Effect of Feed Additives as Alternatives to In-feed Antimicrobials on Production Performance and Intestinal Clostridium perfringens Counts in Broiler Chickens


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Experimental Design
2.3. Clostridium Perfringens Quantification
2.4. Post Mortem Examination
2.5. Production Performance Measurements
2.6. Statistical Analysis
3. Results
3.1. Impact of the Collective ATA Group on Necrotic Enteritis, Intestinal CP Counts and Production Performance
3.2. Impact of ATA Classes on Intestinal CP Counts and Production Performance
3.3. Impact of Treatments on Intestinal CP Counts and Production Performance
3.4. Active Components with Combined Beneficial Effects on FCR14–28 and CP Counts
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grave, K.; Jensen, V.F.; Odensvik, K.; Wierup, M.; Bangen, M. Usage of veterinary therapeutic antimicrobials in Denmark, Norway and Sweden following termination of antimicrobial growth promoter use. Prev. Vet. Med. 2006, 75, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Norwegian Scientific Committee for Food Safety. The risk of development of antimicrobial resistance with the use of coccidiostats in poultry diets. In Opinion of the Panel on Animal Feed of the Norwegian Scientific Committee for Food Safety; Norwegian Scientific Committee for Food Safety: Oslo, Norway, 2015. [Google Scholar]
- Regulation (EC) No 1831/2003 of the European Parliament and of the Council of the European Union on additives for use in animal nutrition. Off. J. Eur. Union 2003, OJ L268, 29–43.
- Castanon, J.I.R. History of the use of antibiotic as growth promoters in European poultry feeds. Poult. Sci. 2007, 86, 2466–2471. [Google Scholar] [CrossRef] [PubMed]
- Kevin, D.A.; Meujo, D.A.F.; Hamann, M.T. Polyether ionophores: Broad-spectrum and promising biologically active molecules for the control of drug-resistant bacteria and parasites. Expert Opin. Drug Dis. 2009, 4, 109–146. [Google Scholar] [CrossRef] [PubMed]
- Elwinger, K.; Schneitz, C.; Berndtson, E.; Fossum, O.; Teglof, B.; Engstom, B. Factors Affecting the Incidence of Necrotic Enteritis, Cecal Carriage of Clostridium-Perfringens and Bird Performance in Broiler Chicks. Acta Vet. Scand. 1992, 33, 369–378. [Google Scholar] [PubMed]
- Martel, A.; Devriese, L.A.; Cauwerts, K.; De Gussem, K.; Decostere, A.; Haesebrouck, F. Susceptibility of Clostridium perfringens strains from broiler chickens to antibiotics and anticoccidials. Avian Pathol. 2004, 33, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Hickey, E.E.; Wong, H.S.; Khazandi, M.; Ogunniyi, A.D.; Petrovski, K.R.; Garg, S.; Page, S.W.; O’Handley, R.; Trott, D.J. Repurposing Ionophores as novel antimicrobial agents for the treatment of bovine mastitis caused by Gram-positive pathogens. J. Vet. Pharmacol. Ther. 2018, 41, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, O.; Greko, C.; Bengtsson, B.; Englund, S. Genetic diversity among VRE isolates from Swedish broilers with the coincidental finding of transferrable decreased susceptibility to narasin. J. Appl. Microbiol. 2012, 112, 716–722. [Google Scholar] [CrossRef]
- NORM/NORM-VET 2017. Usage of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Norway; The Norwegian Surveillance System for Antimicrobial Drug Resistance (NORM): Tromsø/Oslo, Norway, 2018; ISSN 1502-2307. [Google Scholar]
- Broom, L.J. The sub-inhibitory theory for antibiotic growth promoters. Poult. Sci. 2017, 96, 3104–3108. [Google Scholar] [CrossRef]
- Stutz, M.W.; Johnson, S.L.; Judith, F.R.; Miller, B.M. In vitro and in vivo evaluations of the antibiotic efrotomycin. Poult. Sci. 1983, 62, 1612–1618. [Google Scholar] [CrossRef]
- Stutz, M.W.; Lawton, G.C. Effects of diet and antimicrobials on growth, feed efficiency, intestinal Clostridium perfringens, and ileal weight of broiler chicks. Poult. Sci. 1984, 63, 2036–2042. [Google Scholar] [CrossRef] [PubMed]
- Devriese, L.A.; Daube, G.; Hommez, J.; Haesebrouck, F. In-Vitro Susceptibility of Clostridium-Perfringens Isolated from Farm-Animals to Growth-Enhancing Antibiotics. J. Appl. Bacteriol. 1993, 75, 55–57. [Google Scholar] [CrossRef] [PubMed]
- Watkins, K.L.; Shryock, T.R.; Dearth, R.N.; Saif, Y.M. In-vitro antimicrobial susceptibility of Clostridium perfringens from commercial turkey and broiler chicken origin. Vet. Microbiol. 1997, 54, 195–200. [Google Scholar] [CrossRef]
- Silva, R.O.S.; Salvarani, F.M.; Assis, R.A.; Martins, N.R.S.; Pires, P.S.; Lobato, F.C.F. Antimicrobial Susceptibility of Clostridium Perfringens Strains Isolated from Broiler Chickens. Braz. J. Microbiol. 2009, 40, 262–264. [Google Scholar] [CrossRef]
- Briggs, C.A.; Coates, M.E.; Lev, M. Bacteriological studies of infected and uninfected chicks in relation to antibiotic growth stimulation. Nature 1956, 178, 1125–1126. [Google Scholar] [CrossRef]
- Lev, M.; Forbes, M. Growth response to dietary penicillin of germ-free chicks and of chicks with a defined intestinal flora. Br. J. Nutr. 1959, 13, 78–84. [Google Scholar] [CrossRef]
- Kaldhusdal, M.; Hofshagen, M. Barley Inclusion and Avoparcin Supplementation in Broiler Diets. 2. Clinical, Pathological, and Bacteriological Findings in a Mild Form of Necrotic Enteritis. Poult. Sci. 1992, 71, 1145–1153. [Google Scholar] [CrossRef]
- Lovland, A.; Kaldhusdal, M. Severely impaired production performance in broiler flocks with high incidence of Clostridium perfringens-associated hepatitis. Avian Pathol. 2001, 30, 73–81. [Google Scholar] [CrossRef]
- Timbermont, L.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Necrotic enteritis in broilers: An updated review on the pathogenesis. Avian Pathol. 2011, 40, 341–347. [Google Scholar] [CrossRef]
- Hermans, P.G.; Morgan, K.L. Prevalence and associated risk factors of necrotic enteritis on broiler farms in the United Kingdom; a cross-sectional survey. Avian Pathol. 2007, 36, 43–51. [Google Scholar] [CrossRef]
- Prescott, J.F.; Smyth, J.A.; Shojadoost, B.; Vince, A. Experimental reproduction of necrotic enteritis in chickens: A review. Avian Pathol. 2016, 45, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Kabir, S.M.L. The Role of Probiotics in the Poultry Industry. Int. J. Mol. Sci. 2009, 10, 3531–3546. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.A.; Duc, L.H.; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Pourabedin, M.; Zhao, X. Prebiotics and gut microbiota in chickens. FEMS Microbiol. Lett. 2015, 362, fnv122. [Google Scholar] [CrossRef]
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef]
- Suresh, G.; Das, R.K.; Brar, S.K.; Rouissi, T.; Ramirez, A.A.; Chorfi, Y.; Godbout, S. Alternatives to antibiotics in poultry feed: Molecular perspectives. Crit. Rev. Microbiol. 2018, 44, 318–335. [Google Scholar] [CrossRef]
- Yang, C.B.; Chowdhury, M.A.K.; Hou, Y.Q.; Gong, J. Phytogenic Compounds as Alternatives to In-Feed Antibiotics: Potentials and Challenges in Application. Pathogens 2015, 4, 137–156. [Google Scholar] [CrossRef]
- Huyghebaert, G.; Ducatelle, R.; Van Immerseel, F. An update on alternatives to antimicrobial growth promoters for broilers. Vet. J. 2011, 187, 182–188. [Google Scholar] [CrossRef]
- Dittoe, D.K.; Ricke, S.C.; Kiess, A.S. Organic Acids and Potential for Modifying the Avian Gastrointestinal Tract and Reducing Pathogens and Disease. Front. Vet. Sci. 2018, 5. [Google Scholar] [CrossRef]
- Blajman, J.E.; Frizzo, L.S.; Zbrun, M.V.; Astesana, D.M.; Fusari, M.L.; Soto, L.P.; Rosmini, M.R.; Signorini, M.L. Probiotics and broiler growth performance: A meta-analysis of randomised controlled trials. Brit. Poult. Sci. 2014, 55, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Lovland, A.; Kaldhusdal, M.; Redhead, K.; Skjerve, E.; Lillehaug, A. Maternal vaccination against subclinical necrotic enteritis in broilers. Avian Pathol. 2004, 33, 83–92. [Google Scholar] [CrossRef]
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Kawazoe, U.; Bordin, E.L.; de Lima, C.A.; Dias, L.A.V. Characterisation and histopathological observations of a selected Brazilian precocious line of Eimeria acervulina. Vet. Parasitol. 2005, 131, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.O.; Zhao, J.; Yang, M.S.; Kim, W.I.; Cho, H.S.; Lim, C.W.; Kim, B. Oocyst-Shedding Patterns of Three Eimeria Species in Chickens and Shedding Pattern Variation Depending on the Storage Period of Eimeria Tenella Oocysts. J. Parasitol. 2018, 104, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Gholamiandehkordi, A.R.; Timbermont, L.; Lanckriet, A.; Van den Broeck, W.; Pedersen, K.; Dewulf, J.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Quantification of gut lesions in a subclinical necrotic enteritis model. Avian Pathol. 2007, 36, 375–382. [Google Scholar] [CrossRef]
- Novoa-Garrido, M.; Larsen, S.; Kaldhusdal, M. Association between gizzard lesions and increased caecal Clostridium perfringens counts in broiler chickens. Avian Pathol. 2006, 35, 367–372. [Google Scholar] [CrossRef]
- Julian, R.J. Production and growth related disorders and other metabolic diseases of poultry—A review. Vet. J. 2005, 169, 350–369. [Google Scholar] [CrossRef]
- Dierick, E.; Hirvonen, O.P.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F.; Goossens, E. Rapid growth predisposes broilers to necrotic enteritis. Avian Pathol. 2019, 48, 416–422. [Google Scholar] [CrossRef]
- Teo, A.Y.L.; Tan, H.M. Inhibition of Clostridium perfringens by a novel strain of Bacilius subtilis isolated from the gastrointestinal tracts of healthy chickens. Appl. Environ. Microbiol. 2005, 71, 4185–4190. [Google Scholar] [CrossRef]
- Jayaraman, S.; Das, P.P.; Saini, P.C.; Roy, B.; Chatterjee, P.N. Use of Bacillus Subtilis PB6 as a potential antibiotic growth promoter replacement in improving performance of broiler birds. Poult. Sci. 2017, 96, 2614–2622. [Google Scholar] [CrossRef]
- Abudabos, A.M.; Yehia, H.M. Effect of dietary mannan oligosaccharide from Saccharomyces cerevisiae on live performance of broilers under Clostridium perfringens challenge. Ital. J. Anim. Sci. 2013, 12, e38. [Google Scholar] [CrossRef]
- Alizadeh, M.; Rogiewicz, A.; McMillan, E.; Rodriguez-Lecompte, J.C.; Patterson, R.; Slominski, B.A. Effect of yeast-derived products and distillers dried grains with solubles (DDGS) on growth performance and local innate immune response of broiler chickens challenged with Clostridium perfringens. Avian Pathol. 2016, 45, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Skrivanova, E.; Marounek, M.; Dlouha, G.; Kanka, J. Susceptibility of Clostridium perfringens to C-2-C-18 fatty acids. Lett. Appl. Microbiol. 2005, 41, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhang, H.J.; Yu, S.H.; Wu, S.G.; Yoon, I.; Quigley, J.; Gao, Y.P.; Qi, G.H. Effects of yeast culture in broiler diets on performance and immunomodulatory functions. Poult. Sci. 2008, 87, 1377–1384. [Google Scholar] [CrossRef]
- Lee, H.S. Antimicrobial properties of turmeric (Curcuma longa L.) rhizome-derived ar-turmerone and curcumin. Food Sci. Biotechnol. 2006, 15, 559–563. [Google Scholar]
- Lee, S.H.; Lillehoj, H.S.; Jang, S.I.; Lillehoj, E.P.; Min, W.; Bravo, D.M. Dietary supplementation of young broiler chickens with Capsicum and turmeric oleoresins increases resistance to necrotic enteritis. Brit. J. Nutr. 2013, 110, 840–847. [Google Scholar] [CrossRef]
- Kim, J.E.; Lillehoj, H.S.; Hong, Y.H.; Kim, G.B.; Lee, S.H.; Lillehoj, E.P.; Bravo, D.M. Dietary Capsicum and Curcuma longa oleoresins increase intestinal microbiome and necrotic enteritis in three commercial broiler breeds. Res. Vet. Sci. 2015, 102, 150–158. [Google Scholar] [CrossRef]
- Abou-Elkhair, R.; Ahmed, H.A.; Selim, S. Effects of Black Pepper (Piper Nigrum), Turmeric Powder (Curcuma Longa) and Coriander Seeds (Coriandrum Sativum) and Their Combinations as Feed Additives on Growth Performance, Carcass Traits, Some Blood Parameters and Humoral Immune Response of Broiler Chickens. Asian Austral. J. Anim. 2014, 27, 847–854. [Google Scholar] [CrossRef]
- Roy, K.K.; Lyhs, U.; Pedersen, K.T. Inhibition studies of natural resin acids to Clostridium perfringens and Escherichia coli O149. In Proceedings of the 1st International Conference on Necrotic Enteritis in Poultry, Copenhagen, Denmark, 10–12 June 2015. [Google Scholar]
- Dowling, L. Ionophore Toxicity in Chickens - a Review of Pathology and Diagnosis. Avian Pathol. 1992, 21, 355–368. [Google Scholar] [CrossRef]
- La Ragione, R.M.; Woodward, M.J. Competitive exclusion by Bacillus subtilis spores of Salmonella enterica serotype Enteritidis and Clostridium perfringens in young chickens. Vet. Microbiol. 2003, 94, 245–256. [Google Scholar] [CrossRef]
- Jayaraman, S.; Thangavel, G.; Kurian, H.; Mani, R.; Mukkalil, R.; Chirakkal, H. Bacillus subtilis PB6 improves intestinal health of broiler chickens challenged with Clostridium perfringens-induced necrotic enteritis. Poult. Sci. 2013, 92, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.H.; Zhang, N.; Han, J.C.; Chang, C.W.; Hsiao, F.S.H.; Yu, Y.H. Optimization of surfactin production from Bacillus subtilis in fermentation and its effects on Clostridium perfringens-induced necrotic enteritis and growth performance in broilers. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1232–1244. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Lillehoj, H.S.; Siragusa, G.R. Direct-Fed Microbials and Their Impact on the Intestinal Microflora and Immune System of Chickens. J. Poult. Sci. 2010, 47, 106–114. [Google Scholar] [CrossRef]
- Tellez, G.; Pixley, C.; Wolfenden, R.E.; Layton, S.L.; Hargis, B.M. Probiotics/direct fed microbials for Salmonella control in poultry. Food Res. Int. 2012, 45, 628–633. [Google Scholar] [CrossRef]
- Ma, Y.B.; Wang, W.W.; Zhang, H.J.; Wang, J.; Zhang, W.M.; Gao, J.; Wu, S.G.; Qi, G.H. Supplemental Bacillus subtilis DSM 32315 manipulates intestinal structure and microbial composition in broiler chickens. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Mingmongkolchai, S.; Panbangred, W. Bacillus probiotics: An alternative to antibiotics for livestock production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef]
- Zhang, B.; Guo, Y.M.; Wang, Z. The modulating effect of beta-1,3/1,6-glucan supplementation in the diet on performance and immunological responses of broiler chickens. Asian Austral. J. Anim. 2008, 21, 237–244. [Google Scholar] [CrossRef]
- Tian, X.Y.; Shao, Y.J.; Wang, Z.; Guo, Y.M. Effects of dietary yeast beta-glucans supplementation on growth performance, gut morphology, intestinal Clostridium perfringens population and immune response of broiler chickens challenged with necrotic enteritis. Anim. Feed Sci. Technol. 2016, 215, 144–155. [Google Scholar] [CrossRef]
- Chae, B.J.; Lohakare, J.D.; Moon, W.K.; Lee, S.L.; Park, Y.H.; Hahn, T.W. Effects of supplementation of beta-glucan on the growth performance and immunity in broilers. Res. Vet. Sci. 2006, 80, 291–298. [Google Scholar] [CrossRef]
- Morales-Lopez, R.; Auclair, E.; Garcia, F.; Esteve-Garcia, E.; Brufau, J. Use of yeast cell walls; beta-1, 3/1, 6-glucans; and mannoproteins in broiler chicken diets. Poult. Sci. 2009, 88, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Si, W.; Ni, X.; Gong, J.; Yu, H.; Tsao, R.; Han, Y.; Chambers, J.R. Antimicrobial activity of essential oils and structurally related synthetic food additives towards Clostridium perfringens. J. Appl. Microbiol. 2009, 106, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Timbermont, L.; Lanckriet, A.; Dewulf, J.; Nollet, N.; Schwarzer, K.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Control of Clostridium perfringens-induced necrotic enteritis in broilers by target-released butyric acid, fatty acids and essential oils. Avian Pathol. 2010, 39, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Alanazi, S.; Alnoman, M.; Banawas, S.; Saito, R.; Sarker, M.R. The inhibitory effects of essential oil constituents against germination, outgrowth and vegetative growth of spores of Clostridium perfringens type A in laboratory medium and chicken meat. Food Microbiol. 2018, 73, 311–318. [Google Scholar] [CrossRef]
- Mitsch, P.; Zitterl-Eglseer, K.; Kohler, B.; Gabler, C.; Losa, R.; Zimpernik, I. The effect of two different blends of essential oil components on the proliferation of Clostridium perfringens in the intestines of broiler chickens. Poult. Sci. 2004, 83, 669–675. [Google Scholar] [CrossRef]
- Abildgaard, L.; Hojberg, O.; Schramm, A.; Balle, K.M.; Engberg, R.M. The effect of feeding a commercial essential oil product on Clostridium perfringens numbers in the intestine of broiler chickens measured by real-time PCR targeting the alpha-toxin-encoding gene (plc). Anim. Feed Sci. Technol. 2010, 157, 181–189. [Google Scholar] [CrossRef]
- Du, E.C.; Wang, W.W.; Gan, L.P.; Li, Z.; Guo, S.S.; Guo, Y.M. Effects of thymol and carvacrol supplementation on intestinal integrity and immune responses of broiler chickens challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2016, 7. [Google Scholar] [CrossRef]
- Lee, K.W.; Everts, H.; Kappert, H.J.; Yeom, K.H.; Beynen, A.C. Dietary carvacrol lowers body weight gain but improves feed conversion in female broiler chickens. J. Appl. Poult. Res. 2003, 12, 394–399. [Google Scholar] [CrossRef]
- Brenes, A.; Roura, E. Essential oils in poultry nutrition: Main effects and modes of action. Anim. Feed Sci. Technol. 2010, 158, 1–14. [Google Scholar] [CrossRef]
- Hippenstiel, F.; Abdel-Wareth, A.A.A.; Kehraus, S.; Sudekum, K.H. Effects of selected herbs and essential oils, and their active components on feed intake and performance of broilers—A review. Arch. Geflugelkd 2011, 75, 226–234. [Google Scholar]
- Lanckriet, A.; Timbermont, L.; De Gussem, M.; Marien, M.; Vancraeynest, D.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. The effect of commonly used anticoccidials and antibiotics in a subclinical necrotic enteritis model. Avian Pathol. 2010, 39, 63–68. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Chemical Composition | Starter Diet 2 | Grower Diet 3 |
|---|---|---|
| (g/kg feed) | 0–14 days | 14–28 days |
| Dry matter | 887.2 | 881.3 |
| Crude protein | 239.6 | 222.0 |
| Crude fat | 67.8 | 99.6 |
| Crude fiber | 30.3 | 29.0 |
| Nitrogen-free extracts | 493.7 | 479.0 |
| Ash | 55.8 | 51.7 |
| Lysine | 14.0 | 12.9 |
| Methionine + Cysteine | 11.6 | 11.1 |
| Threonine | 9.4 | 9.0 |
| Tryptophan | 2.7 | 2.5 |
| Arginine | 13.8 | 12.7 |
| Calcium (Ca) | 9.2 | 7.4 |
| Phosphorus (P) | 6.3 | 5.9 |
| Sodium (Na) | 1.4 | 1.6 |
| Potassium (K) | 7.7 | 7.4 |
| Chloride (Cl) | 2.3 | 2.2 |
| Magnesium (Mg) | 1.6 | 1.6 |
| NSP enzymes 4 and phytase | 0.15 | 0.15 |
| Metabolizable energy (MJ/kg) | 12.13 | 12.78 |
| ID 1 | Class 2 | Active Components and Product Description 3 | Dosage 4 (Starter/Grower) |
|---|---|---|---|
| 0 | NEG | None | – |
| 1 | NAR | Narasin (100 g narasin/kg additive) | 700/700 |
| 2 | PRO | Lactobacillus farciminis CNMA 67/4R strain (1 × 109 cfu/gram additive) | 500/500 |
| 3 | PRO | Bacillus subtilis PB6 strain (2 × 108 cfu/gram additive) | 500/500 |
| 4 | PRO | One Bacillus subtilis strain, material no. 671265 (1.6 × 109 cfu/gram additive) | 500/500 |
| 5 | PRO/PRE | Enterococcus faecium DSM 16211 (jejunum isolate), Bifidobacterium animalis DSM 16284 (ileum isolate), Lactobacillus salivarus DSM 16351 (caeca isolate) with mix ratio 3:1:6 (total cfu/gram: 2 × 108), plant-derived fructooligosaccharides from inulin | 1000/1000 |
| 6 | PRE | Saccharomyces cerevisiae cell wall extracts (including typ. 25% β-1,3/1,6 glucans and min. 24% mannanoligosaccharides) | 1000/1000 |
| 7 | PRE | Saccharomyces cerevisiae cell wall extracts (including min. 60% purified β-1,3/1,6 glucans) | 250/250 |
| 8 | OA/PFA | Benzoic acid (80%–83%) and a blend of essential oils (including thymol 1.0%–1.9%, eugenol 0.5%–1.0%, and piperine 0.05%–0.1%) | 300/300 |
| 9 | PFA | Essential oil blend (min. 31.9%, including carvacrol, thymol, anethol and limonene) | 150/150 |
| 10 | OA | Medium-chain fatty acids (C6, C8 and C10) | 1600/1600 |
| 11 | OA/PFA | Short- and medium chain fatty acids (including C4 and C12), phenolic compound and organic acids | 1500/1500 |
| 12 | OA | Tri- and diglycerides of butyric acid (C4) | 1000/1000 |
| 13 | OA | Diformate derived from C1 (57% Na-formate, 39% formic acid) | 3000/3000 |
| 14 | OA | Lactylates (C12 and C14 esterified with lactic acid) | 750/750 |
| 15 | OA/PFA | Short- and medium-chain fatty acids (including monoglycerides of C3, C4, C8 and C10) and essential oils (mainly cinnamon aldehyde) | 3000/2500 |
| 16 | PRE | Dehydrated Saccharomyces cerevisiae culture with whole cells, metabolites and medium nutrients | 1250/620 |
| 17 | OA/PFA | Glycerol-esterified short- and medium-chain fatty acids (including C3, C4, C8 and C10) and 6% phytogenics (including essential oils, saponins and bitter and pungent substances) | 750/750 |
| 18 | PFA | Phytogenics including alkaloids, saponins, thymol and glyco-components derived from Yucca plants | 2000/1000 |
| 19 | PRE | Saccharomyces cerevisiae cell wall extracts (primarily mannanoligosaccharides) | 800/400 |
| 20 | PFA | Tall oil fatty acids from coniferous trees, including resin acids (8%–9%) | 1000/1000 |
| 21 | PFA | Oleoresins from turmeric (Curcuma longa) (4.4%) and chili peppers (genus Capsicum) (4.4%) | 100/100 |
| 22 | PRO/PRE | Active components of ID 5 | 1000/1000 |
| +PRE | and ID 7 | 250/250 | |
| 23 | PRE | Active components of ID 7 | 250/250 |
| +PFA | and ID 21 | 100/100 | |
| 24 | OA/PFA | Active components of ID 11 | 1500/1000 |
| +PRE | and ID 16 | 1250/625 | |
| 25 | OA | Active components of ID 13 | 3000/2000 |
| +PRE | and ID 16 | 1250/625 |
| Group | Days 0–14 | Days 14–28 | Days 0–28 | CP Counts log10 cfu/g | |||
|---|---|---|---|---|---|---|---|
| BWG g | FCR g/g | BWG g | FCR g/g | BWG g | FCR g/g | ||
| NEG 2 | 474 ± 4 | 1.098 ± 0.006 | 1240 ± 9 | 1.338 ± 0.005 | 1714 ± 11 | 1.248 ± 0.003 | 6.09 ± 0.14 |
| NAR 3 | 488 ± 6 p = 0.032 | 1.064 ± 0.008 p < 0.001 | 1337 ± 12 p < 0.001 | 1.273 ± 0.007 p < 0.001 | 1825 ± 16 p < 0.001 | 1.192 ± 0.005 p < 0.001 | 2.92 ± 0.20 p < 0.001 |
| ATAs 4 | 478 ± 5 p = 0.419 | 1.087 ± 0.006 p = 0.079 | 1275 ± 10 p < 0.001 | 1.317 ± 0.006 p < 0.001 | 1753 ± 12 p = 0.002 | 1.232 ± 0.004 p < 0.001 | 5.63 ± 0.16 p = 0.005 |
| ICC 5 | 0.35 | 0.61 | 0.43 | 0.35 | 0.42 | 0.28 | 0.08 |
| Class | Days 0–14 | Days 14–28 | Days 0–28 | CP Counts log10 cfu/g | |||
|---|---|---|---|---|---|---|---|
| BWG g | FCR g/g | BWG g | FCR g/g | BWG g | FCR g/g | ||
| NEG 2 | 474 ± 4 | 1.098 ± 0.006 | 1240 ± 9 | 1.338 ± 0.005 | 1714 ± 11 | 1.248 ± 0.003 | 6.09 ± 0.14 |
| NAR 3 | 488 ± 6 p = 0.032 | 1.064 ± 0.008 p < 0.001 | 1337 ± 12 p < 0.001 | 1.273 ± 0.007 p < 0.001 | 1825 ± 16 p < 0.001 | 1.192 ± 0.005 p < 0.001 | 2.92 ± 0.20 p < 0.001 |
| PRO 4 | 455 ± 8 p = 0.012 | 1.113 ± 0.009 p = 0.118 | 1283 ± 15 p = 0.004 | 1.302 ± 0.009 p < 0.001 | 1736 ± 19 p = 0.239 | 1.232 ± 0.006 p = 0.004 | 5.49 ± 0.25 p = 0.017 |
| PRE 5 | 479 ± 7 p = 0.496 | 1.095 ± 0.009 p = 0.761 | 1288 ± 14 p < 0.001 | 1.305 ± 0.008 p < 0.001 | 1767 ± 18 p = 0.003 | 1.229 ± 0.005 p < 0.001 | 5.70 ± 0.23 p = 0.092 |
| PFA 5 | 480 ± 7 p = 0.375 | 1.086 ± 0.009 p = 0.152 | 1247 ± 14 p = 0.610 | 1.323 ± 0.008 p = 0.062 | 1727 ± 17 p = 0.457 | 1.233 ± 0.005 p = 0.004 | 5.18 ± 0.23 p < 0.001 |
| OA 5 | 490 ± 7 p = 0.025 | 1.062 ± 0.009 p < 0.001 | 1288 ± 14 p < 0.001 | 1.325 ± 0.008 p = 0.114 | 1778 ± 17 p < 0.001 | 1.232 ± 0.005 p = 0.002 | 5.74 ± 0.23 p = 0.130 |
| MIX 6 | 479 ± 6 p = 0.339 | 1.087 ± 0.007 p = 0.103 | 1275 ± 11 p = 0.002 | 1.320 ± 0.007 p = 0.007 | 1754 ± 14 p = 0.005 | 1.234 ± 0.004 p = 0.001 | 5.79 ± 0.19 p = 0.113 |
| ICC 7 | 0.35 | 0.61 | 0.43 | 0.35 | 0.42 | 0.28 | 0.08 |
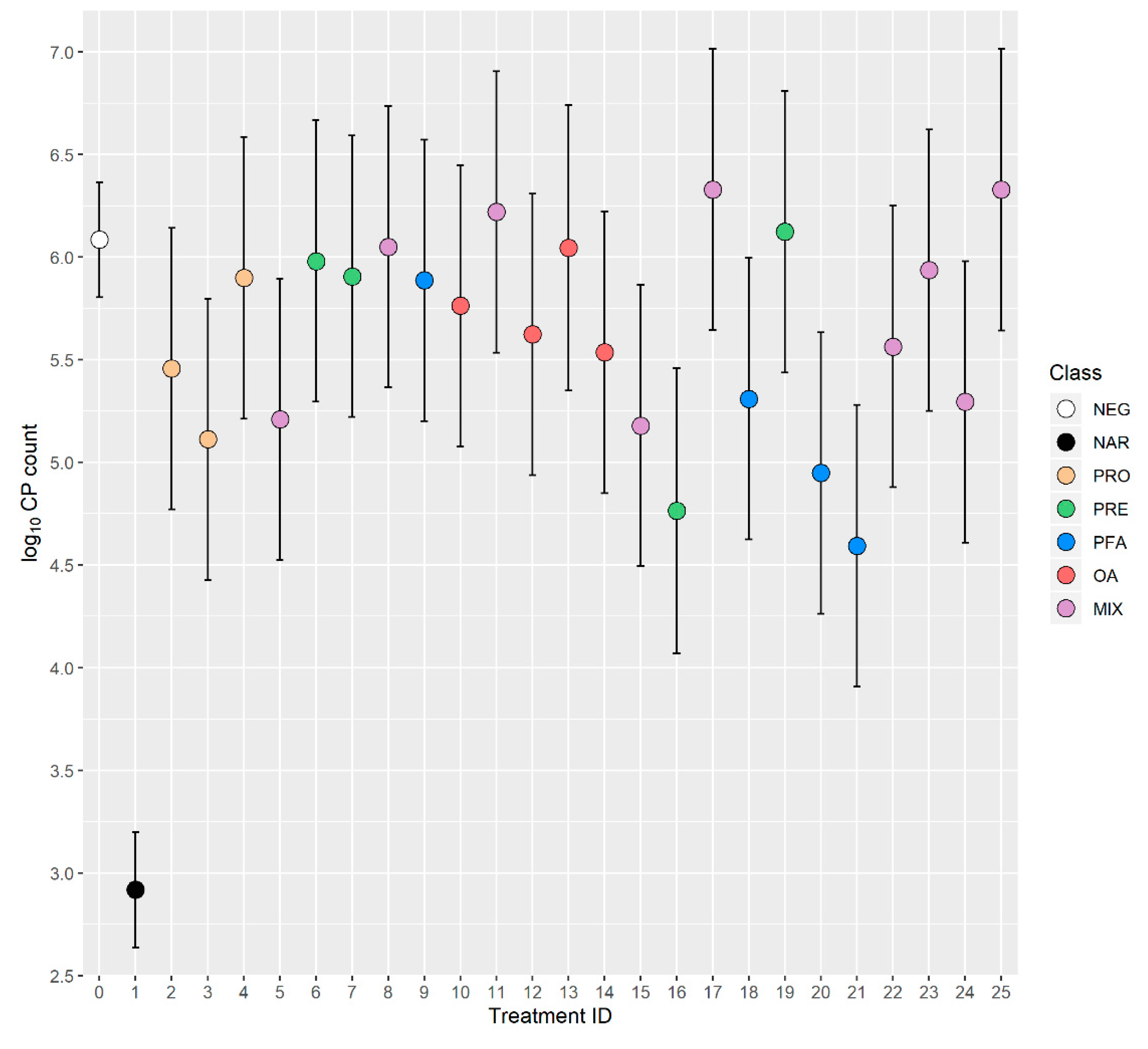
| ID-Class | Days 0–14 | Days 14–28 | Days 0–28 | CP Counts log10 cfu/g | |||
|---|---|---|---|---|---|---|---|
| BWG g | FCR g/g | BWG g | FCR g/g | BWG g | FCR g/g | ||
| 0-NEG 2 | 474 ± 4 | 1.098 ± 0.006 | 1240 ± 9 | 1.338 ± 0.005 | 1714 ± 11 | 1.248 ± 0.003 | 6.09 ± 0.14 |
| 1-NAR 3 | 488 ± 6 p = 0.032 | 1.064 ± 0.008 p < 0.001 | 1337 ± 12 p < 0.001 | 1.273 ± 0.007 p < 0.001 | 1825 ± 16 p < 0.001 | 1.192 ± 0.005 p < 0.001 | 2.92 ± 0.20 p < 0.001 |
| 2-PRO 4 | 452 ± 11 p = 0.049 | 1.118 ± 0.014 p = 0.153 | 1285 ± 22 p = 0.044 | 1.305 ± 0.012 p = 0.007 | 1735 ± 29 p = 0.455 | 1.236 ± 0.008 p = 0.120 | 5.46 ± 0.38 p = 0.097 |
| 3-PRO 4 | 451 ± 11 p = 0.044 | 1.110 ± 0.014 p = 0.383 | 1273 ± 22 p = 0.132 | 1.307 ± 0.012 p = 0.012 | 1723 ± 29 p = 0.740 | 1.235 ± 0.008 p = 0.094 | 5.11 ± 0.38 p = 0.010 |
| 4-PRO 4 | 462 ± 11 p = 0.274 | 1.111 ± 0.014 p = 0.357 | 1290 ± 22 p = 0.024 | 1.295 ± 0.012 p < 0.001 | 1751 ± 29 p = 0.198 | 1.224 ± 0.008 p = 0.002 | 5.90 ± 0.38 p = 0.623 |
| 5-MIX 4 | 472 ± 11 p = 0.872 | 1.084 ± 0.014 p = 0.304 | 1268 ± 22 p = 0.207 | 1.329 ± 0.012 p = 0.459 | 1739 ± 29 p = 0.378 | 1.244 ± 0.008 p = 0.579 | 5.21 ± 0.38 p = 0.021 |
| 6-PRE 4 | 476 ± 11 p = 0.854 | 1.112 ± 0.014 p = 0.324 | 1305 ± 22 p = 0.004 | 1.280 ± 0.012 p < 0.001 | 1782 ± 29 p = 0.023 | 1.216 ± 0.008 p < 0.001 | 5.98 ± 0.38 p = 0.782 |
| 7-PRE 4 | 470 ± 11 p = 0.731 | 1.106 ± 0.014 p = 0.544 | 1311 ± 22 p = 0.002 | 1.269 ± 0.012 p < 0.001 | 1781 ± 29 p = 0.018 | 1.211 ± 0.008 p < 0.001 | 5.91 ± 0.38 p = 0.637 |
| 8-MIX 4 | 469 ± 11 p = 0.672 | 1.093 ± 0.014 p = 0.708 | 1293 ± 22 p = 0.016 | 1.280 ± 0.012 p < 0.001 | 1763 ± 29 p = 0.086 | 1.208 ± 0.008 p < 0.001 | 6.05 ± 0.38 p = 0.928 |
| 9-PFA 4 | 459 ± 11 p = 0.178 | 1.108 ± 0.014 p = 0.480 | 1288 ± 22 p = 0.030 | 1.284 ± 0.012 p < 0.001 | 1747 ± 29 p = 0.243 | 1.221 ± 0.008 p < 0.001 | 5.89 ± 0.38 p = 0.600 |
| 10-OA 4 | 499 ± 11 p = 0.029 | 1.073 ± 0.014 p = 0.070 | 1280 ± 22 p = 0.072 | 1.327 ± 0.012 p = 0.368 | 1780 ± 29 p = 0.021 | 1.233 ± 0.008 p = 0.051 | 5.76 ± 0.38 p = 0.395 |
| 11-MIX 4 | 511 ± 11 p = 0.001 | 1.037 ± 0.014 p < 0.001 | 1335 ± 22 p < 0.001 | 1.317 ± 0.012 p = 0.092 | 1847 ± 29 p < 0.001 | 1.215 ± 0.008 p < 0.001 | 6.22 ± 0.38 p = 0.720 |
| 12-OA 4 | 494 ± 11 p = 0.078 | 1.038 ± 0.014 p < 0.001 | 1287 ± 22 p = 0.034 | 1.324 ± 0.012 p = 0.252 | 1782 ± 29 p = 0.017 | 1.223 ± 0.008 p = 0.001 | 5.62 ± 0.38 p = 0.222 |
| 13-OA 4 | 501 ± 11 p = 0.019 | 1.028 ± 0.014 p < 0.001 | 1318 ± 22 p < 0.001 | 1.311 ± 0.012 p = 0.031 | 1820 ± 29 p < 0.001 | 1.208 ± 0.008 p < 0.001 | 6.05 ± 0.38 p = 0.918 |
| 14-OA 4 | 465 ± 11 p = 0.423 | 1.108 ± 0.014 p = 0.469 | 1266 ± 22 p = 0.237 | 1.340 ± 0.012 p = 0.884 | 1730 ± 29 p = 0.567 | 1.263 ± 0.008 p = 0.058 | 5.54 ± 0.38 p = 0.147 |
| 15-MIX 4 | 476 ± 11 p = 0.845 | 1.097 ± 0.014 p = 0.939 | 1278 ± 22 p = 0.085 | 1.338 ± 0.012 p = 0.977 | 1754 ± 29 p = 0.165 | 1.255 ± 0.008 p = 0.344 | 5.18 ± 0.38 p = 0.017 |
| 16-PRE 4 | 485 ± 11 p = 0.352 | 1.085 ± 0.014 p = 0.346 | 1304 ± 22 p = 0.004 | 1.335 ± 0.012 p = 0.822 | 1788 ± 29 p = 0.009 | 1.251 ± 0.008 p = 0.669 | 4.76 ± 0.38 p < 0.001 |
| 17-MIX 4 | 458 ± 11 p = 0.157 | 1.105 ± 0.014 p = 0.588 | 1228 ± 22 p = 0.593 | 1.354 ± 0.012 p = 0.185 | 1685 ± 29 p = 0.316 | 1.270 ± 0.008 p = 0.004 | 6.33 ± 0.38 p = 0.518 |
| 18-PFA 4 | 491 ± 11 p = 0.132 | 1.067 ± 0.014 p = 0.025 | 1226 ± 22 p = 0.524 | 1.353 ± 0.012 p = 0.216 | 1717 ± 29 p = 0.926 | 1.243 ± 0.008 p = 0.552 | 5.31 ± 0.38 p = 0.040 |
| 19-PRE 4 | 485 ± 11 p = 0.371 | 1.078 ± 0.014 p = 0.158 | 1229 ± 22 p = 0.624 | 1.336 ± 0.012 p = 0.850 | 1713 ± 29 p = 0.971 | 1.237 ± 0.008 p = 0.156 | 6.12 ± 0.38 p = 0.918 |
| 20-PFA 4 | 486 ± 11 p = 0.301 | 1.091 ± 0.014 p = 0.590 | 1228 ± 22 p = 0.592 | 1.334 ± 0.012 p = 0.748 | 1713 ± 29 p = 0.987 | 1.242 ± 0.008 p = 0.428 | 4.95 ± 0.38 p = 0.003 |
| 21-PFA 4 | 485 ± 11 p = 0.330 | 1.077 ± 0.014 p = 0.128 | 1246 ± 22 p = 0.799 | 1.321 ± 0.012 p = 0.170 | 1730 ± 29 p = 0.566 | 1.226 ± 0.008 p = 0.004 | 4.59 ± 0.38 p < 0.001 |
| 22-MIX 4 | 486 ± 11 p = 0.301 | 1.089 ± 0.014 p = 0.502 | 1270 ± 22 p = 0.179 | 1.320 ± 0.012 p = 0.146 | 1755 ± 29 p = 0.147 | 1.231 ± 0.008 p = 0.028 | 5.56 ± 0.38 p = 0.168 |
| 23-MIX 4 | 484 ± 11 p = 0.355 | 1.083 ± 0.014 p = 0.273 | 1251 ± 22 p = 0.612 | 1.327 ± 0.012 p = 0.378 | 1736 ± 29 p = 0.448 | 1.235 ± 0.008 p = 0.090 | 5.94 ± 0.38 p = 0.694 |
| 24-MIX 4 | 494 ± 11 p = 0.078 | 1.086 ± 0.014 p = 0.386 | 1292 ± 22 p = 0.018 | 1.307 ± 0.012 p = 0.014 | 1786 ± 29 p = 0.011 | 1.222 ± 0.008 p < 0.001 | 5.30 ± 0.38 p = 0.037 |
| 25-MIX 4 | 464 ± 11 p = 0.394 | 1.105 ± 0.014 p = 0.610 | 1255 ± 22 p = 0.489 | 1.311 ± 0.012 p = 0.028 | 1720 ± 29 p = 0.844 | 1.229 ± 0.008 p = 0.013 | 6.33 ± 0.38 p = 0.518 |
| ICC 5 | 0.35 | 0.61 | 0.43 | 0.35 | 0.42 | 0.28 | 0.08 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Granstad, S.; Kristoffersen, A.B.; Benestad, S.L.; Sjurseth, S.K.; David, B.; Sørensen, L.; Fjermedal, A.; Edvardsen, D.H.; Sanson, G.; Løvland, A.; et al. Effect of Feed Additives as Alternatives to In-feed Antimicrobials on Production Performance and Intestinal Clostridium perfringens Counts in Broiler Chickens. Animals 2020, 10, 240. https://doi.org/10.3390/ani10020240
Granstad S, Kristoffersen AB, Benestad SL, Sjurseth SK, David B, Sørensen L, Fjermedal A, Edvardsen DH, Sanson G, Løvland A, et al. Effect of Feed Additives as Alternatives to In-feed Antimicrobials on Production Performance and Intestinal Clostridium perfringens Counts in Broiler Chickens. Animals. 2020; 10(2):240. https://doi.org/10.3390/ani10020240
Chicago/Turabian StyleGranstad, Silje, Anja B. Kristoffersen, Sylvie L. Benestad, Siri K. Sjurseth, Bruce David, Line Sørensen, Arnulf Fjermedal, Dag H. Edvardsen, Gorm Sanson, Atle Løvland, and et al. 2020. "Effect of Feed Additives as Alternatives to In-feed Antimicrobials on Production Performance and Intestinal Clostridium perfringens Counts in Broiler Chickens" Animals 10, no. 2: 240. https://doi.org/10.3390/ani10020240
APA StyleGranstad, S., Kristoffersen, A. B., Benestad, S. L., Sjurseth, S. K., David, B., Sørensen, L., Fjermedal, A., Edvardsen, D. H., Sanson, G., Løvland, A., & Kaldhusdal, M. (2020). Effect of Feed Additives as Alternatives to In-feed Antimicrobials on Production Performance and Intestinal Clostridium perfringens Counts in Broiler Chickens. Animals, 10(2), 240. https://doi.org/10.3390/ani10020240