Environmental Conditions Affecting GABA Production in Lactococcus lactis NCDO 2118

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Organism and Growth Conditions
2.1.1. Cultures in Tubes
2.1.2. Cultures in Bioreactor
2.2. GABA Determination
2.3. Glutamate Decarboxylase (GAD) Activity
2.4. RNA Extraction and Gene Expression Analysis
2.5. Osmolarity Measurement
2.6. Growth Rate Estimations
2.7. Statistical Treatments
3. Results
3.1. Impact of Osmolarity on the Growth Rate
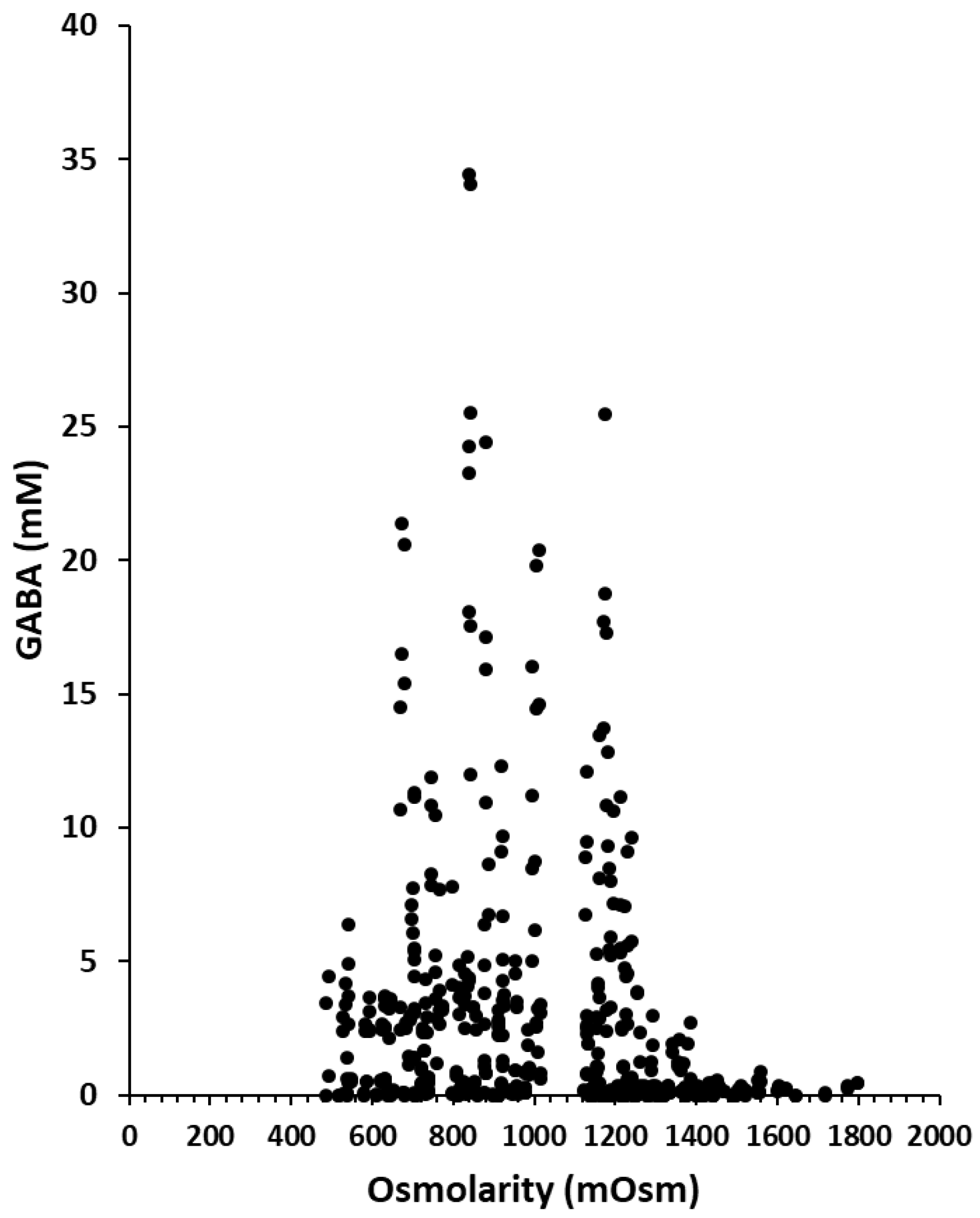
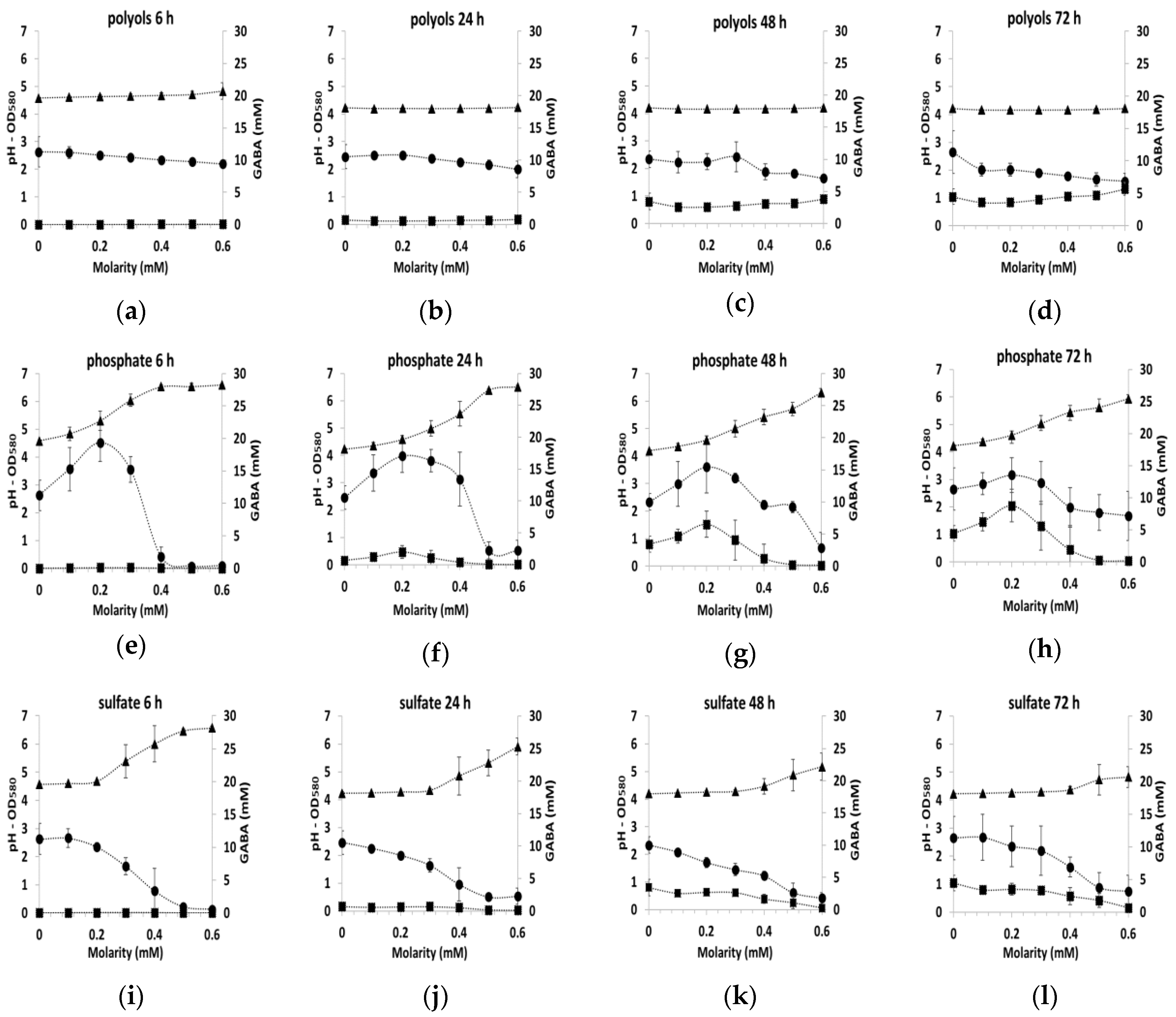
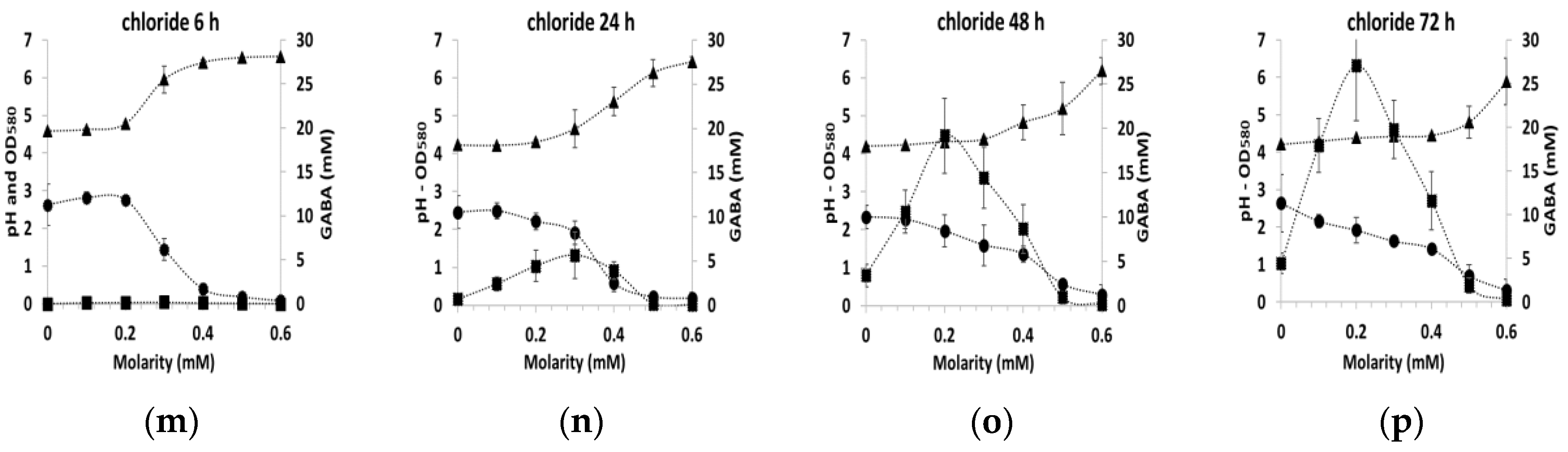
3.2. GABA Production under Osmotic Stress
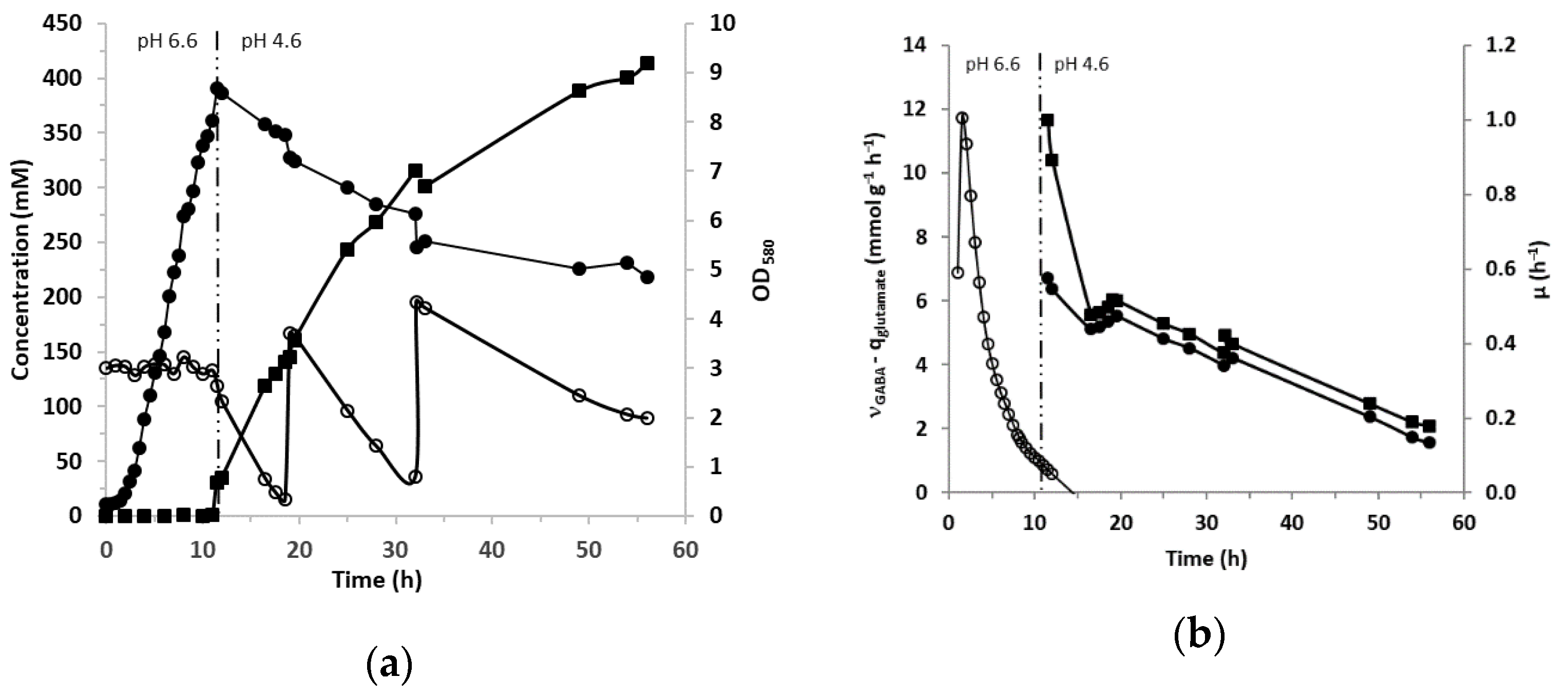
3.3. GABA Production in Bioreactor
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fondén, R.; Saarela, M.; Mättö, J.; Matilla-Sandholm, T. Lactic acid bacteria (LAB) in functional dairy products. In Functional Dairy Products; Elsevier: Amsterdam, The Netherlands, 2003; pp. 244–262. ISBN 978-1-85573-584-2. [Google Scholar]
- Hossain, I.; Sadekuzzaman, M.; Ha, S.-D. Probiotics as potential alternative biocontrol agents in the agriculture and food industries: A review. Food Res. Int. 2017, 100, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Pingitore, E.V.; Pessione, A.; Fontana, C.; Mazzoli, R.; Pessione, E. Comparative proteomic analyses for elucidating metabolic changes during EPS production under different fermentation temperatures by Lactobacillus plantarum Q823. Int. J. Food Microbiol. 2016, 238, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Pessione, E. Lactic acid bacteria contribution to gut microbiota complexity: Lights and shadows. Front. Cell. Infect. Microbiol. 2012, 2, 86. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Pingitore, E.V.; Mozzi, F.; Saavedra, L.; Villegas, J.M.; Hebert, E. Lactic Acid Bacteria as Cell Factories for the Generation of Bioactive Peptides. Protein Pept. Lett. 2017, 24, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Karthik, A. Nutraceuticals: Globals Markets to 2023; BCC Research: Boston, MA, USA, 2018. [Google Scholar]
- Mazzoli, R.; Pessione, E. The Neuro-endocrinological Role of Microbial Glutamate and GABA Signaling. Front. Microbiol. 2016, 7, 1934. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Shirai, T.; Ochiai, H.; Kasao, M.; Hayakawa, K.; Kimura, M.; Sansawa, H. Blood-pressure-lowering effect of a novel fermented milk containing γ-aminobutyric acid (GABA) in mild hypertensives. Eur. J. Clin. Nutr. 2003, 57, 490–495. [Google Scholar] [CrossRef]
- Oh, S.-H.; Oh, C.H. Brown rice extracts with enhanced levels of GABA stimulate immune cells. Food Sci. Biotechnol. 2003, 12, 248–252. [Google Scholar]
- Tsai, J.-S.; Lin, Y.; Pan, B.; Chen, T. Antihypertensive peptides and γ-aminobutyric acid from prozyme 6 facilitated lactic acid bacteria fermentation of soymilk. Process. Biochem. 2006, 41, 1282–1288. [Google Scholar] [CrossRef]
- Park, K.-B.; Oh, S.-H. Production of yogurt with enhanced levels of gamma-aminobutyric acid and valuable nutrients using lactic acid bacteria and germinated soybean extract. Bioresour. Technol. 2007, 98, 1675–1679. [Google Scholar] [CrossRef]
- Lin, S.-D.; Mau, J.-L.; Hsu, C.-A. Bioactive components and antioxidant properties of γ-aminobutyric acid (GABA) tea leaves. LWT 2012, 46, 64–70. [Google Scholar] [CrossRef]
- Nakamura, H.; Takishima, T.; Kometani, T.; Yokogoshi, H. Psychological stress-reducing effect of chocolate enriched with γ -aminobutyric acid (GABA) in humans: Assessment of stress using heart rate variability and salivary chromogranin A. Int. J. Food Sci. Nutr. 2009, 60, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Zhao, A.; Hu, X.; Pan, L.; Wang, X. Isolation and characterization of a gamma-aminobutyric acid producing strain Lactobacillus buchneri WPZ001 that could efficiently utilize xylose and corncob hydrolysate. Appl. Microbiol. Biotechnol. 2015, 99, 3191–3200. [Google Scholar] [CrossRef] [PubMed]
- Song, H.Y.; Yu, R.C. Optimization of culture conditions for gamma-aminobutyric acid production in fermented adzuki bean milk. J. Food Drug Anal. 2018, 26, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Villegas, J.M.; Brown, L.; De Giori, G.S.; Hebert, E.M. Optimization of batch culture conditions for GABA production by Lactobacillus brevis CRL 1942, isolated from quinoa sourdough. LWT 2016, 67, 22–26. [Google Scholar] [CrossRef]
- Lim, H.S.; Cha, I.-T.; Roh, S.W.; Shin, H.-H.; Seo, M.-J. Enhanced Production of Gamma-Aminobutyric Acid by Optimizing Culture Conditions of Lactobacillus brevis HYE1 Isolated from Kimchi, a Korean Fermented Food. J. Microbiol. Biotechnol. 2017, 27, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, X.; Fu, J.; Wang, S.; Chen, Y.; Chang, K.; Li, H. Substrate sustained release-based high efficacy biosynthesis of GABA by Lactobacillus brevis NCL912. Microb. Cell Factories 2018, 17, 80. [Google Scholar] [CrossRef]
- Choi, S.I.; Lee, J.W.; Park, S.M.; Lee, M.Y.; Ji, G.E.; Park, M.S.; Heo, T.R. Improvement of γ-aminobutyric acid (GABA) production using cell entrapment of Lactobacillus brevis GABA 057. J. Microbiol. Biotechnol. 2006, 16, 562–568. [Google Scholar]
- Huang, J.; Mei, L.-H.; Wu, H.; Lin, D.-Q. Biosynthesis of γ-aminobutyric acid (GABA) using immobilized whole cells of Lactobacillus brevis. World J. Microbiol. Biotechnol. 2006, 23, 865–871. [Google Scholar] [CrossRef]
- Lee, S.; Ahn, J.; Kim, Y.-G.; Jung, J.-K.; Lee, H.; Lee, E.G. Gamma-Aminobutyric Acid Production Using Immobilized Glutamate Decarboxylase Followed by Downstream Processing with Cation Exchange Chromatography. Int. J. Mol. Sci. 2013, 14, 1728–1739. [Google Scholar] [CrossRef]
- Lyte, M. Probiotics function mechanistically as delivery vehicles for neuroactive compounds: Microbial endocrinology in the design and use of probiotics. BioEssays 2011, 33, 574–581. [Google Scholar] [CrossRef]
- Mazzoli, R.; Pessione, E.; Dufour, M.; Laroute, V.; Giuffrida, M.G.; Giunta, C.; Cocaign-Bousquet, M.; Loubière, P. Glutamate-induced metabolic changes in Lactococcus lactis NCDO 2118 during GABA production: Combined transcriptomic and proteomic analysis. Amino Acids 2010, 39, 727–737. [Google Scholar] [CrossRef] [PubMed]
- De Biase, D.; Tramonti, A.; Bossa, F.; Visca, P. The response to stationary-phase stress conditions in Escherichia coli: Role and regulation of the glutamic acid decarboxylase system. Mol. Microbiol. 1999, 32, 1198–1211. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.W. Acid Stress Responses of Salmonella and E. coli: Survival Mechanisms, Regulation, and Implications for Pathogenesis. J. Microbiol. 2001, 39, 89–94. [Google Scholar]
- Measures, J.C. Role of amino acids in osmoregulation of non-halophilic bacteria. Nat. Cell Biol. 1975, 257, 398–400. [Google Scholar] [CrossRef]
- Wu, Q.; Shah, N.P. Restoration of GABA production machinery in Lactobacillus brevis by accessible carbohydrates, anaerobiosis and early acidification. Food Microbiol. 2018, 69, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Nouaille, S.; Ribeiro, L.A.; Miyoshi, A.; Pontes, D.; Le Loir, Y.; Oliveira, S.C.; Langella, P.; De Azevedo, V. Heterologous protein production and delivery systems for Lactococcus lactis. Genet. Mol. Res. 2003, 2, 102–111. [Google Scholar]
- Even, S.; Lindley, N.D.; Cocaign-Bousquet, M. Transcriptional, translational and metabolic regulation of glycolysis in Lactococcus lactis subsp. cremoris MG 1363 grown in continuous acidic cultures. Microbiology 2003, 149, 1935–1944. [Google Scholar] [CrossRef]
- Boucher, I.; Émond, É.; Parrot, M.; Moineau, S. DNA Sequence Analysis of Three Lactococcus lactis Plasmids Encoding Phage Resistance Mechanisms. J. Dairy Sci. 2001, 84, 1610–1620. [Google Scholar] [CrossRef]
- Dougherty, B.A.; Hill, C.; Weidman, J.F.; Richardson, D.R.; Venter, J.C.; Ross, R.P. Sequence and analysis of the 60 kb conjugative, bacteriocin-producing plasmid pMRC01 fromLactococcus lactisDPC3147. Mol. Microbiol. 1998, 29, 1029–1038. [Google Scholar] [CrossRef]
- Laroute, V.; Yasaro, C.; Narin, W.; Mazzoli, R.; Pessione, E.; Cocaign-Bousquet, M.; Loubière, P. GABA Production in Lactococcus lactis Is Enhanced by Arginine and Co-addition of Malate. Front. Microbiol. 2016, 7, 1050. [Google Scholar] [CrossRef]
- Otto, R.; Brink, B.T.; Veldkamp, H.; Konings, W.N. The relation between growth rate and electrochemical proton gradient of Streptococcus cremoris. FEMS Microbiol. Lett. 1983, 16, 69–74. [Google Scholar] [CrossRef]
- Poolman, B.; Konings, W.N. Relation of growth of Streptococcus lactis and Streptococcus cremoris to amino acid transport. J. Bacteriol. 1988, 170, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Nouaille, S.; Mondeil, S.; Finoux, A.-L.; Moulis, C.; Girbal, L.; Cocaign-Bousquet, M. The stability of an mRNA is influenced by its concentration: A potential physical mechanism to regulate gene expression. Nucleic Acids Res. 2017, 45, 11711–11724. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Nomura, M.; Nakajima, I.; Fujita, Y.; Kobayashi, M.; Kimoto, H.; Suzuki, I.; Aso, H. Lactococcus lactis contains only one glutamate decarboxylase gene. Micriobiology 1999, 145, 1375–1380. [Google Scholar] [CrossRef] [PubMed]
- Van de Guchte, M.; Serror, P.; Chervaux, C.; Smokvina, T.; Ehrlich, S.D.; Maguin, E. Stress responses in lactic acid bacteria. In Lactic Acid Bacteria: Genetics, Metabolism and Applications; Siezen, R.J., Kok, J., Abee, T., Schasfsma, G., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2002; pp. 187–216. ISBN 978-90-481-6141-6. [Google Scholar]
- Csonka, L.N. Physiological and genetic responses of bacteria to osmotic stress. Microbiol. Rev. 1989, 53, 121–147. [Google Scholar] [CrossRef] [PubMed]
- Ogahara, T.; Ohno, M.; Takayama, M.; Igarashi, K.; Kobayashi, H. Accumulation of glutamate by osmotically stressed Escherichia coli is dependent on pH. J. Bacteriol. 1995, 177, 5987–5990. [Google Scholar] [CrossRef]
- Ji, H.; Lu, X.; Zong, H.; Zhuge, B. γ-aminobutyric acid accumulation enhances the cell growth of Candida glycerinogenes under hyperosmotic conditions. J. Gen. Appl. Microbiol. 2018, 64, 84–89. [Google Scholar] [CrossRef]
- Wood, J.M.; Bremer, E.; Csonka, L.N.; Kraemer, R.; Poolman, B.; Van Der Heide, T.; Smith, L.T. Osmosensing and osmoregulatory compatible solute accumulation by bacteria. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 130, 437–460. [Google Scholar] [CrossRef]
- Fichman, Y.; Gerdes, S.Y.; Kovács, H.; Szabados, L.; Zilberstein, A.; Csonka, L.N. Evolution of proline biosynthesis: Enzymology, bioinformatics, genetics, and transcriptional regulation. Biol. Rev. 2014, 90, 1065–1099. [Google Scholar] [CrossRef]
- Csonka, L.N.; Leisinger, T. Biosynthesis of Proline. EcoSal Plus 2007, 2. [Google Scholar] [CrossRef] [PubMed]
- Glaasker, E.; Tjan, F.S.B.; Ter Steeg, P.F.; Konings, W.N.; Poolman, B. Physiological Response of Lactobacillus plantarum to Salt and Nonelectrolyte Stress. J. Bacteriol. 1998, 180, 4718–4723. [Google Scholar] [CrossRef] [PubMed]
- Loos, H.; Krämer, R.; Sahm, H.; Sprenger, G.A. Sorbitol promotes growth of Zymomonas mobilis in environments with high concentrations of sugar: Evidence for a physiological function of glucose-fructose oxidoreductase in osmoprotection. J. Bacteriol. 1994, 176, 7688–7693. [Google Scholar] [CrossRef] [PubMed]
- Myers, D.K.; Lawlor, D.T.; Attfield, P.V. Influence of invertase activity and glycerol synthesis and retention on fermentation of media with a high sugar concentration by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1997, 63, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Khatibi, S.M.H.; Vahed, F.Z.; Sharifi, S.; Ardalan, M.; Shoja, M.M.; Vahed, S.Z. Osmolytes resist against harsh osmolarity: Something old something new. Biochimica 2019, 158, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, K.; Alegría, Á.; Bron, P.A.; De Angelis, M.; Gobbetti, M.; Kleerebezem, M.; Lemos, J.A.; Linares, D.M.; Ross, P.; Stanton, C.; et al. Stress Physiology of Lactic Acid Bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 837–890. [Google Scholar] [CrossRef]
- Seo, M.-J.; Nam, Y.-D.; Lee, S.-Y.; Park, S.-L.; Yi, S.-H.; Lim, S.-I. Expression and Characterization of a Glutamate Decarboxylase fromLactobacillus brevis877G Producing γ-Aminobutyric Acid. Biosci. Biotechnol. Biochem. 2013, 77, 853–856. [Google Scholar] [CrossRef]
- Lin, Q.; Li, D.; Qin, H. Molecular cloning, expression, and immobilization of glutamate decarboxylase from Lactobacillus fermentum YS2. Electron. J. Biotechnol. 2017, 27, 8–13. [Google Scholar] [CrossRef]
- Hiraga, K.; Ueno, Y.; Oda, K. Glutamate Decarboxylase fromLactobacillus brevis: Activation by Ammonium Sulfate. Biosci. Biotechnol. Biochem. 2008, 72, 1299–1306. [Google Scholar] [CrossRef]
- Sanders, J.W.; Leenhouts, K.; Burghoorn, J.; Brands, J.R.; Venema, G.; Kok, J. A chloride-inducible acid resistance mechanism in Lactococcus lactis and its regulation. Mol. Microbiol. 1998, 27, 299–310. [Google Scholar] [CrossRef]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-aminobutyric acid as a bioactive compound in foods: A review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Xu, N.; Wei, L.; Liu, J. Biotechnological advances and perspectives of gamma-aminobutyric acid production. World J. Microbiol. Biotechnol. 2017, 33, 64. [Google Scholar] [CrossRef] [PubMed]
- Chua, J.-Y.; Koh, M.K.P.; Liu, S.Q. Gamma-aminobutyric acid. In Sprouted Grains; Elsevier BV: Amsterdam, The Netherlands, 2019; pp. 25–54. [Google Scholar]
- Cui, Y.; Miao, K.; Niyaphorn, S.; Qu, X. Production of Gamma-Aminobutyric Acid from Lactic Acid Bacteria: A Systematic Review. Int. J. Mol. Sci. 2020, 21, 995. [Google Scholar] [CrossRef] [PubMed]
- Cataldo, P.G.; Villegas, J.M.; De Giori, G.S.; Saavedra, L.; Hebert, E. Enhancement of γ-aminobutyric acid (GABA) production by Lactobacillus brevis CRL 2013 based on carbohydrate fermentation. Int. J. Food Microbiol. 2020, 333, 108792. [Google Scholar] [CrossRef]
- Li, H.; Qiu, T.; Huang, G.; Cao, Y. Production of gamma-aminobutyric acid by Lactobacillus brevis NCL912 using fed-batch fermentation. Microb. Cell Factories 2010, 9, 85. [Google Scholar] [CrossRef]
- Nomura, M.; Kimoto, H.; Someya, Y.; Furukawa, S.; Suzuki, I. Production of g-Aminobutyric Acid by Cheese Starters during Cheese Ripening. J. Dairy Sci. 1998, 81, 1486–1491. [Google Scholar] [CrossRef]
- Lu, X.; Chen, Z.; Gu, Z.; Han, Y. Isolation of γ-aminobutyric acid-producing bacteria and optimization of fermentative medium. Biochem. Eng. J. 2008, 41, 48–52. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| gadB | CAACATGATCGCTGACCTTTGG | GCCATTCCACCAAGCATACAAG |
| gadC | CAGCAGAAATGGCGACGGTTG | GCTCCCCTAAAGTTTGGCTCAC |
| gadR | ATGCAAGTGCCATTAGCTGAGTAC | AGTCCCAAGCTTCGTTTTAACGG |
| tuf | AAGGAGTGGTTTGTCAGTGTCG | CTTGGTGCTTTGAACGGTGAAC |
| CDM | CDM + NaCl | Gene Expression FC by NaCl Addition | |||
|---|---|---|---|---|---|
| Time | GAD Activity | GAD Activity | |||
| (h) | mmole/min/g | mmole/min/g | gadB | gadC | gadR |
| 5 | 1.8 ± 0.4 | 36.4 ± 9.0 | 10.7 ± 3.1 | 13.6 ± 4.9 | 0.6 ± 0.3 |
| 7 | 2.3 ± 1.0 | 37.0 ± 2.0 | 17.4 ± 4.2 | 17.3 ± 3.0 | 1.4 ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laroute, V.; Mazzoli, R.; Loubière, P.; Pessione, E.; Cocaign-Bousquet, M. Environmental Conditions Affecting GABA Production in Lactococcus lactis NCDO 2118. Microorganisms 2021, 9, 122. https://doi.org/10.3390/microorganisms9010122
Laroute V, Mazzoli R, Loubière P, Pessione E, Cocaign-Bousquet M. Environmental Conditions Affecting GABA Production in Lactococcus lactis NCDO 2118. Microorganisms. 2021; 9(1):122. https://doi.org/10.3390/microorganisms9010122
Chicago/Turabian StyleLaroute, Valérie, Roberto Mazzoli, Pascal Loubière, Enrica Pessione, and Muriel Cocaign-Bousquet. 2021. "Environmental Conditions Affecting GABA Production in Lactococcus lactis NCDO 2118" Microorganisms 9, no. 1: 122. https://doi.org/10.3390/microorganisms9010122
APA StyleLaroute, V., Mazzoli, R., Loubière, P., Pessione, E., & Cocaign-Bousquet, M. (2021). Environmental Conditions Affecting GABA Production in Lactococcus lactis NCDO 2118. Microorganisms, 9(1), 122. https://doi.org/10.3390/microorganisms9010122