
Leuconostoc mesenteroides Strains Isolated from Carrots Show Probiotic Features

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Isolation and Identification
2.2. Growth Conditions of Bacterial Isolates
2.3. Resistance to Lysozyme, Acid pH and Bile Salts
2.4. Antibiotic Resistance
2.5. Antimicrobial Activity
2.6. C. elegans Strain and Lifespan Assay
2.7. Fertility Assay
2.8. Colonization Assay of C. elegans Gut
2.9. Aging Markers’ Analysis
2.10. Resistance to Pathogens in C. elegans
2.11. Real-Time qPCR
2.12. Statistical Analysis
3. Results
3.1. Bacteria Isolation
3.2. In Vitro Tests
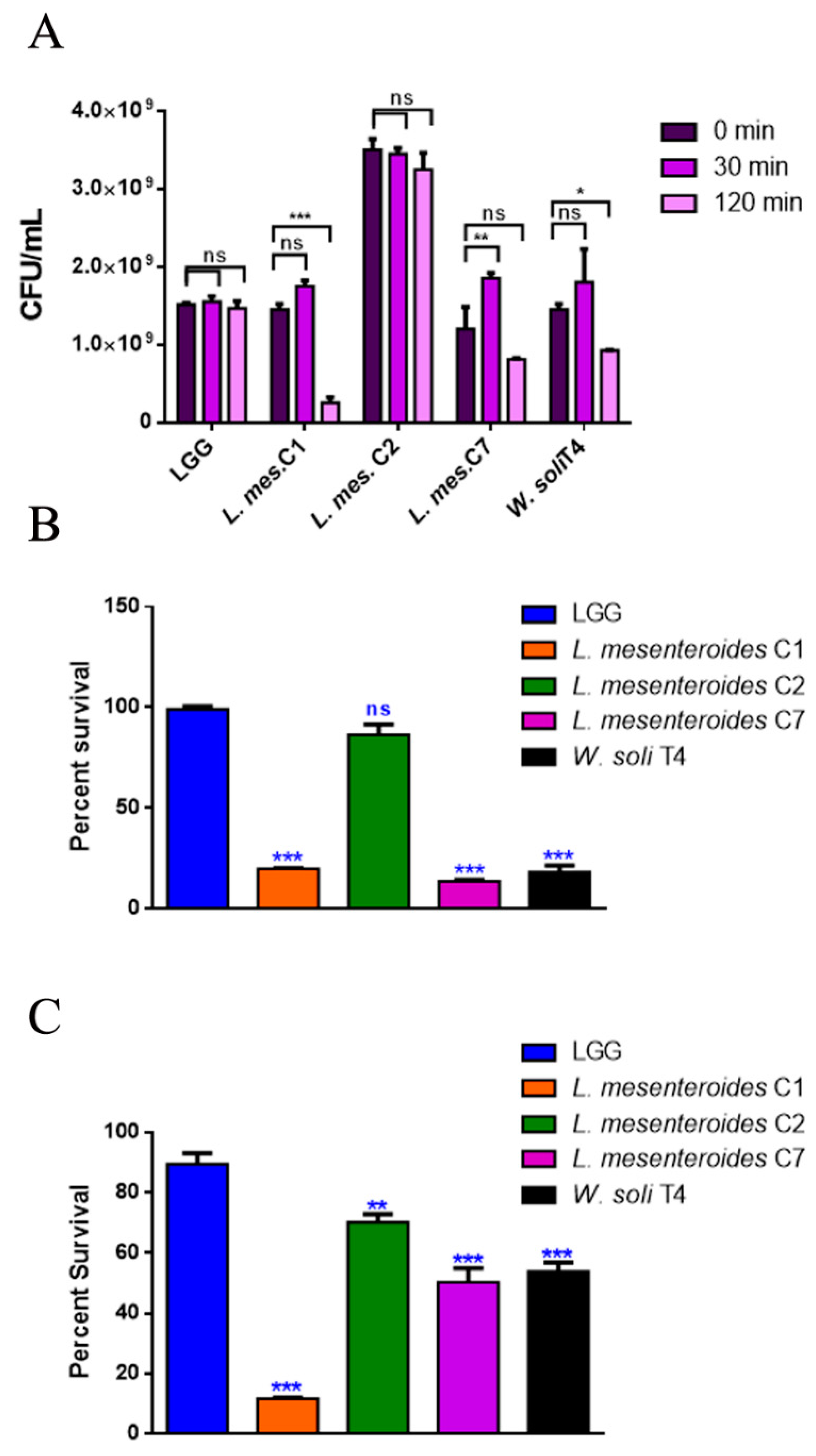
3.2.1. Resistance to Lysozyme, Low pH and Bile Salts
3.2.2. Antibiotic Susceptibility and Antagonistic Activity to Pathogens
3.3. In Vivo Tests
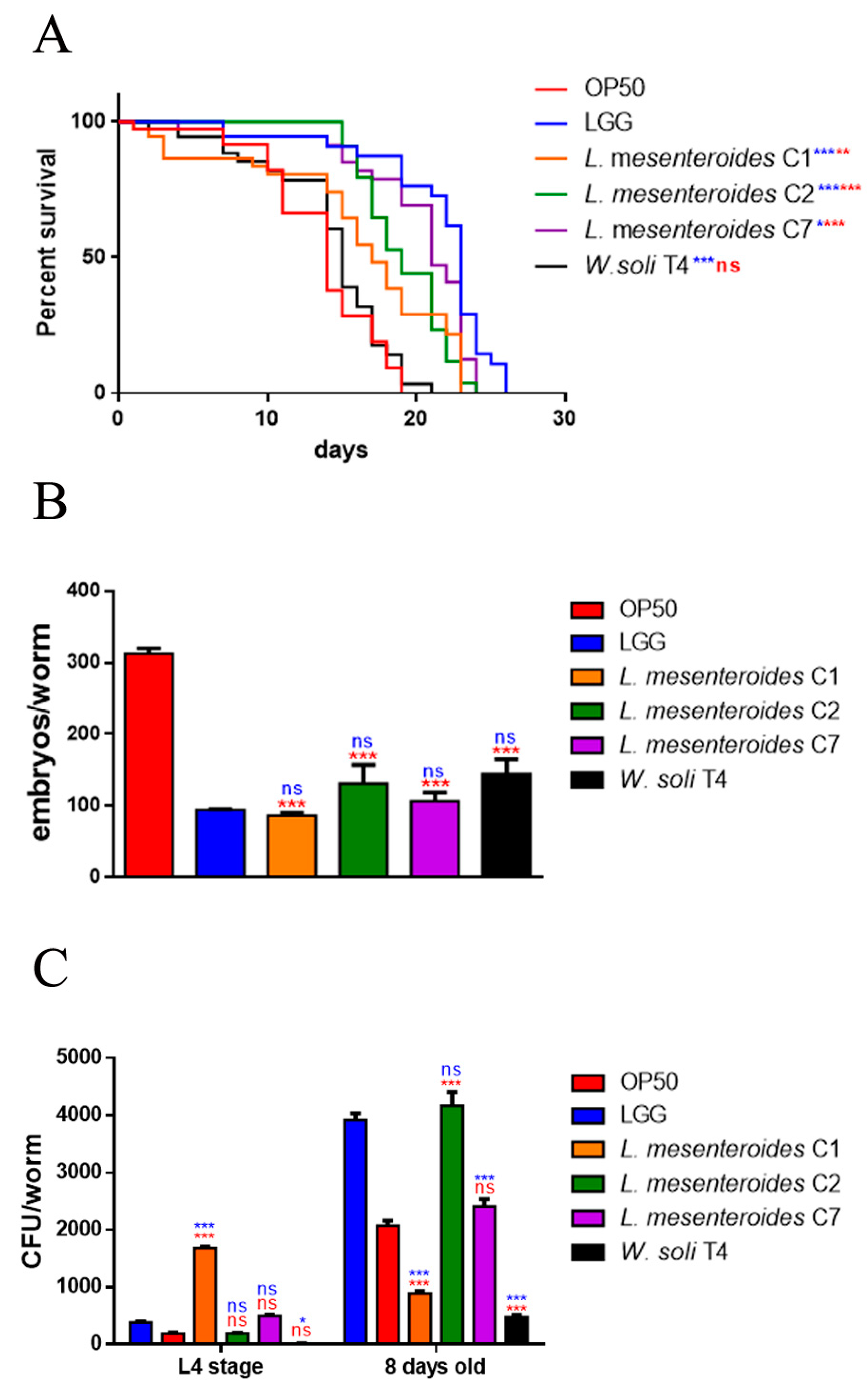
3.3.1. Effects on C. elegans Lifespan and Colonization Capability
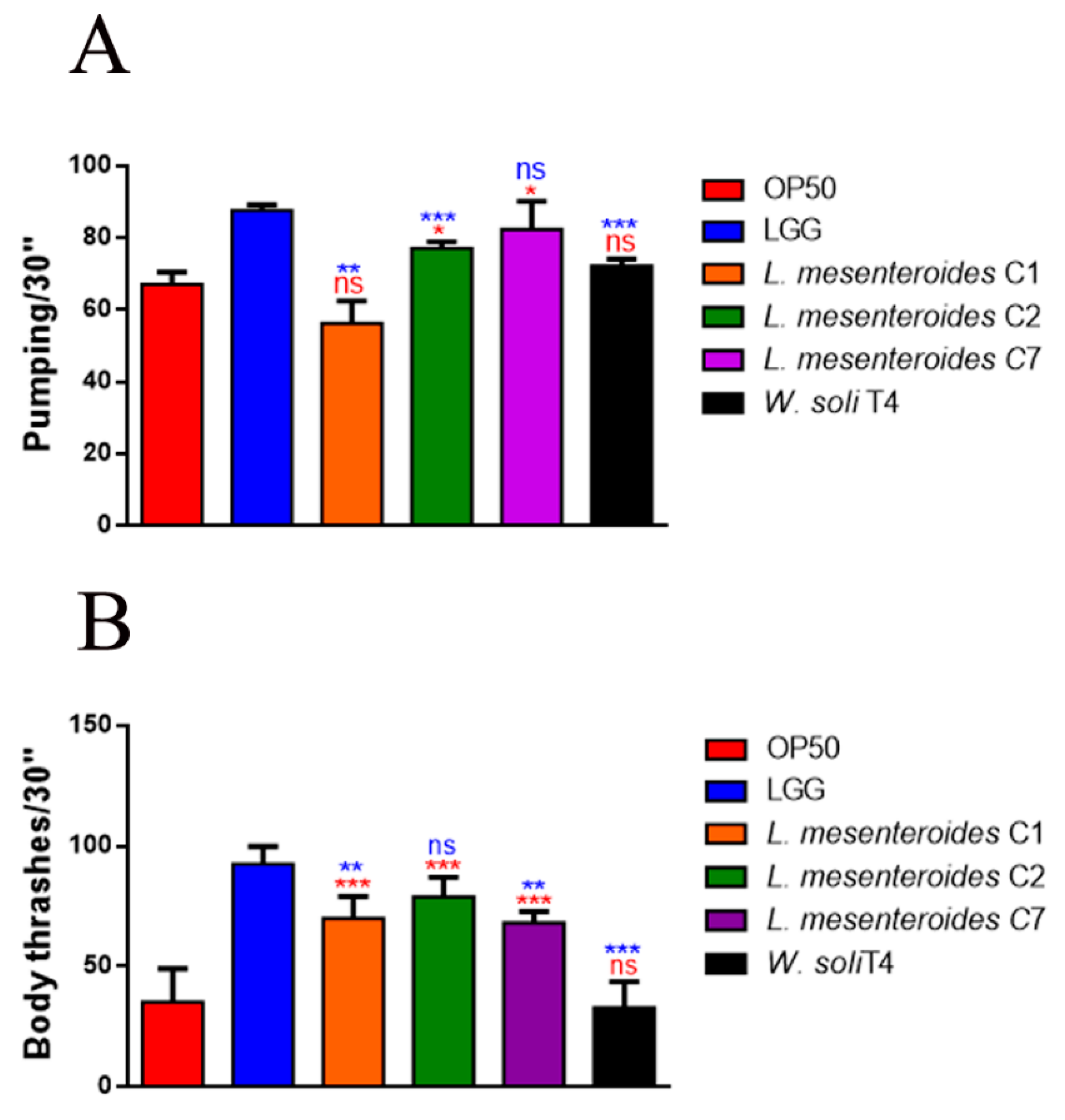
3.3.2. Effects on C. elegans Aging Processes
3.3.3. Pathogen Resistance and Innate Immunity Stimulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO/WHO Probiotics in Food. Health and Nutritional Properties and Guidelines for Evaluation 2006. Available online: http://www.fao.org/food/food-safety-quality/a-z-index/probiotics/en/ (accessed on 16 September 2021).
- Markowiak-Kopeć, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Roselli, M.; Natella, F.; Zinno, P.; Guantario, B.; Canali, R.; Schifano, E.; De Angelis, M.; Nikoloudaki, O.; Gobbetti, M.; Perozzi, G.; et al. Colonization Ability and Impact on Human Gut Microbiota of Foodborne Microbes from Traditional or Probiotic-Added Fermented Foods: A Systematic Review. Front. Nutr. 2021, 8, 689084. [Google Scholar] [CrossRef] [PubMed]
- Sciubba, F.; Tomassini, A.; Giorgi, G.; Brasili, E.; Pasqua, G.; Capuani, G.; Aureli, W.; Miccheli, A. NMR-Based Metabolomic Study of Purple Carrot Optimal Harvest Time for Utilization as a Source of Bioactive Compounds. Appl. Sci. 2020, 10, 8493. [Google Scholar] [CrossRef]
- Tomassini, A.; Sciubba, F.; Di Cocco, M.E.; Capuani, G.; Delfini, M.; Aureli, W.; Miccheli, A. 1H NMR-Based Metabolomics Reveals a Pedoclimatic Metabolic Imprinting in Ready-to-Drink Carrot Juices. J. Agric. Food Chem. 2016, 64, 5284–5291. [Google Scholar] [CrossRef] [PubMed]
- Arscott, S.A.; Tanumihardjo, S.A. Carrots of Many Colors Provide Basic Nutrition and Bioavailable Phytochemicals Acting as a Functional Food. Compr. Rev. Food Sci. Food Saf. 2010, 9, 223–239. [Google Scholar] [CrossRef]
- Que, F.; Hou, X.-L.; Wang, G.-L.; Xu, Z.-S.; Tan, G.-F.; Li, T.; Wang, Y.-H.; Khadr, A.; Xiong, A.-S. Advances in research on the carrot, an important root vegetable in the Apiaceae family. Hortic. Res. 2019, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, E.A.; Vieira, M.A.; Vieira, E.A.; Amboni, R.D.D.M.C.; Amante, E.R.; Teixeira, E. Chemical, Physical and Sensory Parameters of Different Carrot Varieties (Daucus carota L.). J. Food Process. Eng. 2007, 30, 746–756. [Google Scholar] [CrossRef]
- Forssten, S.D.; Sindelar, C.W.; Ouwehand, A. Probiotics from an industrial perspective. Anaerobe 2011, 17, 410–413. [Google Scholar] [CrossRef]
- Sornplang, P.; Piyadeatsoontorn, S. Probiotic isolates from unconventional sources: A review. J. Anim. Sci. Technol. 2016, 58, 26. [Google Scholar] [CrossRef] [Green Version]
- Poupet, C.; Chassard, C.; Nivoliez, A.; Bornes, S. Caenorhabditis elegans, a Host to Investigate the Probiotic Properties of Beneficial Microorganisms. Front. Nutr. 2020, 7, 135. [Google Scholar] [CrossRef]
- Kwon, G.; Lee, J.; Lim, Y.-H. Dairy Propionibacterium extends the mean lifespan of Caenorhabditis elegans via activation of the innate immune system. Sci. Rep. 2016, 6, 31713. [Google Scholar] [CrossRef] [PubMed]
- Schifano, E.; Zinno, P.; Guantario, B.; Roselli, M.; Marcoccia, S.; Devirgiliis, C.; Uccelletti, D. The Foodborne Strain Lactobacillus fermentum MBC2 Triggers pept-1-Dependent Pro-Longevity Effects in Caenorhabditis elegans. Microorganisms 2019, 7, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guantario, B.; Zinno, P.; Schifano, E.; Roselli, M.; Perozzi, G.; Palleschi, C.; Uccelletti, D.; Devirgiliis, C. In Vitro and In Vivo Selection of Potentially Probiotic Lactobacilli from Nocellara del Belice Table Olives. Front. Microbiol. 2018, 9, 595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roselli, M.; Schifano, E.; Guantario, B.; Zinno, P.; Uccelletti, D.; Devirgiliis, C. Caenorhabditis Elegans and Probiotics Interactions from a Prolongevity Perspective. Int. J. Mol. Sci. 2019, 20, 5020. [Google Scholar] [CrossRef] [Green Version]
- Schifano, E.; Cavallini, D.; De Bellis, G.; Bracciale, M.P.; Felici, A.C.; Santarelli, M.L.; Sarto, M.S.; Uccelletti, D. Antibacterial Effect of Zinc Oxide-Based Nanomaterials on Environmental Biodeteriogens Affecting Historical Buildings. Nanomaterials 2020, 10, 335. [Google Scholar] [CrossRef] [Green Version]
- Samedi, L.; Charles, A.L. Isolation and characterization of potential probiotic Lactobacilli from leaves of food plants for possible additives in pellet feeding. Ann. Agric. Sci. 2019, 64, 55–62. [Google Scholar] [CrossRef]
- Erkkilä, S.; Petäjä, E. Screening of commercial meat starter cultures at low pH and in the presence of bile salts for potential probiotic use. Meat Sci. 2000, 55, 297–300. [Google Scholar] [CrossRef]
- Zanni, E.; Laudenzi, C.; Schifano, E.; Palleschi, C.; Perozzi, G.; Uccelletti, D.; Devirgiliis, C. Impact of a Complex Food Microbiota on Energy Metabolism in the Model Organism Caenorhabditis elegans. BioMed Res. Int. 2015, 2015, 621709. [Google Scholar] [CrossRef] [Green Version]
- Zanni, E.; Schifano, E.; Motta, S.; Sciubba, F.; Palleschi, C.; Mauri, P.; Perozzi, G.; Uccelletti, D.; Devirgiliis, C.; Miccheli, A. Combination of Metabolomic and Proteomic Analysis Revealed Different Features among Lactobacillus delbrueckii Subspecies bulgaricus and lactis Strains While In Vivo Testing in the Model Organism Caenorhabditis elegans Highlighted Probiotic Properties. Front. Microbiol. 2017, 8, 1206. [Google Scholar] [CrossRef] [Green Version]
- Schifano, E.; Marazzato, M.; Ammendolia, M.G.; Zanni, E.; Ricci, M.; Comanducci, A.; Goldoni, P.; Conte, M.P.; Uccelletti, D.; Longhi, C. Virulence behavior of uropathogenic Escherichia colistrains in the host model Caenorhabditis elegans. Microbiologyopen 2019, 8, e00756. [Google Scholar] [CrossRef]
- Schifano, E.; Ficociello, G.; Vespa, S.; Ghosh, S.; Cipollo, J.F.; Talora, C.; Lotti, L.V.; Mancini, P.; Uccelletti, D. Pmr-1 gene affects susceptibility of Caenorhabditis elegans to Staphylococcus aureus infection through glycosylation and stress response pathways’ alterations. Virulence 2019, 10, 1013–1025. [Google Scholar] [CrossRef] [Green Version]
- Schifano, E.; Cicalini, I.; Pieragostino, D.; Heipieper, H.J.; Del Boccio, P.; Uccelletti, D. In vitro and in vivo lipidomics as a tool for probiotics evaluation. Appl. Microbiol. Biotechnol. 2020, 104, 8937–8948. [Google Scholar] [CrossRef]
- Dunne, C.; O’Mahony, L.; Murphy, L.; Thornton, G.; Morrissey, D.; O’Halloran, S.; Feeney, M.; Flynn, S.; Fitzgerald, G.; Daly, C.; et al. In vitro selection criteria for probiotic bacteria of human origin: Correlation with in vivo findings. Am. J. Clin. Nutr. 2001, 73, 386s–392s. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, L.; Laghi, L.; Correani, V.; Schifano, E.; Landi, C.; Uccelletti, D.; Mattei, B. A Combined Proteomics, Metabolomics and In Vivo Analysis Approach for the Characterization of Probiotics in Large-Scale Production. Biomolecules 2020, 10, 157. [Google Scholar] [CrossRef] [Green Version]
- Terpou, A.; Papadaki, A.; Lappa, I.K.; Kachrimanidou, V.; Bosnea, L.A.; Kopsahelis, N. Probiotics in Food Systems: Significance and Emerging Strategies Towards Improved Viability and Delivery of Enhanced Beneficial Value. Nutrients 2019, 11, 1591. [Google Scholar] [CrossRef] [Green Version]
- Domingos-Lopes, M.; Stanton, C.; Ross, P.; Dapkevicius, M.; Silva, C. Genetic diversity, safety and technological characterization of lactic acid bacteria isolated from artisanal Pico cheese. Food Microbiol. 2017, 63, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Mokoena, M.P.; Mutanda, T.; Olaniran, A.O. Perspectives on the probiotic potential of lactic acid bacteria from African traditional fermented foods and beverages. Food Nutr. Res. 2016, 60, 29630. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, L.G.R.; Gasga, V.M.Z.; Pescuma, M.; Van Nieuwenhove, C.; Mozzi, F.; Burgos, J.A.S. Fruits and fruit by-products as sources of bioactive compounds. Benefits and trends of lactic acid fermentation in the development of novel fruit-based functional beverages. Food Res. Int. 2021, 140, 109854. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; Filannino, P.; Gobbetti, M. Vegetable and Fruit Fermentation by Lactic Acid Bacteria. Biotechnol. Lact. Acid Bact. 2015, 216–230. [Google Scholar] [CrossRef]
- Ngea, G.L.N.; Yang, Q.; Tchabo, W.; Castoria, R.; Zhang, X.; Zhang, H. Leuconostoc mesenteroides subsp. mesenteroides LB7 isolated from apple surface inhibits P. expansum in vitro and reduces patulin in fruit juices. Int. J. Food Microbiol. 2021, 339, 109025. [Google Scholar] [CrossRef]
- O’Sullivan, L.; Ross, R.; Hill, C. Potential of bacteriocin-producing lactic acid bacteria for improvements in food safety and quality. Biochimie 2002, 84, 593–604. [Google Scholar] [CrossRef]
- Dicks, L.M.T.; Botes, M. Probiotic lactic acid bacteria in the gastro-intestinal tract: Health benefits, safety and mode of action. Benef. Microbes 2010, 1, 11–29. [Google Scholar] [CrossRef]
- Imperial, I.C.V.; Ibana, J.A. Addressing the Antibiotic Resistance Problem with Probiotics: Reducing the Risk of Its Double-Edged Sword Effect. Front. Microbiol. 2016, 7, 1983. [Google Scholar] [CrossRef] [PubMed]
- Rafieian-Kopaei, M.; Karami, S.; Roayaei, M.; Hamzavi, H.; Bahmani, M.; Hassanzad-Azar, H.; Leila, M. Isolation and identification of probiotic Lactobacillus from local dairy and evaluating their antagonistic effect on pathogens. Int. J. Pharm. Investig. 2017, 7, 137–141. [Google Scholar] [CrossRef]
- Mathipa, M.G.; Thantsha, M.S. Probiotic engineering: Towards development of robust probiotic strains with enhanced functional properties and for targeted control of enteric pathogens. Gut Pathog. 2017, 9, 28. [Google Scholar] [CrossRef]
- Pawlowska, A.M.; Zannini, E.; Coffey, A.; Arendt, E.K. “Green Preservatives”: Combating Fungi in the Food and Feed Industry by Applying Antifungal Lactic Acid Bacteria. Nutr. Benefits Kiwifruit 2012, 66, 217–238. [Google Scholar] [CrossRef]
- Peyer, L.C.; Zannini, E.; Arendt, E.K. Lactic acid bacteria as sensory biomodulators for fermented cereal-based beverages. Trends Food Sci. Technol. 2016, 54, 17–25. [Google Scholar] [CrossRef]
- Sadiq, F.A.; Yan, B.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Lactic Acid Bacteria as Antifungal and Anti-Mycotoxigenic Agents: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1403–1436. [Google Scholar] [CrossRef] [Green Version]
- Silva, H.; Balthazar, C.F.; Silva, R.; Vieira, A.; Costa, R.; Esmerino, E.; Freitas, M.; Cruz, A. Sodium reduction and flavor enhancer addition in probiotic prato cheese: Contributions of quantitative descriptive analysis and temporal dominance of sensations for sensory profiling. J. Dairy Sci. 2018, 101, 8837–8846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, H.; Shiozaki, T.; Kobatake, E.; Hosoya, T.; Moriya, T.; Sakai, F.; Taru, H.; Miyazaki, T. Effects and mechanisms of prolongevity induced by Lactobacillus gasseri SBT2055 in Caenorhabditis elegans. Aging Cell 2015, 15, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, T.; Yasui, C.; Hoshino, K.; Arikawa, K.; Nishikawa, Y. Influence of Lactic Acid Bacteria on Longevity of Caenorhabditis elegans and Host Defense against Salmonella enterica Serovar Enteritidis. Appl. Environ. Microbiol. 2007, 73, 6404–6409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komura, T.; Ikeda, T.; Yasui, C.; Saeki, S.; Nishikawa, Y. Mechanism underlying prolongevity induced by bifidobacteria in Caenorhabditis elegans. Biogerontology 2013, 14, 73–87. [Google Scholar] [CrossRef] [Green Version]
- Oh, A.; Daliri, E.B.-M.; Oh, D.H. Screening for potential probiotic bacteria from Korean fermented soybean paste: In vitro and Caenorhabditis elegans model testing. LWT 2018, 88, 132–138. [Google Scholar] [CrossRef]
- Clark, L.C.; Hodgkin, J. Commensals, probiotics and pathogens in the Caenorhabditis elegansmodel. Cell. Microbiol. 2013, 16, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Irazoqui, J.E.; Troemel, E.R.; Feinbaum, R.L.; Luhachack, L.G.; Cezairliyan, B.O.; Ausubel, F.M. Distinct Pathogenesis and Host Responses during Infection of C. elegans by P. aeruginosa and S. aureus. PLoS Pathog. 2010, 6, e1000982. [Google Scholar] [CrossRef] [Green Version]
- Marsh, E.; May, R.C. Caenorhabditis elegans, a Model Organism for Investigating Immunity. Appl. Environ. Microbiol. 2012, 78, 2075–2081. [Google Scholar] [CrossRef] [Green Version]
- Kwon, G.; Lee, J.; Koh, J.-H.; Lim, Y.-H. Lifespan Extension of Caenorhabditis elegans by Butyricicoccus pullicaecorum and Megasphaera elsdenii with Probiotic Potential. Curr. Microbiol. 2017, 75, 557–564. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| hsf-1 | FOR | 5′-ATGACTCCACTGTCCCAAGG |
| REV | 5′-TCTTGCCGATTGCTTTCTCT | |
| pmk-1 | FOR | 5′-AAATGACTCGCCGTGATTTC |
| REV | 5′-CATCGTGATAAGCAGCCAGA | |
| sod-3 | FOR | 5′-AGAACCTTCAAAGGAGCTGATG |
| REV | 5′-CCGCAATAGTGATGTCAGAAAG | |
| act-1 | FOR | 5′-GAGCGTGGTTACTCTTTCA |
| REV | 5′-CAGAGCTTCTCCTTGATGTC | |
| skn-1 | FOR | 5′-GTTCCCAACATCCAACTACG |
| REV | 5′-TGGAGTCTGACCAGTGGATT | |
| daf-16 | FOR | 5′-TCAAGACCTCAAAGCCAATCAACTC |
| REV | 5′-ACGAGAAAGAAGGAGTAAGAGGAGG |
| Antibiotic | Amount on Disc (µg) | Zone of Inhibition (mm) | |||||
|---|---|---|---|---|---|---|---|
| LGG | L. mesenteroides C1 | L. mesenteroides C2 | L. mesenteroides C7 | W. soli T4 | p-Value | ||
| Amikacin | 30 | 4 ± 0.03 | 5 ± 0.06 | 10 ± 0.08 | 4 ± 0.05 | 4 ± 0.03 | p < 0.001 |
| Ampicillin | 10 | 7 ± 0.06 | 7 ± 0.05 | 6 ± 0.10 | 5 ± 0.20 | 5 ± 0.05 | p < 0.01 |
| Aztreonam | 30 | + | + | + | + | + | ns |
| Carbenicillin | 100 | 8 ± 0.08 | 7 ± 0.03 | 5 ± 0.09 | 7 ± 0.03 | 5 ± 0.08 | p < 0.01 |
| Cefalotin | 30 | + | 3 ± 0.10 | + | 4 ± 0.08 | + | p < 0.01 |
| Cefotaxime | 30 | 7 ± 0.08 | + | 7 ± 0.20 | + | + | p < 0.001 |
| Cefuroxime | 30 | 5 ± 0.02 | + | 12 ± 0.08 | + | + | p < 0.001 |
| Clindamycin | 2 | 8 ± 0.12 | 9 ± 0.35 | 4 ± 0.08 | 10 ± 0.10 | 9 ± 0.30 | p < 0.01 |
| Chloramphenicol | 30 | 8 ± 0.08 | 8 ± 0.35 | 14 ± 0.10 | 8 ± 0.20 | 10 ± 0.15 | p < 0.01 |
| Erythromycin | 15 | 8 ± 0.03 | 7 ± 0.08 | 8 ± 0.15 | 7 ± 0.09 | 8 ± 0.08 | ns |
| Fosfomycin | 50 | + | + | + | + | + | ns |
| Gentamicin | 10 | 4 ± 0.05 | 5 ± 0.12 | + | 5 ± 0.15 | 5 ± 0.10 | p < 0.05 |
| Mezlocillin | 75 | 14 ± 0.05 | 10 ± 0.15 | 13 ± 0.20 | 8 ± 0.20 | 11 ± 0.10 | p < 0.001 |
| Oxacillin | 1 | + | + | 4 ± 0.03 | + | + | p < 0.05 |
| Penicillin | 10 | 13 ± 0.10 | 9 ± 0.20 | 14 ± 0.20 | 9 ± 0.05 | 11 ± 0.09 | p < 0.001 |
| Rinfampicin | 30 | 13 ± 0.02 | 11 ± 0.09 | 8 ± 0.10 | 11 ± 0.08 | 10 ± 0.08 | p < 0.01 |
| Streptomycin | 25 | 4 ± 0.12 | 5 ± 0.08 | + | 4 ± 0.06 | 4 ± 0.05 | p < 0.01 |
| Tetracycline | 30 | 14 ± 0.15 | 10 ± 0.30 | 7 ± 0.05 | 8 ± 0.06 | 7 ± 0.10 | p < 0.001 |
| Tobramycin | 10 | 4 ± 0.08 | 4 ± 0.05 | 4 ± 0.09 | 3 ± 0.05 | 4 ± 0.05 | ns |
| Vancomycin | 30 | + | + | + | + | + | ns |
| Pathogen | LGG | L. mesenteroides C1 | L. mesenteroides C2 | L. mesenteroides C7 | W. soli T4 | p-Value |
|---|---|---|---|---|---|---|
| S. aureus | 40 ± 0.08 | 38 ± 0.4 | 35 ± 0.5 | 35 ± 0.1 | 35 ± 0.5 | p < 0.01 |
| L. monocytogenes | 30 ± 0.2 | 29 ± 0.2 | 30 ± 0.8 | 31 ± 0.6 | 30 ± 0.5 | ns |
| P. aeruginosa | 40 ± 0.07 | 38 ± 0.5 | 33 ± 0.5 | 40 ± 0.1 | 35 ± 0.6 | p < 0.01 |
| S. enterica | 30 ± 0.3 | 31 ± 0.3 | 30 ± 0.5 | 28 ± 0.5 | 30 ± 0.08 | ns |
| C. elegans Strain | Diet | Median Lifespan | Maximum Lifespan | Statistics |
|---|---|---|---|---|
| Wild-type N2 | P. aeruginosa | 3 ± 0.8 | 5 ± 0.8 | - |
| LGG + P. aeruginosa | 5 ± 0.9 | 9 ± 0.4 | p < 0.001 | |
| L. mesenteroides C2 + P. aeruginosa | 6 ± 1.2 | 12 ± 0.8 | p < 0.001 | |
| L. mesenteroides C7 + P. aeruginosa | 6 ± 0.2 | 11 ± 0.3 | p < 0.001 | |
| S. aureus | 5 ± 0.4 | 8 ± 0.9 | - | |
| LGG + S. aureus | 8 ± 0.5 | 13 ± 0.5 | p < 0.001 | |
| L. mesenteroides C2 + S. aureus | 7 ± 0.6 | 11 ± 0.5 | p < 0.001 | |
| L. mesenteroides C7 + S. aureus | 7 ± 0.9 | 11 ± 1.2 | p < 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schifano, E.; Tomassini, A.; Preziosi, A.; Montes, J.; Aureli, W.; Mancini, P.; Miccheli, A.; Uccelletti, D. Leuconostoc mesenteroides Strains Isolated from Carrots Show Probiotic Features. Microorganisms 2021, 9, 2290. https://doi.org/10.3390/microorganisms9112290
Schifano E, Tomassini A, Preziosi A, Montes J, Aureli W, Mancini P, Miccheli A, Uccelletti D. Leuconostoc mesenteroides Strains Isolated from Carrots Show Probiotic Features. Microorganisms. 2021; 9(11):2290. https://doi.org/10.3390/microorganisms9112290
Chicago/Turabian StyleSchifano, Emily, Alberta Tomassini, Adele Preziosi, Jorge Montes, Walter Aureli, Patrizia Mancini, Alfredo Miccheli, and Daniela Uccelletti. 2021. "Leuconostoc mesenteroides Strains Isolated from Carrots Show Probiotic Features" Microorganisms 9, no. 11: 2290. https://doi.org/10.3390/microorganisms9112290
APA StyleSchifano, E., Tomassini, A., Preziosi, A., Montes, J., Aureli, W., Mancini, P., Miccheli, A., & Uccelletti, D. (2021). Leuconostoc mesenteroides Strains Isolated from Carrots Show Probiotic Features. Microorganisms, 9(11), 2290. https://doi.org/10.3390/microorganisms9112290