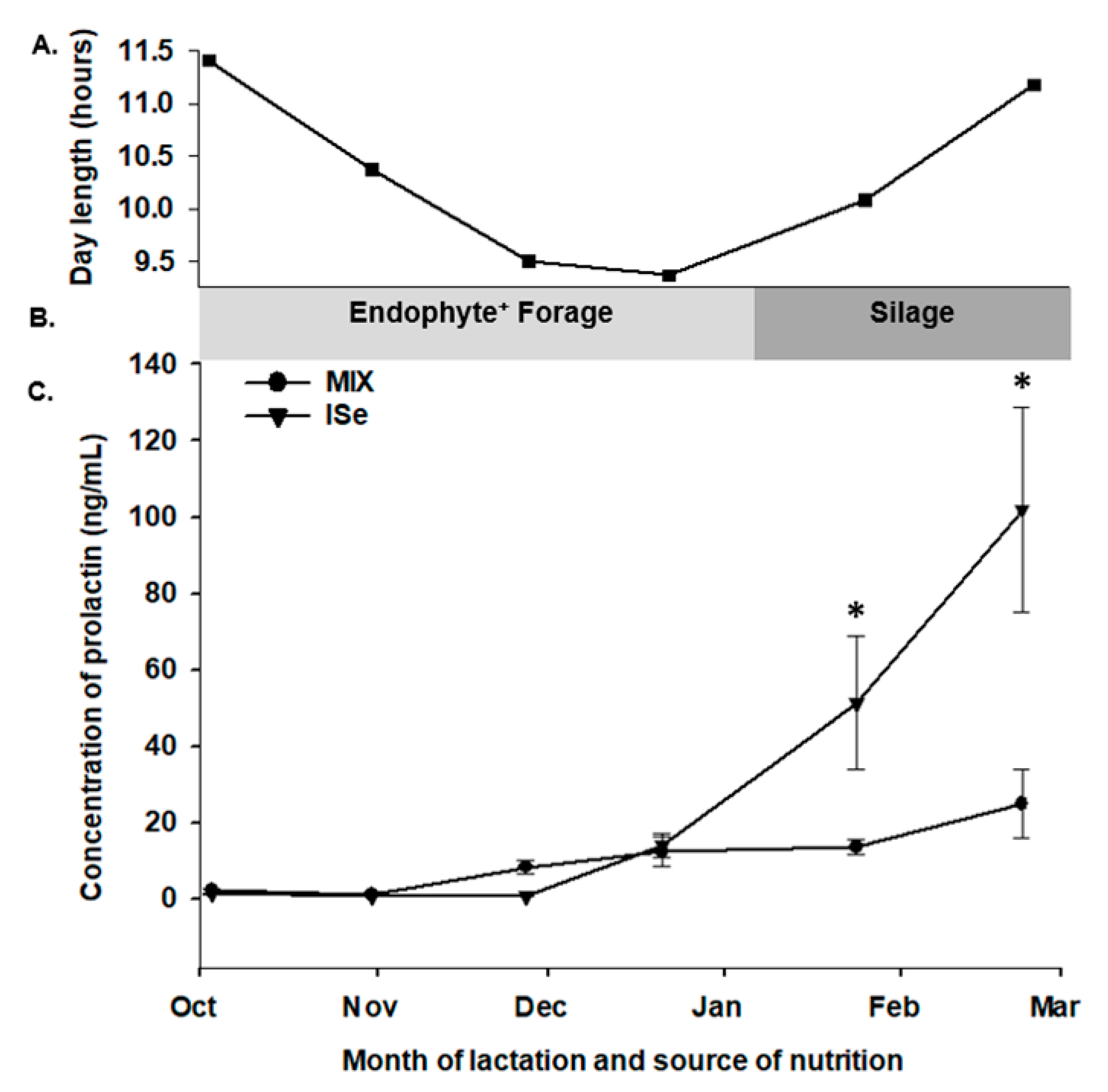
3.4. Concentrations of PRL during Lactation
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Paszkowski, T.; Traub, A.I.; Robinson, S.Y.; McMaster, D. Selenium dependent glutathione peroxidase activity in human follicular fluid. Clin. Chim. Acta 1995, 236, 173–180. [Google Scholar] [CrossRef]
- Combs, G., Jr.; Noguchi, T.; Scott, M.L. Mechanisms of action of selenium and vitamin E in protection of biological membranes. Fed. Proc. 1975, 34, 2090–2095. [Google Scholar]
- Steinbrenner, H.; Bilgic, E.; Alili, L.; Sies, H.; Brenneisen, P. Selenoprotein P protects endothelial cells from oxidative damage by stimulation of glutathione peroxidase expression and activity. Free Radic. Res. 2006, 40, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Ghadi, F.E.; Majhotra, A.; Ghara, A.R.; Dhawan, D.K. Selenium as a modulator of membrane stability parameters and surface changes during the initiation phase of 1,2-dimethylhydrazine induced colorectal carcinogenesis. Mol. Cell. Biochem. 2012, 369, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Gleed, P.T.; Allen, W.M.; Mallinson, C.B.; Rowlands, G.J.; Sansom, B.F.; Vagg, M.J.; Casell, R.D. Effects of selenium and copper supplementation on the growth of beef steers. Vet. Rec. 1983, 113, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Erskine, R.J.; Eberhart, R.J.; Grasso, P.J.; Scholz, R.W. Induction of Escherichia coli mastitis in cows fed selenium-deficient or selenium-supplemented diets. Am. J. Vet. Res. 1989, 50, 2093–2100. [Google Scholar]
- Boyne, R.; Arthur, J.R. Alterations of neutrophil function in selenium-deficient cattle. J. Comp. Pathol. 1979, 89, 151–158. [Google Scholar] [CrossRef]
- McClure, T.J.; Eamens, G.J.; Healy, P.J. Improved fertility in dairy cows after treatment with selenium pellets. Aust. Vet. J. 1986, 63, 144–146. [Google Scholar] [CrossRef]
- Enjalbert, F.; Lebreton, P.; Salat, O. Effects of copper, zinc and selenium status on performance and health in commercial dairy and beef herds: Retrospective study. J. Anim. Physiol. Anim. Nutr. (Berl.) 2006, 90, 459–466. [Google Scholar] [CrossRef]
- Kubota, J.; Allaway, W. Geographic distribution of trace element problems. In Micronutrients in Agriculture; Mortvedt, J.J., Lindsay, W., Giordano, P., Eds.; Soil Science Society of America: Madison, WI, USA, 1972; pp. 525–554. [Google Scholar]
- Pereira, A.S.C.; Santos, M.V.D.; Aferri, G.; da Silva Corte, R.R.P.; de Freitas Junior, J.E.; Leme, P.R.; Rennó, F.P. Lipid and selenium sources on fatty acid composition of intramuscular fat and muscle selenium concentration of Nellore steers. R. Bras. Zootec. 2012, 41, 2357–2363. [Google Scholar] [CrossRef]
- Liao, S.F.; Brown, K.R.; Stromberg, A.J.; Burris, W.R.; Boling, J.A.; Matthews, J.C. Dietary supplementation of selenium in inorganic and organic forms differentially and commonly alters blood and liver selenium concentrations and liver gene expression profiles of growing beef heifers. Biol. Trace Elem. Res. 2011, 140, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Brennen, K.M.; Burris, W.R.; Boling, J.A.; Matthews, J.C. Selenium content in blood fractions and liver of beef heifers is greater with a mix of inorganic/organic or organic versus inorganic supplemental selenium but the time required for maximal assimilation is tissue-specific. Biol. Trace Elem. Res. 2011, 144, 504–516. [Google Scholar] [CrossRef] [PubMed]
- Slavik, P.; Illek, J.; Brix, M.; Hlavicova, J.; Rajmon, R.; Jilek, F. Influence of organic versus inorganic dietary selenium supplementation on the concentration of selenium in colostrum, milk and blood of beef cows. Acta Vet. Scand. 2008, 50, 43. [Google Scholar] [CrossRef] [PubMed]
- Cerny, K.L.; Garbacik, S.; Skees, C.; Burris, W.R.; Matthews, J.C.; Bridges, P.J. Gestational form of selenium in free-choice mineral mixes affects transcriptome profiles of the neonatal calf testis, including those of steroidogenic and spermatogenic pathways. Biol. Trace Elem. Res. 2016, 169, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Cerny, K.L.; Anderson, L.H.; Burris, W.R.; Rhoads, M.; Matthews, J.C.; Bridges, P.J. Form of supplemental selenium fed to cycling cows affects systemic concentrations of progesterone but not those of estradiol. Theriogenology 2016, 85, 800–806. [Google Scholar] [CrossRef]
- Forde, N.; Carter, F.; Fair, T.; Crowe, M.A.; Evans, A.C.O.; Spencer, T.E.; Bazer, F.W.; McBride, R.; Boland, M.P.; O’Gaora, P.O.; et al. Progesterone-regulated changes in endometrial gene expression contribute to advanced conceptus development in cattle. Biol. Reprod. 2009, 81, 784–794. [Google Scholar] [CrossRef]
- Carter, F.; Forde, N.; Duffy, P.; Wade, M.; Fair, T.; Crowe, M.A.; Evans, A.C.; Kenny, D.A.; Roche, J.F.; Lonergan, P. Effect of increasing progesterone concentration from day 3 of pregnancy on subsequent embryo survival and development in beef heifers. Reprod. Fertil. Dev. 2008, 20, 368–375. [Google Scholar] [CrossRef]
- Garrett, J.E.; Geisert, R.D.; Zavy, M.T.; Morgan, G.L. Evidence for maternal regulation of early conceptus growth and development in beef cattle. J. Reprod. Fertil. 1988, 84, 437–446. [Google Scholar] [CrossRef]
- Wiltbank, M.C.; Souza, A.H.; Carvalho, P.D.; Bender, R.W.; Nascimento, A.B. Improving fertility to timed artificial insemination by manipulation of circulating progesterone concentrations in lactating dairy cattle. Reprod. Fertil. Dev. 2012, 24, 238–243. [Google Scholar] [CrossRef]
- Jia, Y.; Li, Q.; Burris, W.R.; Aiken, G.E.; Bridges, P.J.; Matthews, J.C. Forms of selenium in vitamin-mineral mixes differentially affect serum prolactin concentration and hepatic glutamine synthetase of steers grazing endophyte-infected tall fescue. J. Anim. Sci. 2018, 96, 715–727. [Google Scholar] [CrossRef]
- Stricker, P.; Grueter, R. Anction du lobe anterieur de l’hypophyse sur la montee laiteuse. C. R. Soc. Biol. 1928, 99, 1978–1980. [Google Scholar]
- Bole-Feysot, C.; Goffin, V.; Edery, M.; Binart, N.; Kelly, P.A. Prolactin and its receptor: Actions, signal transduction pathways, and phenotypes observed in prolactin receptor knockout mice. Endocr. Rev. 1998, 19, 225–268. [Google Scholar] [CrossRef] [PubMed]
- Goffin, V.; Binart, N.; Touraine, P.; Kelly, P. Prolactin: The new biology of an old hormone. Annu. Rev. Physiol. 2002, 64, 47–67. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.C.; Zheng, Z.; Patterson, J.D.; Bridges, P.J.; Stromberg, A.J.; Boling, J.A. Hepatic transcriptome profiles differ among maturing beef heifers supplemented with inorganic, organic, or mixed (50% inorganic: 50% organic) forms of dietary selenium. Biol. Trace Elem. Res. 2014, 160, 321–339. [Google Scholar] [CrossRef] [PubMed]
- Patterson, J.D.; Burris, W.R.; Boling, J.A.; Matthews, J.C. Individual intake of free-choice mineral mix by grazing beef cows may be less than typical formulation assumptions and form of selenium in mineral mix affects blood Se concentrations of cows and their suckling calves. Biol. Trace Elem. Res. 2013, 155, 38–48. [Google Scholar] [CrossRef]
- Monteiro, P.L.J., Jr.; Ribeiro, R.E.; Maciel, R.P.; Dias, A.L.G.; Sole, E., Jr.; Lima, F.S.; Bisinotto, R.S.; Thatcher, W.W.; Sartori, R.; Santos, J.E. Effects of supplemental progesterone after artificial insemination on expression of interferon-stimulated genes and fertility in dairy cows. J. Dairy. Sci. 2014, 97, 4907–4921. [Google Scholar] [CrossRef]
- Monteiro, P.L.J., Jr.; Nascimento, A.B.; Pontes, G.C.S.; Fernandes, G.O.; Melo, L.F.; Wiltbank, M.C.; Sartori, R. Progesterone supplementation after ovulation: Effects on corpus luteum function and on fertility of dairy cows subjected to AI or ET. Theriogenology 2015, 84, 1215–1224. [Google Scholar] [CrossRef]
- Carter, F.; Rings, F.; Mamo, S.; Holker, M.; Kuzmany, A.; Besenfedler, U.; Havlicek, V.; Mehta, J.P.; Tesfaye, D.; Schellander, K.; et al. Effect of elevated circulating progesterone concentration on bovine blastocyst development and global transcriptome following endoscopic transfer of in vitro produced embryos to the bovine oviduct. Biol. Reprod. 2010, 83, 707–719. [Google Scholar] [CrossRef]
- Stronge, A.J.; Sreenan, J.M.; Diskin, M.G.; Mee, J.F.; Kenny, D.A.; Morris, D.G. Post-insemination milk progesterone concentration and embryo survival in dairy cows. Theriogenology 2005, 64, 1212–1224. [Google Scholar] [CrossRef]
- Hatler, T.B.; Hayes, S.H.; Ray, D.L.; Reames, P.S.; Silvia, W.J. Effect of subluteal concentrations of progesterone on luteinizing hormone and ovulation in lactating dairy cows. Vet. J. 2008, 177, 360–368. [Google Scholar] [CrossRef]
- Schuenemann, G.M.; Edwards, J.L.; Hopkins, F.M.; Rohrbach, N.R.; Adair, H.S.; Scenna, F.N.; Waller, J.C.; Oliver, J.W.; Saxton, A.M.; Schrick, F.N. Fertility aspects in yearling beef bulls grazing endophyte-infected tall fescue pastures. Reprod. Fertil. Dev. 2005, 17, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Wahlen, R.; Evans, L.; Turner, J.; Hearn, R. The use of collision/reaction cell ICP-MS for the determination of elements in blood and serum samples. Spectroscopy 2005, 20, 84–89. [Google Scholar]
- Bernard, J.K.; Chestnut, A.B.; Erickson, B.H.; Kelly, F.M. Effects of prepartum consumption of endophyte-infected tall fescue on serum prolactin and subsequent milk production of holstein cows. J. Dairy Sci. 1993, 76, 1928–1933. [Google Scholar] [CrossRef]
- Gerloff, B.J. Effect of selenium supplementation on dairy cattle. J. Anim. Sci. 1992, 70, 3934–3940. [Google Scholar] [CrossRef]
- Dargatz, D.A.; Ross, P.F. Blood selenium concentrations in cows and heifers on 253 cow-calf operations in 18 states. J. Anim. Sci. 1996, 74, 2891–2895. [Google Scholar] [CrossRef]
- Beltman, M.E.; Lonergan, P.; Diskin, M.G.; Roche, J.F.; Crowe, M.A. Effect of progesterone supplementation in the first week post conception on embryo survival in beef heifers. Theriogenology 2009, 71, 1173–1179. [Google Scholar] [CrossRef]
- Mann, G.E.; Green, M.P.; Sinclair, K.D.; Demmers, K.J.; Fray, M.D.; Gutierrez, C.G.; Garnsworthy, P.C.; Webb, R. Effects of circulating progesterone and insulin on early embryo development in beef heifers. Anim. Reprod. Sci. 2003, 79, 71–79. [Google Scholar] [CrossRef]
- Forde, N.; Beltman, M.E.; Duffy, G.B.; Duffy, P.; Mehta, J.P.; O’Gaora, P.; Roche, J.F.; Lonergan, P.; Crowe, M.A. Changes in the endometrial transcriptome during the bovine estrous cycle: Effect of low circulating progesterone and consequences for conceptus elongation. Biol. Reprod. 2011, 84, 266–278. [Google Scholar] [CrossRef]
- Yan, L.; Robinson, R.; Shi, Z.; Mann, G. Efficacy of progesterone supplementation during early pregnancy in cows: A meta-analysis. Theriogenology 2016, 85, 1390–1398. [Google Scholar] [CrossRef]
- Prenant, A. La valeur morphologique du corps jaune. Son action physiologique et therapeutique possible. Rev. Gen. Sci. Pures Appl. 1898, 9, 646–650. (In French) [Google Scholar]
- Magnus, V. Ovariets betydning for svangerskabet med saerligt hensyn til corpus luteum. Nor. Mag. Laegevidensk 1901, 1138–1145. [Google Scholar]
- Fraenkel, L. Die function des corpus luteum. Arch. Gynecol. 1903, 68, 438–545. (In German) [Google Scholar] [CrossRef]
- Fraenkel, L. Experimentelle untersuchungen uber den einfluss des corpus luteum auf de insertion des eies. Anat. Anz. 1901, 294–300. [Google Scholar]
- Estergreen, V.L., Jr.; Frost, O.L.; Gomes, W.R.; Erb, R.E.; Bullard, J.F. The effect of ovariectomy on pregnancy maintenance and parturition in dairy cows. J. Dairy Sci. 1967, 50, 1293–1295. [Google Scholar] [CrossRef]
- McDonald, L.E.; Nichols, R.E.; McNutt, S.H. Studies on corpus luteum ablation and progesterone replacement therapy during pregnancy in the cow. Am. J. Vet. Res. 1952, 13, 446–451. [Google Scholar]
- McDonald, L.E.; McNutt, S.H.; Nichols, R.E. Retained placenta; experimental production and prevention. Am. J. Vet. Res. 1954, 15, 22–24. [Google Scholar]
- Chew, B.P.; Keller, H.F.; Erb, R.E.; Malven, P.V. Periparturient concentrations of prolactin, progesterone and the estrogens in blood plasma of cows retaining and not retaining fetal membranes. J. Anim. Sci. 1977, 44, 1055–1060. [Google Scholar] [CrossRef]
- Erb, R.E.; Gomes, W.R.; Randel, R.D.; Estergreen, V.L., Jr.; Frost, O.L. Effect of ovariectomy on concentration of progesterone in blood plasma and urinary estrogen excretion rate in the pregnant bovine. J. Dairy Sci. 1968, 51, 420–427. [Google Scholar] [CrossRef]
- Peters, R.R.; Tucker, H.A. Prolactin and growth hormone responses to photoperiod in heifers. Endocrinology 1978, 103, 229–234. [Google Scholar] [CrossRef]
- Tucker, H.A.; Petitclerc, D.; Zinn, S.A. The Influence of photoperiod on body weight gain, body composition, nutrient intake and hormone secretion. J. Anim. Sci. 1984, 59, 1610–1620. [Google Scholar] [CrossRef]
- Davenport, G.M.; Boling, J.A.; Rahe, C.H. Growth and endocrine responses of cattle to implantation of estradiol-17 beta during continuous or discontinuous grazing of high- and low-endophyte-infected tall fescue. J. Anim. Sci. 1993, 71, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Goetsch, A.L.; Jones, A.L.; Stokes, S.R.; Beers, K.W.; Piper, E.L. Intake, digestion, passage rate and serum prolactin in growing dairy steers fed endophyte-infected fescue with noninfected fescue, clover or wheat straw. J. Anim. Sci. 1987, 64, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Jia, Y.; Burris, W.R.; Bridges, P.J.; Matthews, J.C. Forms of selenium in vitamin-mineral mixes differentially affect the expression of genes responsible for prolactin, ACTH, and alpha-MSH synthesis and mitochondrial dysfunction in pituitaries of steers grazing endophyte-infected tall fescue. J. Anim. Sci. 2018, 97, 631–643. [Google Scholar] [CrossRef] [PubMed]
- O’Neal, K.D.; Montgomery, D.W.; Truong, T.M.; Yu-Lee, L.Y. Prolactin gene expression in human thymocytes. Mol. Cell. Endocrinol. 1992, 87, 19–23. [Google Scholar] [CrossRef]
- Pellegrini, I.; Lebrun, J.J.; Ali, S.; Kelly, P.A. Expression of prolactin and its receptor in human lymphoid cells. Mol. Endocrinol. 1992, 6, 1023–1031. [Google Scholar] [CrossRef]
- Matera, L.; Cutufia, M.; Geuna, M.; Contarini, M.; Buttiglieri, S.; Galin Fazzari, A.; Cavaliere, C. Prolactin is an autocrine growth factor for the Jurkat human T-leukemic cell line. J. Neuroimmunol. 1997, 79, 12–21. [Google Scholar] [CrossRef]
- Gingras, M.C.; Margolin, J.F. Differential expression of multiple unexpected genes during U937 cell and macrophage differentiation detected by suppressive subtractive hybridization. Exp. Hematol. 2000, 28, 65–76. [Google Scholar] [CrossRef]
- Orbach, H.; Zandman-Goddard, G.; Amital, H.; Barak, V.; Szekanecz, Z.; Szucs, G.; Danko, K.; Nagy, K.; Csepany, T.; Carvalho, J.F.; et al. Novel biomarkers in autoimmune diseases: Prolactin, ferritin, vitamin D, and TPA levels in autoimmune diseases. Ann. N. Y. Acad. Sci. 2007, 1109, 385–400. [Google Scholar] [CrossRef]
- Freeman, M.E.; Kanyicska, B.; Lerant, A.; Nagy, G. Prolactin: Structure, function, and regulation of secretion. Physiol. Rev. 2000, 80, 1523–1631. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variable | Treatment | p-Value 2 | |
|---|---|---|---|
| ISe | MIX | ||
| Mean ± SEM | Mean ± SEM | ||
| Progesterone (ng/mL) | |||
| Cerny et al., 2016 * | |||
| No. of cows (n) | 9 | 9 | |
| Day 6 † | 3.44 ± 0.18 a | 5.14 ± 0.60 b | 0.035 |
| Experimental Study | |||
| No. of cows (n) | 12 | 12 | |
| Day 4 | 1.02 ± 0.22 | 0.94 ± 0.12 | 0.740 |
| Day 7 † | 2.92 ± 0.27 a | 3.91 ± 0.16 b | 0.006 |
| Day 10 | 7.17 ± 0.54 | 6.36 ± 0.55 | 0.308 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carr, S.; Jia, Y.; Crites, B.; Hamilton, C.; Burris, W.; Edwards, J.L.; Matthews, J.; Bridges, P.J. Form of Supplemental Selenium in Vitamin-Mineral Premixes Differentially Affects Early Luteal and Gestational Concentrations of Progesterone, and Postpartum Concentrations of Prolactin in Beef Cows. Animals 2020, 10, 967. https://doi.org/10.3390/ani10060967
Carr S, Jia Y, Crites B, Hamilton C, Burris W, Edwards JL, Matthews J, Bridges PJ. Form of Supplemental Selenium in Vitamin-Mineral Premixes Differentially Affects Early Luteal and Gestational Concentrations of Progesterone, and Postpartum Concentrations of Prolactin in Beef Cows. Animals. 2020; 10(6):967. https://doi.org/10.3390/ani10060967
Chicago/Turabian StyleCarr, Sarah, Yang Jia, Benjamin Crites, Charles Hamilton, Walter Burris, J. Lannett Edwards, James Matthews, and Phillip J. Bridges. 2020. "Form of Supplemental Selenium in Vitamin-Mineral Premixes Differentially Affects Early Luteal and Gestational Concentrations of Progesterone, and Postpartum Concentrations of Prolactin in Beef Cows" Animals 10, no. 6: 967. https://doi.org/10.3390/ani10060967
APA StyleCarr, S., Jia, Y., Crites, B., Hamilton, C., Burris, W., Edwards, J. L., Matthews, J., & Bridges, P. J. (2020). Form of Supplemental Selenium in Vitamin-Mineral Premixes Differentially Affects Early Luteal and Gestational Concentrations of Progesterone, and Postpartum Concentrations of Prolactin in Beef Cows. Animals, 10(6), 967. https://doi.org/10.3390/ani10060967