Psychological Stress, Its Reduction, and Long-Term Consequences: What Studies with Laboratory Animals Might Teach Us about Life in the Dog Shelter

Simple Summary
Abstract
1. Introduction
2. HPA Responses and Social Buffering in the Shelter
2.1. How Stressors Like Those in the Shelter Affect HPA Activity
2.2. Brief History of Social Buffering in Nonhuman Primates
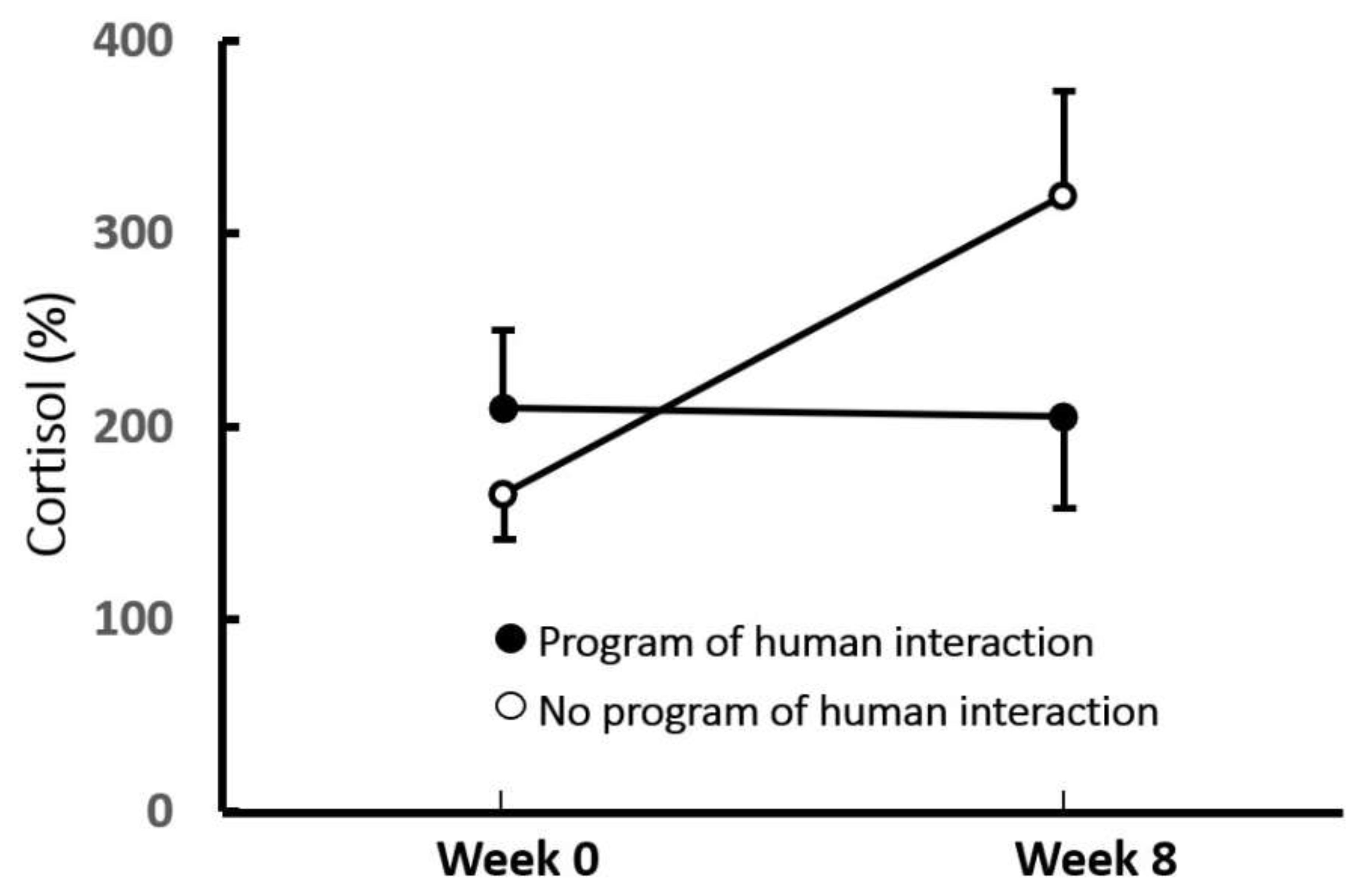
2.3. Social Buffering of HPA Responses in Dogs
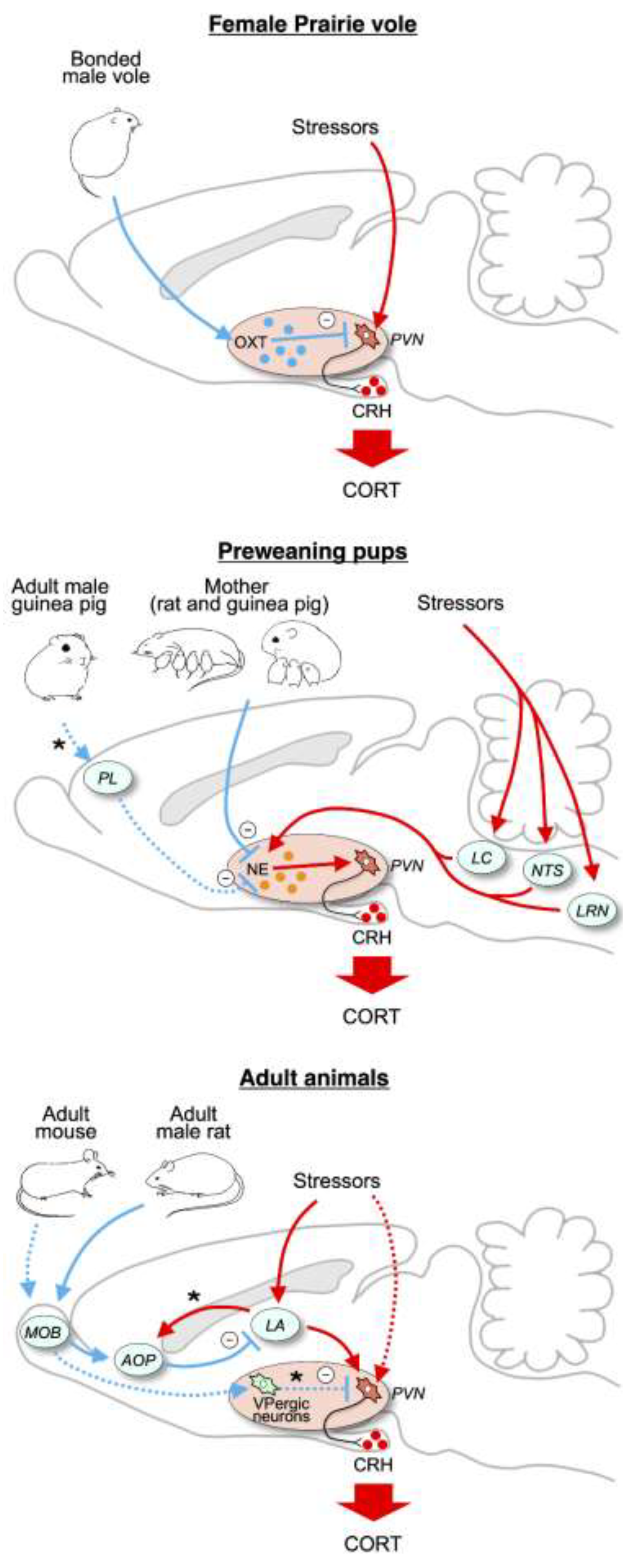
2.4. Mechanism of Social Buffering of HPA Responses
3. Stress Effects on Behavior
3.1. The Challenge of Detecting Behavioral Consequences of Stress in Shelter Dogs
3.2. Reduction in Fear-Induced Aggression in the Shelter
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Finnell, J.E.; Lombard, C.M.; Padi, A.R.; Moffitt, C.M.; Wilson, L.B.; Wood, C.S.; Wood, S.K. Physical versus psychological social stress in male rats reveals distinct cardiovascular, inflammatory, and behavioral consequences. PLoS ONE 2017, 12, e0172868. [Google Scholar] [CrossRef]
- Manouze, H.; Ghestem, A.; Poillerat, V.; Bennis, M.; Ba-M’hamed, S.; Benoliel, J.J.; Becker, C.; Bernard, C. Effects of single cage housing on stress, cognitive, and seizure parameters in the rat and mouse pilocarpine models of epilepsy. eNeuro 2019, 6. [Google Scholar] [CrossRef]
- Monk, C.; Lugo-Candelas, C.; Trumpff, C. Prenatal developmental origins of future psychopathology: Mechanisms and pathways. Ann. Rev. Clin. Psychol. 2019, 15, 317–344. [Google Scholar] [CrossRef]
- Naqvi, F.; Haider, S.; Batool, Z.; Perveen, T.; Haleem, D.J. Sub-chronic exposure to noise affects locomotor activity and produces anxiogenic and depressive like behavior in rats. Pharmacol. Rep. 2012, 64, 64–69. [Google Scholar] [CrossRef]
- Schneider, R.L.; Schiml, P.A.; Deak, T.; Hennessy, M.B. Persistent sensitization of depressive-like behavior and thermogenic response during maternal separation in pre- and post-weaning guinea pigs. Dev. Psychobiol. 2012, 54, 514–522. [Google Scholar] [CrossRef]
- Schroeder, A.; Notaras, M.; Du, X.; Hill, R.A. On the developmental timing of stress: Delineating sex-specific effects of stress across development on adult behavior. Brain Sci. 2018, 8, 121. [Google Scholar] [CrossRef]
- Verbitski, A.; Dodfel, D.; Zhang, N. Rodent models of post-traumatic stress disorder: Behavioral assessment. Transl. Psychiatry 2020, 14, 132. [Google Scholar] [CrossRef]
- Walker, C.-D.; Bath, K.G.; Joels, M.; Korosi, A.; Larauche, M.; Lucassen, J.; Morris, M.J.; Raineki, C.; Roth, T.L.; Sullivan, R.M.; et al. Chronic early life stress induced by limited bedding and nesting (LBN) material in rodents: Critical considerations of methodology, outcomes, and translational potential. Stress 2017, 20, 421–448. [Google Scholar] [CrossRef]
- Dudley, E.S.; Schiml, P.A.; Hennessy, M.B. White blood cells, parasite prevalence, and plasma cortisol levels of dogs in a county animal shelter: Changes over days and impact of a program of repeated human interaction. JAVMA 2015, 247, 1289–1298. [Google Scholar] [CrossRef]
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of stress throughout the lifespan on the brain, behaviour, and cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef]
- Raymond, C.; Marin, M.-F.; Majeur, D.; Lupien, S. Early child adversity and psychopathology in adulthood: HPA axis and cognitive dysregulations as potential mechanisms. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 85, 152–160. [Google Scholar] [CrossRef]
- Mason, J.W. A Historical view of the stress field Part II. J. Hum. Stress 1975, 1, 22–36. [Google Scholar] [CrossRef]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the hypothalamic-pituitary-adrenocortical stress response. Compr. Physiol. 2016, 6, 603–621. [Google Scholar]
- Selye, H.A. A syndrome produced by diverse nocuous agents. Nature 1936, 138, 32. [Google Scholar] [CrossRef]
- Gunnar, M.R.; Hostinar, C.E.; Sanchez, M.M.; Tottenham, N.; Sullivan, R. Parental buffering of fear and stress neurobiology: Reviewing parallels across rodent, monkey, and human models. Soc. Neurosci. 2015, 10, 474–478. [Google Scholar] [CrossRef]
- Sanchez, M.M.; McCormack, K.M.; Howell, B.R. Social buffering of stress responses in nonhuman primates: Maternal regulation of the development of emotional regulatory brain circuits. Soc. Neurosci. 2015, 10, 512–526. [Google Scholar] [CrossRef]
- Hennessy, M.B.; Kaiser, S.; Sachser, N. Social buffering of the stress response: Diversity, mechanisms, and functions. Front. Neuroendocrinol. 2009, 30, 470–482. [Google Scholar] [CrossRef]
- Hill, S.D.; McCormack, S.A.; Mason, W.A. Effects of artificial mothers and visual experience on adrenal responsiveness of infant monkeys. Dev. Psychobiol. 1973, 6, 421–429. [Google Scholar] [CrossRef]
- Levine, S.; Johnson, D.F.; Gonzalez, C.A. Behavioral and hormonal responses to separation in infant rhesus monkeys and mothers. Behav. Neurosci. 1985, 99, 399–410. [Google Scholar] [CrossRef]
- Smotherman, W.P.; Hunt, L.E.; McGinnis, L.E.; Levine, S. Mother-infant separation in group-living rhesus macaques: A hormonal analysis. Dev. Psychobiol. 1979, 12, 211–217. [Google Scholar] [CrossRef]
- Coe, C.L.; Mendoza, S.P.; Smotherman, W.P.; Levine, S. Mother-infant attachment in the squirrel monkey: Adrenal response to separation. Behav. Biol. 1978, 22, 256–263. [Google Scholar] [CrossRef]
- Mendoza, S.P.; Smotherman, W.P.; Miner, M.T.; Kaplan, J.; Levine, S. Pituitary-adrenal response to separation in mother and infant squirrel monkeys. Dev. Psychobiol. 1978, 11, 169–175. [Google Scholar] [CrossRef]
- Hennessy, M.B. Effects of social partners on pituitary-adrenal activity during novelty exposure in adult female squirrel monkeys. Physiol. Behav. 1986, 38, 803–807. [Google Scholar] [CrossRef]
- Hennessy, M.B.; Mendoza, S.P.; Kaplan, J.N. Behavior and plasma cortisol following brief peer separation in juvenile squirrel monkeys. Am. J. Primatol. 1982, 3, 143–151. [Google Scholar] [CrossRef]
- Mendoza, S.P.; Mason, W.A. Parental division of labour and differentiation of attachments in a monogamous primate (Callicebus moloch). Anim. Behav. 1986, 34, 1336–1347. [Google Scholar] [CrossRef]
- Hoffman, K.A.; Mendoza, S.P.; Hennessy, M.B.; Mason, W.A. Responses of infant titi monkeys, Calicebus moloch, to removal of one or both parents: Evidence for paternal attachment. Dev. Psychobiol. 1995, 28, 399–407. [Google Scholar] [CrossRef]
- Kiyokawa, Y.; Hiroshima, S.; Takeuchi, Y.; Mori, Y. Social buffering reduces male rats’ behavioral and corticosterone responses to a conditioned stimulus. Horm. Behav. 2014, 65, 114–118. [Google Scholar] [CrossRef]
- Kiyokawa, Y.; Hennessy, M.B. Comparative studies of social buffering: A consideration of approaches, terminology, and pitfalls. Neurosci. Biobehav. Rev. 2018, 86, 131–141. [Google Scholar] [CrossRef]
- Gunnar, M.R.; Hostinar, C.E. The social buffering of the hypothalamic-pituitary- adrenocortical axis in humans: Developmental and experiential determinants. Soc. Neurosci. 2015, 10, 479–488. [Google Scholar] [CrossRef]
- Tuber, D.S.; Hennessy, M.B.; Sanders, S.; Miller, J.A. Behavioral and glucocorticoid responses of adult domestic dogs (Canis familiaris) to companionship and social separation. J. Comp. Psychol. 1996, 110, 103–108. [Google Scholar] [CrossRef]
- Hennessy, M.B.; Davis, H.N.; Williams, M.T.; Mellott, C.; Douglas, C.W. Plasma cortisol levels of dogs at a county animal shelter. Physiol. Behav. 1997, 62, 485–490. [Google Scholar] [CrossRef]
- Blackwell, E.-J.; Bodnariu, A.; Tyson, J.; Bradshaw, J.W.S.; Casey, R.A. Rapid shaping of behavior associated with high urinary cortisol in domestic dogs. Appl. Anim. Behav. Sci. 2010, 124, 113–120. [Google Scholar] [CrossRef]
- Coppola, C.L.; Grandin, T.; Enns, M. Human interaction and cortisol: Can human contact reduce stress for shelter dogs. Physiol. Behav. 2006, 87, 537–541. [Google Scholar] [CrossRef]
- Hennessy, M.B.; Voith, V.L.; Hawke, J.L.; Young, T.L.; Centrone, J.; McDowell, A.L.; Linden, F.; Davenport, G.M. Effects of a program of human interaction and alterations in diet composition on activity of the hypothalamic-pituitary-adrenal axis in dogs housed in a public animal shelter. JAVMA 2002, 221, 65–71. [Google Scholar] [CrossRef]
- Hiby, E.F.; Rooney, J.J.; Bradshaw, J.W.S. Behavioural and physiological responses of dogs entering re-homing kennels. Physiol. Behav. 2006, 89, 385–389. [Google Scholar] [CrossRef]
- Menor-Campos, D.J.; Molleda-Carbonell, J.M.; López-Rodríuez, R. Effects of exercise and human contact on animal welfare in a dog shelter. Vet. Rec. 2011, 169, 388. [Google Scholar] [CrossRef]
- Stephen, J.M.; Ledger, R.A. A longitudinal evaluation of urinary cortisol in kenneled dogs, Canis familiaris. Physiol. Behav. 2006, 87, 911–916. [Google Scholar] [CrossRef]
- Hennessy, M.B.; Williams, M.T.; Miller, D.D.; Douglas, C.W.; Voith, V.L. Influence of male and female petters on plasma cortisol and behaviour: Can human interaction reduce the stress of dogs in a public animal shelter? Appl. Anim. Behav. Sci. 1998, 61, 63–77. [Google Scholar] [CrossRef]
- Shiverdecker, M.; Schiml, P.A.; Hennessy, M.B. Human interaction moderates plasma cortisol and behavioral responses of dogs to shelter housing. Physiol. Behav. 2013, 109, 75–79. [Google Scholar] [CrossRef]
- Willen, R.M.; Mutwill, A.; MacDonald, L.J.; Schiml, P.A.; Hennessy, M.B. Factors determining the effect of human interaction on the cortisol levels of shelter dogs. Appl. Animal Behav. Sci. 2017, 186, 41–48. [Google Scholar] [CrossRef]
- Gunter, L.M.; Feuerbacher, E.N.; Gilchrist, R.J.; Wynne, C.D.L. Evaluating the effects of a temporary fostering program on shelter dog welfare. PeerJ 2019, 7, e6620. [Google Scholar] [CrossRef]
- Nagasawa, M.; Mitsui, S.; En, S.; Ohtani, N.; Ohta, M.; Sakuma, Y.; Onaka, T.; Mogi, K.; Kikusui, T. Oxytocin-gaze positive loop and the coevolution of human-dog bonds. Science 2015, 48, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Uvnäs-Moberg, K.; Handlin, L.; Petersson, M. Self-soothing behaviors with particular reference to oxytocin release induced by non-noxious sensory stimulation. Front. Psychol. 2015, 5. [Google Scholar] [CrossRef]
- Carter, C.S.; Williams, J.R.; Witt, D.M.; Insel, T.R. Oxytocin and social bonding. Ann. N. Y. Acad. Sci. 1992, 652, 204–211. [Google Scholar] [CrossRef]
- Young, L.J.; Flannagan-Cato, L.M. Editorial comment: Oxytocin, vasopressin, and social behavior. Horm. Behav. 2012, 61, 227–229. [Google Scholar] [CrossRef]
- Barrera, G.; Dzik, V.; Cavalli, C.; Bentosela, M. Effect of intranasal oxytocin administration on human-directed social behaviors in shelter and pet dogs. Front. Psychol. 2018, 9, 2227. [Google Scholar] [CrossRef]
- Romero, T.; Nagasawa, M.; Mogi, K.; Hasegawa, T.; Kikusui, T. Oxytocin promotes social bonding in dogs. Proc. Natl. Acad. Sci. USA 2014, 111, 9085–9090. [Google Scholar] [CrossRef]
- Buttner, A.P. Neurobiological underpinnings of dogs’ human-like social competence: How interactions between the stress response systems and oxytocin mediate dogs’ social skills. Neurosci. Biobehav. Rev. 2016, 71, 198–214. [Google Scholar] [CrossRef]
- Smith, A.S.; Tabbaa, M.; Lei, K.; Eastham, P.; Butler, M.J.; Linton, L.; Altshuler, R.; Liu, Y.; Wang, Z. Local oxytocin tempers anxiety by activating GABAA receptors in the hypothalamic paraventricular nucleus. Psychoneuroendocrinology 2016, 63, 50–58. [Google Scholar] [CrossRef]
- Smith, A.S.; Wang, Z. Hypothalamic oxytocin mediates social buffering of the stress response. Biol. Psychiatry 2014, 76, 281–288. [Google Scholar] [CrossRef]
- Shionoya, K.; Moriceau, S.; Bradstock, P.; Sullivan, R.M. Maternal attenuation of hypothalamic paraventricular nucleus norepinephrine switches avoidance learning to preference learning in preweanling rat pups. Horm. Behav. 2007, 52, 391–400. [Google Scholar] [CrossRef]
- Hennessy, M.B.; Ritchey, R.L. Hormonal and behavioral attachment responses in infant guinea pigs. Dev. Psychobiol. 1987, 20, 613–625. [Google Scholar] [CrossRef]
- Harvey, A.T.; Moore, H.; Lucot, J.B.; Hennessy, M.B. Monoamine activity in anterior hypothalamus of guinea pig pups separated from their mothers. Behav. Neurosci. 1994, 108, 171–176. [Google Scholar] [CrossRef]
- Hennessy, M.B.; Watansriyakul, W.T.; Price, B.C.; Bertke, A.S.; Schiml, P.A. Adult males buffer the cortisol response of young guinea pigs: Changes with age, mediation by behavior, and comparison with prefrontal activity. Horm. Behav. 2018, 98, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, H.F.; Bruestle, A.; Bodie, B.; Dolgas, C.M.; Herman, J.P. The medial prefrontal cortex differentially regulates stress-induced c-fos expression in the forebrain depending on the type of stressor. Eur. J. Neurosci. 2003, 18, 2357–2364. [Google Scholar] [CrossRef]
- Kiyokawa, Y.; Takeuchi, Y.; Nishihara, M.; Mori, Y. Main olfactory system mediates social buffering of conditioned fear responses in male rats. Eur. J. Neurosci. 2009, 29, 777–785. [Google Scholar] [CrossRef]
- Kiyokawa, Y.; Wakabayashi, Y.; Takeuchi, Y.; Mori, Y. The neural pathway underlying social buffering of conditioned fear responses in male rats. Eur. J. Neurosci. 2012, 36, 3429–3437. [Google Scholar] [CrossRef]
- Klein, B.; Bautze, V.; Maier, A.M.; Deussing, J.; Breer, H.; Strotmann, J. Activation of the mouse odorant receptor 37 subsystem coincides with a reduction of novel environment-induced activity within the paraventricular nucleus of the hypothalamus. Eur. J. Neurosci. 2015, 41, 793–801. [Google Scholar] [CrossRef]
- Protopopova, A.; Hauser, H.; Goldman, K.J.; Wynne, C.D.L. The effects of exercise and calm interactions on in-kennel behavior of shelter dogs. Behav. Processes 2018, 146, 54–60. [Google Scholar] [CrossRef]
- Wells, D.L.; Hepper, P.G. Prevalence of behaviour problems reported by owners of dogs purchased from an animal rescue shelter. Appl. Anim. Behav. Sci. 2000, 69, 55–65. [Google Scholar] [CrossRef]
- Hodges, T.E.; Baumbach, J.L.; Marcolin, M.L.; Bredewold, R.; Veenema, A.H.; McCormick, C.M. Social instability stress in adolescent male rats reduces social interaction and social recognition performance and increases oxytocin receptor binding. Neuroscience 2017, 359, 172–182. [Google Scholar] [CrossRef] [PubMed]
- McCormick, C.M.; Hodges, T.E.; Simone, J.J. Peer pressures: Social instability stress in adolescence and social deficits in adulthood in a rodent model. Dev. Cog. Neurosci. 2015, 11, 2–11. [Google Scholar] [CrossRef]
- Holland, F.H.; Ganguly, P.; Potter, D.N.; Chartoff, E.H.; Brenhouse, H.C. Early life stress disrupts social behavior and prefrontal cortex parvalbumin interneurons at an earlier time-point in females than in males. Neurosci. Lett. 2014, 566, 131–136. [Google Scholar] [CrossRef]
- Liu, N.; Wang, Y.; An, A.Y.; Banker, C.; Qian, Y.-H.; O’Donnell, J.M. Single housing-induced effects on cognitive impairment and depression-like behavior in male and female mice involve neuroplasticity-related signaling. Eur. J. Neurosci. 2019, 52, 2694–2704. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.L.; Roughton, E.C.; Koehler, A.J.; Lubach, G.R. Growth and development following prenatal stress exposure in primates: An examination of ontogenetic vulnerability. Child Dev. 1999, 70, 263–274. [Google Scholar] [CrossRef]
- Clarke, A.S.; Schneider, M.L. Effects of prenatal stress on behavior in adolescent rhesus monkeys. Ann. N. Y. Acad. Sci. 1997, 807, 490–491. [Google Scholar] [CrossRef]
- Felitti, V.J.; Anda, R.F.; Nordenberg, D.; Williamson, D.F.; Spitz, A.M.; Edwards, V.; Koss, M.P.; Marks, J.S. Relationship of childhood abuse and household dysfunction to many leading causes of death in adults. Am. J. Prev. Med. 1998, 14, 245–258. [Google Scholar] [CrossRef]
- Feigenson, K.A.; Kusnecov, A.W.; Silverstein, S.M. Inflammation and the two-hit hypothesis of schizophrenia. Neurosci. Biobehav. Rev. 2014, 38, 72–93. [Google Scholar] [CrossRef]
- Hammersley, P.; Dias, A.; Todd, G.; Bowen-Jones, K.; Reilly, B.; Bentall, R.P. Childhood trauma and hallucinations in bipolar affective disorder: Preliminary investigation. Brit. J. Psychiatry 2003, 182, 543–547. [Google Scholar] [CrossRef]
- Van Teighem, M.R.; Tottenham, N. Neurobiological programming of early life stress: Functional development of amygdala-prefrontal circuitry and vulnerability for stress-related psychopathology. Curr. Top. Behav. Neurosci. 2018, 38, 117–136. [Google Scholar]
- Gold, P.W.; Goodwin, F.K.; Chrousos, G.P. Clinical and biochemical manifestations of depression: Relation to the neurobiology of stress (Part 2). N. Engl. J. Med. 1988, 319, 413–420. [Google Scholar] [CrossRef]
- Yohn, N.L.; Blendy, J.A. Adolescent chronic unpredictable stress exposure is a sensitive window for long-term changes in adult behavior in mice. Neuropsychopharmacology 2017, 42, 1670–1678. [Google Scholar] [CrossRef]
- Raineki, C.; Opendak, M.; Sarro, E.; Showler, A.; Bui, K.; McEwen, B.S.; Wilson, D.A.; Sullivan, R.M. During infant maltreatment, stress targets hippocampus, but stress with mother targets amygdala and social behavior. Proc. Natl. Acad. Sci. USA 2019, 116, 22821–22832. [Google Scholar] [CrossRef]
- Revest, J.-M.; Di Blasi, F.; Kitchener, P.; Rougé-Pont, F.; Desmedt, A.; Turiault, M.; Tronche, F.; Piazza, P.V. The MAPK pathway and Egr-1 mediate stress-related behavioral effects of glucocorticoids. Nat. Neurosci. 2005, 8, 664–672. [Google Scholar] [CrossRef]
- Perry, R.E.; Rincón-Cortés, M.; Braren, S.H.; Brandes-Aitken, A.N.; Opendak, M.; Pollonini, G.; Chopra, D.; Raver, C.C.; Alberini, C.M.; Blair, C.; et al. Corticosterone administration targeting a hypo-reactive HPA axis rescues a socially-avoidant phenotype in scarcity-adversity reared rats. Dev. Cog. Neurosci. 2019, 40, 100716. [Google Scholar] [CrossRef]
- Dantzer, B.; Newman, A.E.M.; Boonstra, R.; Palme, R.; Boutin, S.; Humphries, M.M.; McAdam, A.G. Density triggers maternal hormones that increase adaptive growth in a wild mammal. Science 2013, 340, 1215–1217. [Google Scholar] [CrossRef] [PubMed]
- Maccari, S.; Darnaudery, M.; Morley-Fletcher, S.; Zuena, A.R.; Cinque, C.; Van Reeth, O. Prenatal stress and long-term consequences: Implications for glucocorticoid hormones. Neurosci. Biobehav. Rev. 2003, 27, 119–127. [Google Scholar] [CrossRef]
- Dettmer, A.M.; Murphy, A.M.; Guitarra, D.; Slonecker, E.; Suomi, S.J.; Rosenberg, K.L.; Novak, M.A.; Meyer, J.S.; Hinde, K. Cortisol in neonatal mother’s milk predicts later infant social and cognitive functioning in rhesus monkeys. Child Dev. 2018, 89, 525–538. [Google Scholar] [CrossRef]
- Gouin, J.-P.; Glaser, R.; Malarkey, W.B.; Beversdorf, D.; Kiecolt-Glaser, J.K. Childhood abuse and inflammatory responses to daily stressors. Ann. Behav. Med. 2012, 44, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Slopen, N.; Loucks, E.B.; Appleton, A.A.; Kawachi, I.; Kubzansky, L.D.; Non, A.L.; Buka, S.; Gilman, S.E. Early origins of inflammation: An examination of prenatal and childhood social adversity in a prospective cohort study. Psychoneuroendocrinology 2015, 51, 403–413. [Google Scholar] [CrossRef]
- Furtado, M.; Katzman, M.A. Neuroinflammatory pathways in anxiety, posttraumatic stress, and obsessive compulsive disorders. Psychiatry Res. 2015, 229, 37–48. [Google Scholar] [CrossRef]
- Slavich, G.M.; Irwin, M.R. From stress to inflammation and major depressive disorder: A social signal transduction theory of depression. Psychol. Bull. 2014, 140, 774–815. [Google Scholar] [CrossRef]
- Depino, A.M. Perinatal inflammation and adult psychopathology: From preclinical models to humans. Semin. Cell Dev. Biol. 2018, 77, 104–114. [Google Scholar] [CrossRef]
- Hennessy, M.B.; Schiml, P.A.; Berberich, K.; Beasley, N.L.; Deak, T. Early attachment disruption, inflammation, and vulnerability for depression in rodent and primate models. Front. Behav. Neurosci. 2019, 12, 314. [Google Scholar] [CrossRef]
- LeDoux, J. Rethinking the emotional brain. Neuron 2012, 73, 653–676. [Google Scholar] [CrossRef]
- Nusslock, R.; Miller, G.E. Early-life adversity and physical and emotional health across the lifespan: A neuroimmune network hypothesis. Biol. Psychiatry 2016, 80, 23–32. [Google Scholar] [CrossRef]
- Tottenham, N.; Hare, T.A.; Millner, A.; Gilhooly, T.; Zevin, J.D.; Casey, B.J. Elevated amygdala response to faces following early deprivation. Dev. Sci. 2011, 14, 190–204. [Google Scholar] [CrossRef]
- Felger, J.C. Imaging the role of inflammation in mood and anxiety disorders. Curr. Neuropharmacol. 2018, 16, 533–558. [Google Scholar] [CrossRef]
- Berens, A.E.; Jensen, S.K.G.; Nelson, C.A., III. Biological embedding of childhood adversity: From physiological mechanisms to clinical implications. BMC Med. 2017, 15, 135. [Google Scholar] [CrossRef]
- Gold, A.L.; Shechner, T.; Farber, M.J.; Spiro, C.N.; Leibenluft, E.; Pine, D.S.; Britton, J.C. Amygdala-cortical connectivity: Associations with anxiety, development, and threat. Depress. Anxiety 2016, 33, 917–926. [Google Scholar] [CrossRef]
- Li, C.T.; Chen, M.H.; Lin, W.C.; Hong, C.J.; Yang, B.H.; Liu, R.S.; Tu, P.C.; Su, T.P. The effects of low-dose ketamine on the prefrontal cortex and amygdala in treatment-resistant depression: A randomized control study. Hum. Brain Mapp. 2016, 37, 1080–1090. [Google Scholar] [CrossRef]
- Liu, H.; Tang, Y.; Womer, F.; Fan, G.; Lu, T.; Driesen, N.; Ren, L.; Wang, Y.; He, Y.; Blumberg, H.P.; et al. Differentiating patterns of amygdala-frontal functional connectivity in schizophrenia and bipolar disorder. Schizophr. Bull. 2014, 40, 469–477. [Google Scholar] [CrossRef]
- Nicholson, A.A.; Rabellino, D.; Densmore, M.; Frewen, P.A.; Paret, C.; Kluetsch, R.; Schmahl, C.; Théberge, J.; Neufeld, R.W.; McKinnon, M.C.; et al. The neurobiology of emotion regulation in posttraumatic stress disorder: Amygdala downregulation via real-time fMRI neurofeedback. Hum. Brain Mapp. 2017, 38, 541–560. [Google Scholar] [CrossRef]
- Protopopova, A. Effects of sheltering on physiology, immune function, behavior, and the welfare of dogs. Physiol. Behav. 2016, 159, 95–103. [Google Scholar] [CrossRef]
- Blackshaw, J.K. An overview of types of aggressive behaviour in dogs and methods of treatment. Appl. Anim. Behav. Sci. 2014, 30, 351–361. [Google Scholar] [CrossRef]
- Moyer, K.E. Kinds of aggression and their physiological basis. Commun. Behav. Biol. 1968, 2, 65–87. [Google Scholar]
- Weiss, E. Meet Your Match SAFER Manual and Training Guide (meetyourmatch@aspca.org); American Society for Prevention of Cruelty to Animals: New York, NY, USA, 2007. [Google Scholar]
- Conley, M.J.; Fisher, A.D.; Hemsworth, P.H. Effects of human contact and toys on the fear responses to humans of shelter dogs. Appl. Anim. Behav. Sci. 2014, 156, 62–69. [Google Scholar] [CrossRef]
- Gácsi, M.; Topál, J.; Miklósi, Á.; Dóka, A.; Csányi, V. Attachment behavior of adult dogs (Canis familaris) living at rescue centers: Forming new bonds. J. Comp. Psych. 2001, 115, 423–431. [Google Scholar] [CrossRef]
- Willen, R.M.; Schiml, P.A.; Hennessy, M.B. Enrichment centered on human interaction moderates fear-induced aggression and increases positive expectancy in fearful shelter dogs. Appl. Anim. Behav. Sci. 2019, 217, 57–62. [Google Scholar] [CrossRef]
- Graham, L.; Wells, D.L.; Hepper, P.G. The influence of olfactory stimulation on the behavior of dogs housed in a rescue shelter. Appl. Anim. Behav. Sci. 2005, 91, 143–153. [Google Scholar] [CrossRef]
- Kogan, L.R.; Schoenfeld-Tacher, R.; Simon, A.A. Behavioral effects of auditory stimulation on kenneled dogs. J. Vet. Behavi. 2012, 7, 268–275. [Google Scholar] [CrossRef]
- Patronek, G.J.; Bradley, J. No better than flipping a coin: Reconsidering canine behavior evaluations in animal shelters. J. Vet. Behav. 2016, 15, 66–77. [Google Scholar] [CrossRef]
- Blas, J.; Bortolotti, G.R.; Tella, J.L.; Baos, R.; Marchant, T.A. Stress response during development predicts fitness in a wild, long lived vertebrate. Proc. Natl. Acad. Sci. USA 2007, 104, 8880–8884. [Google Scholar] [CrossRef] [PubMed]
- Schülke, O.; Ostner, J.; Berghänel, A. Prenatal maternal stress effects on the development of primate social behavior. Behav. Ecol. Sociobiol. 2019, 73, 128. [Google Scholar] [CrossRef]
- Bateson, P.; Gluckman, P.; Hanson, M. The biology of developmental plasticity and the predictive adaptive response hypothesis. J. Physiol. 2014, 592, 2357–2368. [Google Scholar] [CrossRef]
- Sachser, N.; Kaiser, S.; Hennessy, M.B. Behavioural profiles are shaped by social experience: When, how, and why. Philos. Trans. R. Soc. B 2013, 368, 20120344. [Google Scholar] [CrossRef]
- Zimmerman, T.; Kaiser, S.; Hennessy, M.B.; Sachser, N. Adaptive shaping of the behavioural phenotype during adolescence. Proc. R. Soc. Biol. 2017, 284, 20162784. [Google Scholar] [CrossRef]
- Frankenhuis, W.E.; de Weerth, C. Does early-life exposure to stress shape or impair cognition? Curr. Dir. Psychol. Sci. 2013, 22, 407–412. [Google Scholar] [CrossRef]
- Sachser, N.; Zimmerman, T.D.; Hennessy, M.B.; Kaiser, S. Sensitive phases in the development of rodent social behavior. Curr. Opin. Behav. Sci. 2020, 36, 1–8. [Google Scholar]



{kind=link}
{kind=link}
| Subject | Preference for | Buffering by |
|---|---|---|
| Mother | Father > Infant | Father yes; Infant no |
| Father | Mother > Infant | Mother yes: Infant no |
| Infant | Father > Mother | Both yes, Father > Mother |
| Pass | Fail | % Pass | |
|---|---|---|---|
| Experiment 1 | |||
| Fearful | |||
| Enriched*** | 23 | 7 | 77 |
| Control | 10 | 20 | 33 |
| Experiment 2 | |||
| Fearful | |||
| Enriched*** | 15 | 1 | 94 |
| Control | 2 | 14 | 12 |
| Non-Fearful | |||
| Enriched*** | 14 | 2 | 87 |
| Control | 15 | 1 | 94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hennessy, M.B.; Willen, R.M.; Schiml, P.A. Psychological Stress, Its Reduction, and Long-Term Consequences: What Studies with Laboratory Animals Might Teach Us about Life in the Dog Shelter. Animals 2020, 10, 2061. https://doi.org/10.3390/ani10112061
Hennessy MB, Willen RM, Schiml PA. Psychological Stress, Its Reduction, and Long-Term Consequences: What Studies with Laboratory Animals Might Teach Us about Life in the Dog Shelter. Animals. 2020; 10(11):2061. https://doi.org/10.3390/ani10112061
Chicago/Turabian StyleHennessy, Michael B., Regina M. Willen, and Patricia A. Schiml. 2020. "Psychological Stress, Its Reduction, and Long-Term Consequences: What Studies with Laboratory Animals Might Teach Us about Life in the Dog Shelter" Animals 10, no. 11: 2061. https://doi.org/10.3390/ani10112061
APA StyleHennessy, M. B., Willen, R. M., & Schiml, P. A. (2020). Psychological Stress, Its Reduction, and Long-Term Consequences: What Studies with Laboratory Animals Might Teach Us about Life in the Dog Shelter. Animals, 10(11), 2061. https://doi.org/10.3390/ani10112061