
Basidiomycetous Yeast, Glaciozyma antarctica, Forming Frost-Columnar Colonies on Frozen Medium

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Fungal Isolation on Frozen Media
2.2. Phylogenic Analysis
2.3. Morphological Analyses on Frost-Columnar Colonies
2.4. Chemical Analyses of EPS
2.5. IBP Activity in Frost-Columnar Colonies and Culture Broth
2.6. Statistical Analysis
3. Results and Discussion
3.1. Fungal Growth on Frozen Media
3.2. The Structure of Frozen Colonies
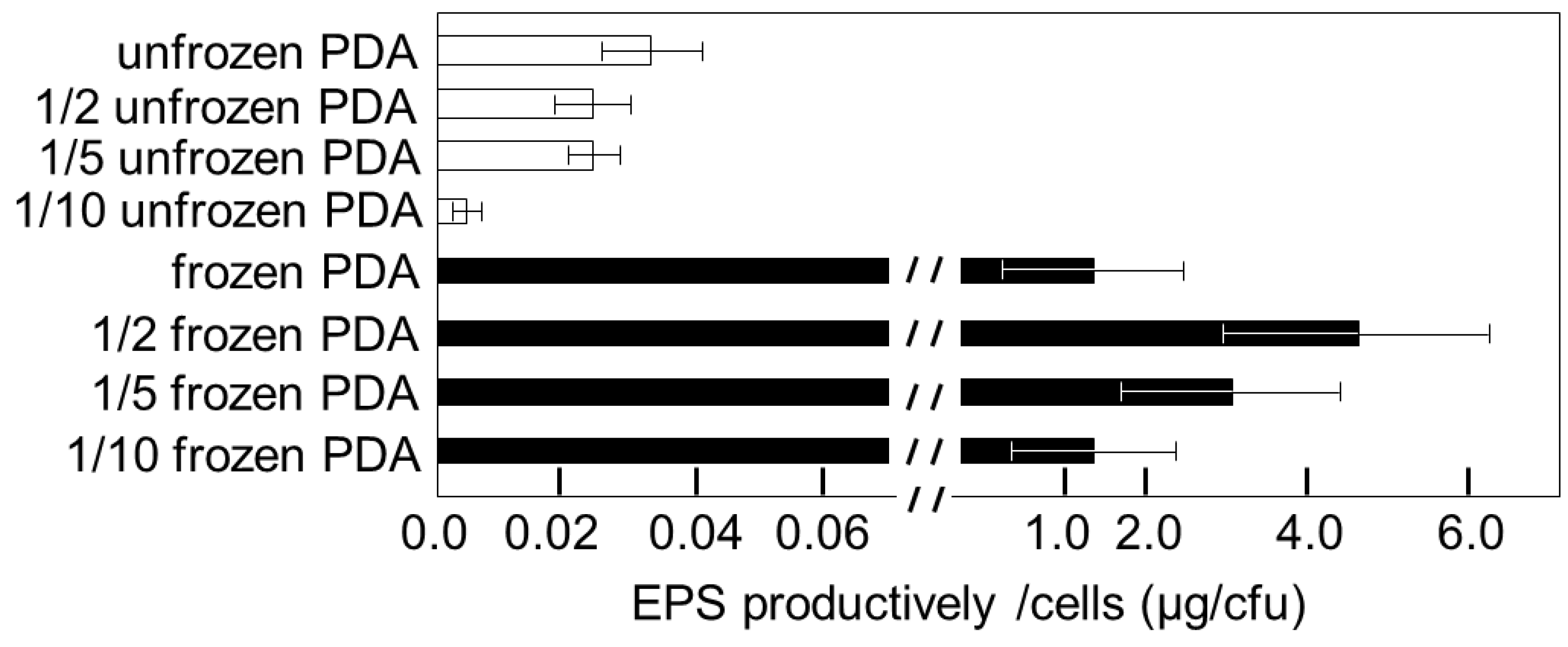
3.3. Cell Growth and EPS Production on Frozen Media
3.4. IBP Activity in Frozen Media
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Onofri, S.; Zucconi, L.; Tosi, S. Continental Antarctic Fungi; IHW Verlag: München, Germany, 2007. [Google Scholar]
- Fell, J.W.; Scorzetti, G.; Connell, L.; Craig, S. Biodiversity of micro-eukaryotes in Antarctic Dry Valley soils with <5% soil moisture. Soil Biol. Biochem. 2006, 38, 3107–3119. [Google Scholar]
- Tsuji, M. An index of fungi recorded from the vicinity of Syowa Station. Mycoscience 2018, 59, 319–324. [Google Scholar] [CrossRef]
- Tsuji, M. Genetic diversity of yeasts from East Ongul Island, East Antarctica and their extracellular enzymes secretion. Polar Biol. 2017, 41, 249–258. [Google Scholar] [CrossRef]
- Tsuji, M. An Index of Non-Lichenized Fungi Recorded in the Vicinity of Syowa Station, East Antarctica. In Fungi in Polar Regions; Informa UK Limited: London, UK, 2019; pp. 1–16. [Google Scholar]
- Tsuji, M.; Kudoh, S. Soil Yeasts in the Vicinity of Syowa Station, East Antarctica: Their diversity and extracellular enzymes, cold adaptation strategies, and secondary metabolites. Sustainability 2020, 12, 4518. [Google Scholar] [CrossRef]
- Canini, F.; Geml, J.; D’Acqui, L.; Buzzini, P.; Turchetti, B.; Onofri, S.; Ventura, S.; Zucconi, L. Fungal diversity and functionality are driven by soil texture in Taylor Valley, Antarctica. Fungal Ecol. 2021, 50, 101041. [Google Scholar] [CrossRef]
- Canini, F.; Geml, J.; Buzzini, P.; Turchetti, B.; Onofri, S.; D’Accqui, L.P.; Ripa, C.; Zucconi, L. Growth forms and functional guilds distribution of soil fungi in coastal versus lnland sites of Victoria Land, Anatrctica. Biology 2021, 10, 320. [Google Scholar] [CrossRef]
- de Menezes, G.C.A.; Câmara, P.E.A.S.; Pinto, O.H.B.; Carvalho-Siva, M.; Oliveira, F.S.; Souza, C.D.; Schaefer, C.E.G.R.; Convery, P.; Rosa, C.A.; Rosa, L.H. Fungal diversity present on rocks from a polar desert in continental Antarctica assessed using DNA metabardoding. Extremophiles 2021, 25, 193–202. [Google Scholar] [CrossRef]
- Hoshino, T.; Xiao, N.; Yajima, Y.; Kida, K.; Tokura, K.; Murakami, R.; Tojo, M.; Matsumoto, N. Ecological Strategies of Snow Molds to Tolerate Freezing Stress. In Plant and Microbe Adaptations to Cold in a Changing World; Springer: Berlin/Heidelberg, Germany, 2013; pp. 285–292. [Google Scholar]
- Hoshino, T.; Kiriaki, M.; Ohgiya, S.; Fujiwara, M.; Kondo, H.; Nishimiya, Y.; Yumoto, I.; Tsuda, S. Antifreeze proteins from snow mold fungi. Can. J. Bot. 2003, 81, 1175–1181. [Google Scholar] [CrossRef]
- Amato, P.; Doyle, S.; Christner, B.C. Macromolecular synthesis by yeasts under frozen conditions. Environ. Microbiol. 2009, 11, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Panikov, N.S. Subsero activity of cold-adapted yeasts. In Cold-Adapted Yeast; Buzzini, P., Margesin, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 294–323. [Google Scholar]
- Buzzini, P.; Turk, M.; Perini, L.; Turchetti, B.; Gunde-Cimerman, N. Yeasts in Polar and Subpolar Habitats. In Yeasts in Natural Ecosystems: Diversity; Buzzini, P., Lachance, M.A., Yurkov, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 331–365. [Google Scholar]
- Sannino, C.; Tasselli, G.; Filippucci, S.; Turchetti, B.; Buzzini, P. Yeasts in Nonpolar Cold Habitats. In Yeasts in Natural Ecosystems: Diversity; Buzzini, P., Lachance, M.A., Yurkov, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 367–396. [Google Scholar]
- Deegenaars, M.L.; Watson, K. Heat shock response in psychrophilic and psychrotrophic yeast from Antarctica. Extremophiles 1998, 2, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Maggi, O.; Tosi, S.; Angelova, M.; Lagostina, E.; Fabbri, A.A.; Pecoraro, L.; Altobelli, E.; Picco, A.M.; Savino, E.; Branda, E.; et al. Adaptation of fungi, including yeasts, to cold environments. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2013, 147, 247–258. [Google Scholar] [CrossRef]
- Tsuji, M. Cold-stress responses in the Antarctic basidiomycetous yeast Mrakia blollopis. R. Soc. Open Sci. 2016, 3, 160106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Jiang, X.; Wu, W.; Wang, M.; Hamid, M.I.; Xiang, M.; Liu, X. Genomic transcriptomic, and proteomic analysis provide insights into the cold adaptation mechanism of obligate psychrophilic fungus Mrakia psychrohila. G3 2016, 6, 3603–3613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, N.; Inaba, S.; Tojo, M.; Degawa, Y.; Fujiu, S.; Hanada, Y.; Kudoh, S.; Hoshino, T. Antifreeze activities of various fungi and Stramenopila isolated from Antactica. N. Am. Fungi 2010, 5, 215–220. [Google Scholar]
- Hashim, N.H.F.; Bharudin, I.; Nguong, D.L.S.; Higa, S.; Abu Bakar, F.D.; Nathan, S.; Rabu, A.; Kawahara, H.; Illias, R.M.; Najimudin, N.; et al. Characterization of Afp1, an antifreeze protein from the psychrophilic yeast Glaciozyma antarctica PI12. Extremophiles 2013, 17, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Hashim, N.H.F.; Sulaiman, S.; Baker, F.D.A.; Illias, R.M.; Kawahara, H.; Najimudin, N.; Mahadi, N.M.; Murad, A.M.A. Mo-lecular cloning, expression and characterization of Afp4, an antifreeze protein from Glacozyma antarctica. Polar Biol. 2014, 37, 1495–1505. [Google Scholar] [CrossRef]
- Lee, J.K.; Park, A.K.; Do, H.; Park, K.S.; Moh, A.H.; Kang, S.-H.; Kim, H.J. An extracellular ice-binding glycoprotein from an Arctic yeast. Cryobiology 2010, 50, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, P.; Carrasco, M.; Barahona, S.; Alcaíno, J.; Cifuentes, V.; Baeza, M. Antarctic yeasts: Analysis of their freeze-thaw tolerance and production of antifreeze proteins, fatty acids and ergosterol. BMC Microbiol. 2018, 18, 1–10. [Google Scholar] [CrossRef]
- Singh, P.; Singh, S.M.; Tsuji, M.; Prasad, G.S.; Hoshino, T. Rhodotorula svalbardensis sp. nov., a novel yeast species isolated from cryoconite holes of Ny-Ålesund, Arctic. Cryobiology 2014, 68, 122–128. [Google Scholar] [CrossRef]
- Tsuji, M.; Kudoh, S.; Hoshino, T.; Isojärvi, J.; Shunmugam, S.; Sivonen, K.; Allahverdiyeva, Y.; Aro, E.-M.; Battchikova, N. Draft Genome Sequence of Cryophilic Basidiomycetous Yeast Mrakia blollopis SK-4, Isolated from an Algal Mat of Naga-ike Lake in the Skarvsnes Ice-Free Area, East Antarctica. Genome Announc. 2015, 3, e01454-14. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, T.; Terami, F.; Tkachenko, O.B.; Tojo, M.; Matsumoto, N. Mycelial growth of the snow mold fungus, Sclerotinia borealis, improved at low water potentials: An adaption to frozen environment. Mycoscience 2010, 51, 98–103. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycor-rhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Abe, J.; Moriyama, M.; Kuwabara, T. Carbohydrate levels among winter wheat cultivars varying in freezing tolerance and snow mold resistance during autumn and winter. Physiol. Plant. 1998, 103, 8–16. [Google Scholar] [CrossRef]
- Matsuyama, H.; Kamesaki, T.; Sasaki, R.; Minami, H.; Yumoto, I. Production of two types of exopolysaccharide by Novo-sphingobium rosa. J. Biosci. Bioeng. 2003, 95, 152–156. [Google Scholar] [CrossRef]
- Xiao, N.; Suzuki, K.; Nishimiya, Y.; Kondo, H.; Miura, A.; Tsuda, S.; Hoshino, T. Comparison of functional properties of two fungal antifreeze proteins from Antarctomyces psychrotrophicus and Typhula ishikariensis. FEBS J. 2010, 277, 394–403. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Hoshino, T.; Kiriaki, M.; Nakajima, T. Novel thermal hysteresis proteins from low temperature basidiomycete, Coprinus psychromorbidus. Cryo Lett. 2003, 24, 135–142. [Google Scholar]
- Takamichi, M.; Nishimiya, Y.; Miura, A.; Tsuda, S. Effect of annealing time of an ice crystal on the activity of type III antifreeze protein. FEBS J. 2007, 274, 6469–6476. [Google Scholar] [CrossRef]
- Kachalkin, A.V. Yeasts of the White Sea intertidal zone and description of Glaciozyma litrale sp. nov. Antonie Leeuwenhoek 2014, 105, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Fell, J.W.; Statzell, A.C.; Hunter, I.L.; Phaff, H.J. Leucosporidium gen. n., the heterobasidiomycetous stage of several yeasts of the genus Candida. Antonie Leeuwenhoek 1969, 35, 433–462. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, M.; Yokota, Y.; Kudoh, S.; Hoshino, T. Effects of nitrogen concentration and culturing temperature on lipase secretion and morphology of the Antarctic basidiomycetous yeast Mrakia blollopis. Int. J. Res. Eng. Sci. 2014, 2, 49–54. [Google Scholar]
- Firdaus-Raih, M.; Hashim, N.H.F.; Bharudin, I.; Bakar, M.F.A.; Huang, K.K.; Alias, H.; Lee, B.K.B.; Isa, M.N.M.; Mat-Sharani, S.; Sulaiman, S.; et al. The Glaciozyma antartctica genome reveals an array of system that provide sustained responses towards temperature variations in a persistently cold habitat. PLoS ONE 2018, 13, e0189947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krembs, C.; Eicken, H.; Junge, K.; Deming, J.W. High concentrations of exopolymeric substances in Arctic winter sea ice: Implications for the polar ocean carbon cycle and cryoprotection of diatoms. Deep Sea Res. Part I 2002, 49, 2162–2181. [Google Scholar] [CrossRef]
- Kondo, H.; Hanada, Y.; Sugimoto, H.; Hoshino, T.; Garnham, C.P.; Davies, P.L.; Tsuda, S. Ice-binding site of snow mold fungus antifreeze protein deviates from structural regularity and high conservation. Proc. Natl. Acad. Sci. USA 2012, 109, 9360–9365. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | No. Samples Examined | No. Samples with Fungal Growth | Samples Producing Frost-Columnar Colonies |
|---|---|---|---|
| Sôya Coast Ongule Islands | |||
| East Ongul Is. | 54 | 36 | 8 |
| West Ongul Is. | 13 | 9 | 0 |
| Ongul Kalven Is. | 7 | 0 | 0 |
| Langhovde | 40 | 26 | 6 |
| Skarvsnes | 125 | 71 | 15 |
| Skallen | 11 | 7 | 1 |
| Rundvågshetta | 14 | 12 | 6 |
| Prince Harald Coast | |||
| Riiser-Larsen Peninsula | 13 | 5 | 1 |
| Total | 277 | 166 | 37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujiu, S.; Ito, M.; Kobayashi, E.; Hanada, Y.; Yoshida, M.; Kudoh, S.; Hoshino, T. Basidiomycetous Yeast, Glaciozyma antarctica, Forming Frost-Columnar Colonies on Frozen Medium. Microorganisms 2021, 9, 1679. https://doi.org/10.3390/microorganisms9081679
Fujiu S, Ito M, Kobayashi E, Hanada Y, Yoshida M, Kudoh S, Hoshino T. Basidiomycetous Yeast, Glaciozyma antarctica, Forming Frost-Columnar Colonies on Frozen Medium. Microorganisms. 2021; 9(8):1679. https://doi.org/10.3390/microorganisms9081679
Chicago/Turabian StyleFujiu, Seiichi, Masanobu Ito, Eriko Kobayashi, Yuichi Hanada, Midori Yoshida, Sakae Kudoh, and Tamotsu Hoshino. 2021. "Basidiomycetous Yeast, Glaciozyma antarctica, Forming Frost-Columnar Colonies on Frozen Medium" Microorganisms 9, no. 8: 1679. https://doi.org/10.3390/microorganisms9081679
APA StyleFujiu, S., Ito, M., Kobayashi, E., Hanada, Y., Yoshida, M., Kudoh, S., & Hoshino, T. (2021). Basidiomycetous Yeast, Glaciozyma antarctica, Forming Frost-Columnar Colonies on Frozen Medium. Microorganisms, 9(8), 1679. https://doi.org/10.3390/microorganisms9081679