Microbiota of the Digestive Gland of Red Abalone (Haliotis rufescens) Is Affected by Withering Syndrome

 , and
, and 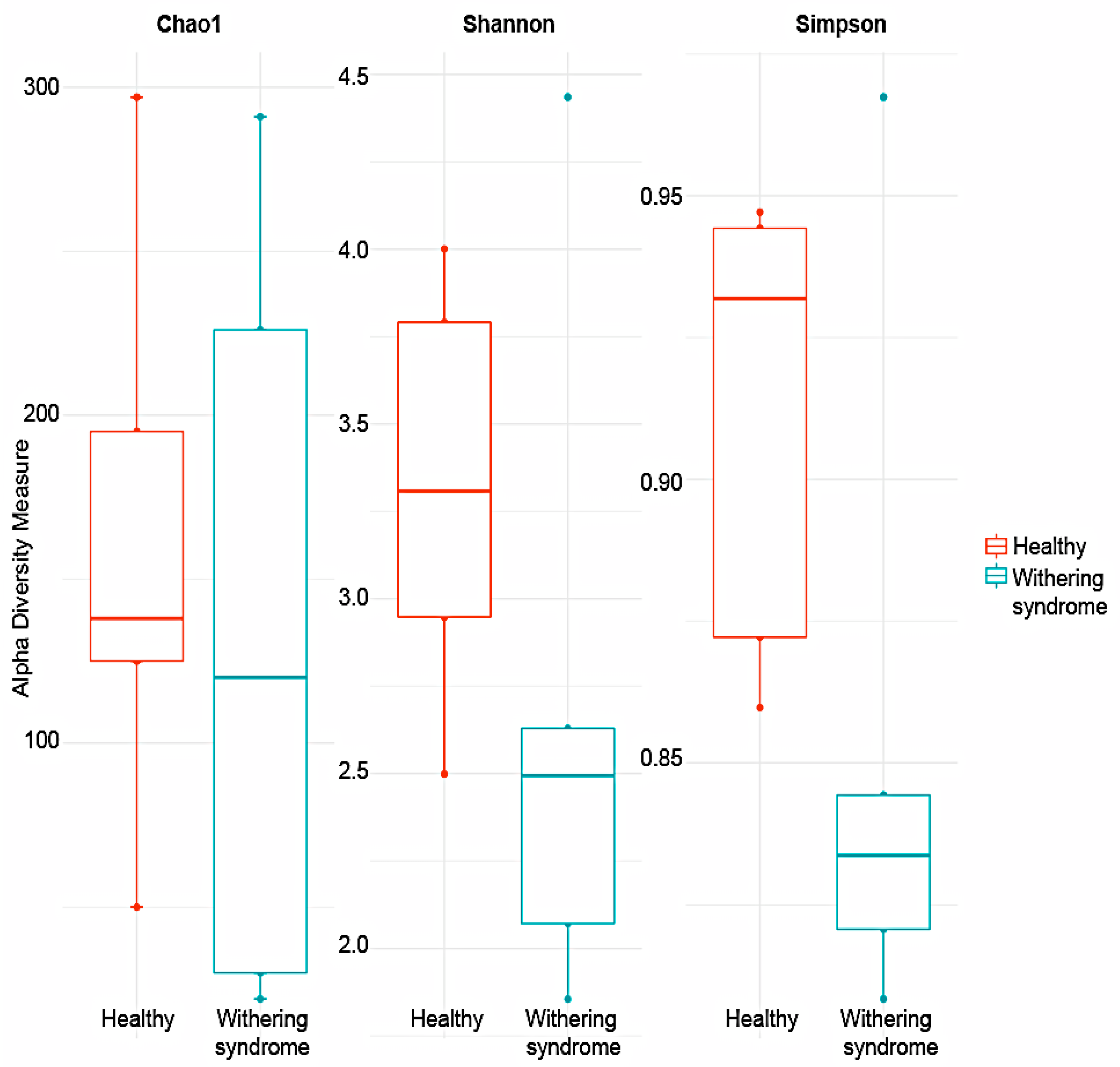{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction and Sequencing
2.3. Bioinformatic Analysis
2.4. Ethical Notes
2.5. Statistical Analysis
3. Results
3.1. High-Throughput Sequence Data
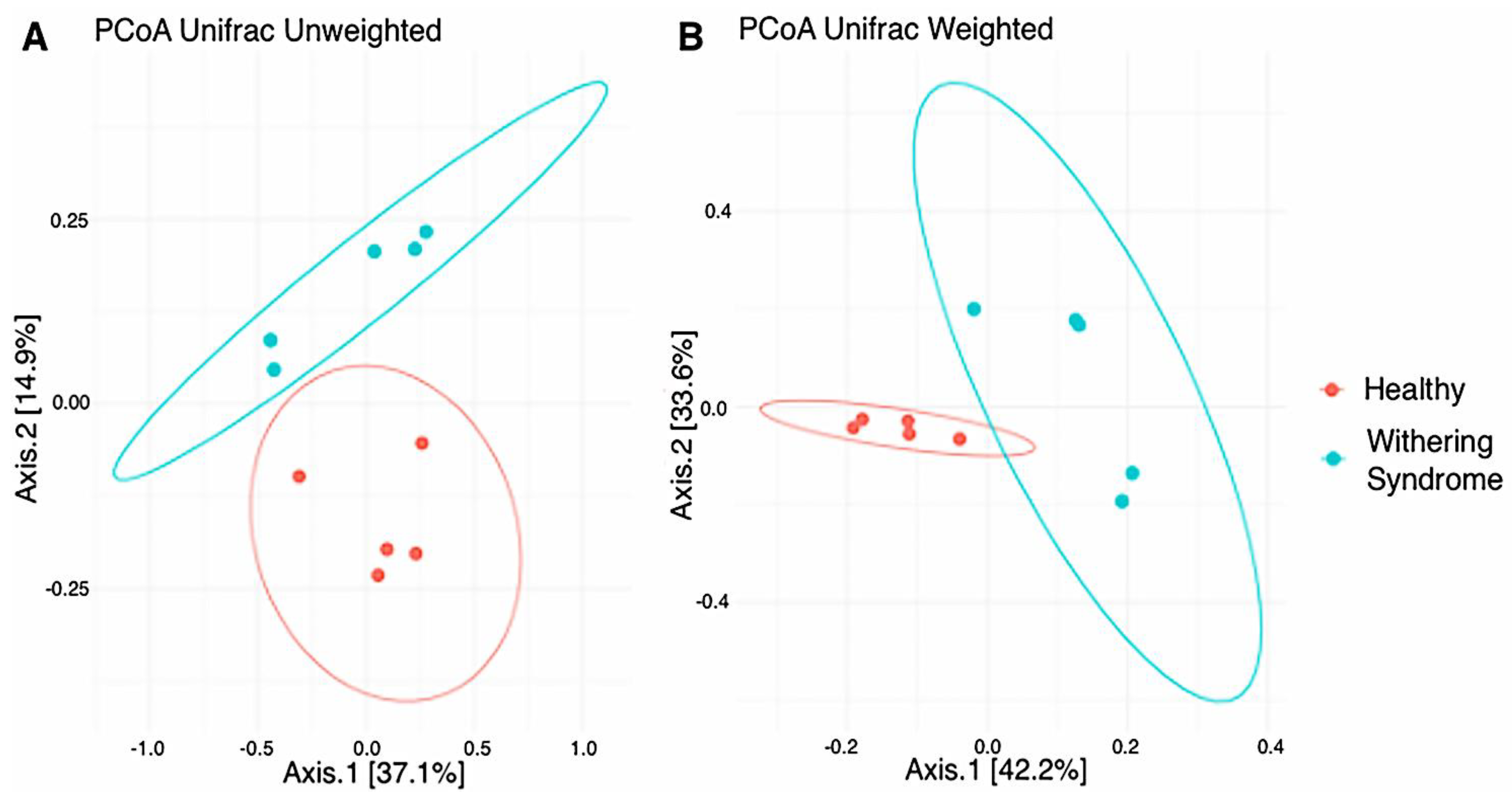
3.2. Analysis of Microbiota Diversity
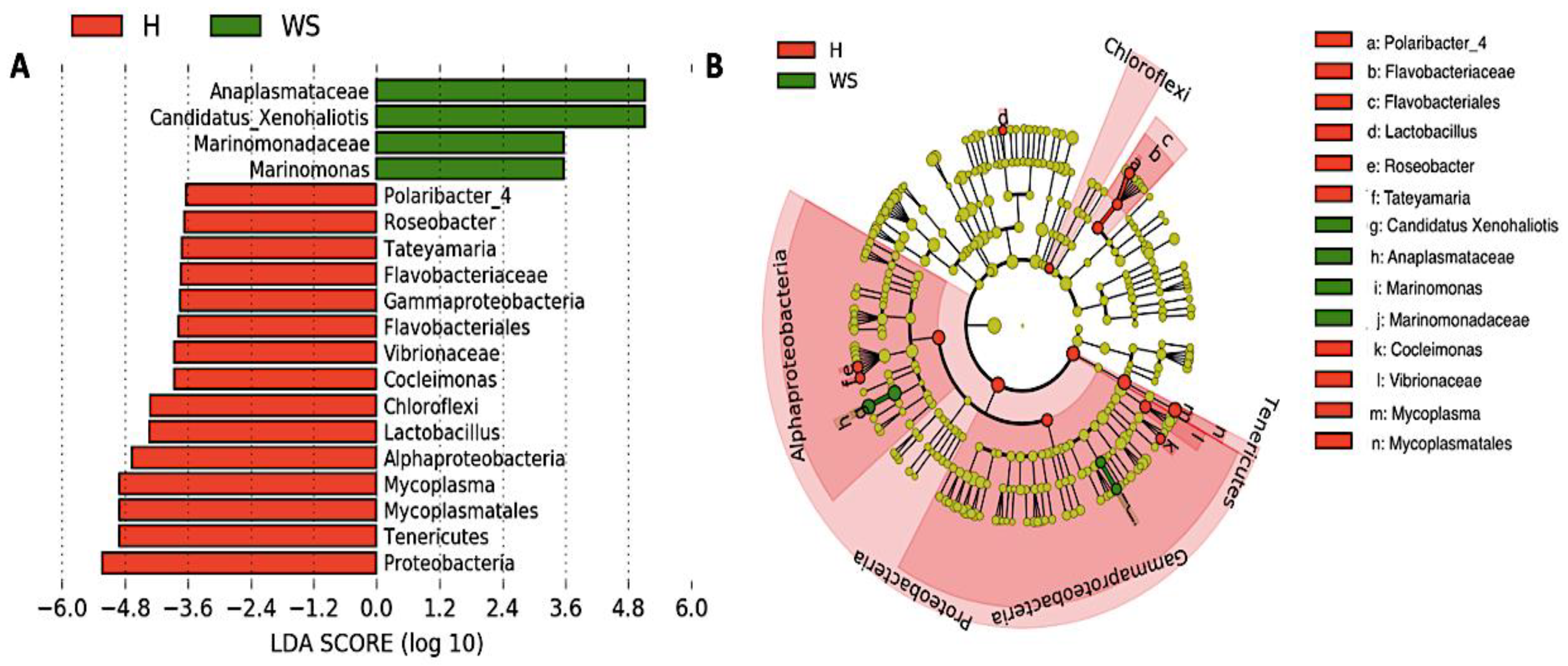
3.3. Taxonomic Composition and Differential Abundance of Bacterial Communities of Digestive Gland of Healthy Red Abalone and Red Abalone with WS Disease
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2018. Available online: http://www.fao.org/state-of-fisheries-aquaculture (accessed on 18 October 2018).
- Flores-Aguilar, R.A.; Gutiérrez, A.; Ellwanger, A.; Searcy-Bernal, R. Development and current status of abalone aquaculture in Chile. J. Shellfish Res. 2007, 26, 705–711. [Google Scholar] [CrossRef]
- Enríquez, R.; Villagrán, R. La experiencia del desarrollo del cultivo de abalón (Haliotis spp.) en Chile: Oportunidades y desafíos. OIE Rev. Sci. Tech. 2008, 27, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Brokordt, K.; González, R.; Farías, W.; Winkler, F.E.; Lohrmann, K.B. First insight into the heritable variation of the resistance to infection with the bacteria causing the withering syndrome disease in Haliotis rufescens abalone. J. Invertebr. Pathol. 2017, 150, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Crosson, L.M.; Wight, N.; VanBlaricom, G.R.; Kiryu, I.; Moore, J.D.; Friedman, C.S. Abalone withering syndrome: Distribution, impacts, current diagnostic methods and new findings. Dis. Aquat. Org. 2014, 108, 261–270. [Google Scholar] [CrossRef]
- Honda, K.; Littman, D.R. The microbiome in infectious disease and inflammation. Annu. Rev. Immunol. 2012, 30, 759–795. [Google Scholar] [CrossRef]
- Corthier, G.; Doré, J. Une ère nouvelle dans le domaine des interactions entre le microbiote et la santé humaine. Gastroenterol. Clin. Biol. 2010, 34, S1–S6. [Google Scholar] [CrossRef]
- Libertucci, J.; Young, V.B. The role of the microbiota in infectious diseases. Nat. Microbiol. 2019, 4, 35–45. [Google Scholar] [CrossRef]
- Cáceres-Martínez, J.; Vásquez-Yeomans, R.; Flores-Saaib, R.D. Intracellular prokaryote Xenohaliotis californiensis in abalone Haliotis spp. from Baja California, México. Cienc. Pesq. 2011, 19, 5–11. [Google Scholar]
- Friedman, C.S.; Biggs, W.; Shields, J.D.; Hedrick, R.P. Transmission of withering syndrome in black abalone, Haliotis cracherodii Leach. J. Shellfish Res. 2002, 21, 817–824. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package 2011. R Package Version 1.17–10. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 10 March 2019).
- Segata, N.; Bornigen, D.; Morgan, X.C.; Huttenhower, C. PhyloPhlAn is a new method for improved phylogenetic and taxonomic placement of microbes. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Neu, A.T.; Allen, E.E.; Roy, K. Diversity and composition of intertidal gastropod microbiomes across a major marine biogeographic boundary. Environ. Microbiol. Rep. 2019, 11, 434–447. [Google Scholar] [CrossRef]
- Ito, M.; Watanabe, K.; Maruyama, T.; Mori, T.; Niwa, K.; Chow, S.; Takeyama, H. Enrichment of bacteria and alginate lyase genes potentially involved in brown alga degradation in the gut of marine gastropods. Sci. Rep. 2019, 9, 2129. [Google Scholar] [CrossRef]
- Gobet, A.; Mest, L.; Perennou, M.; Dittami, S.M.; Caralp, C.; Coulombet, C.; Huchette, S.; Roussel, S.; Michel, G.; Leblanc, C. Seasonal and algal diet-driven patterns of the digestive microbiota of the European abalone Haliotis tuberculata, a generalist marine herbivore. Microbiome 2018, 6, 60. [Google Scholar] [CrossRef]
- Aceves, A.K.; Johnson, P.; Bullard, S.A.; Lafrentz, S.; Arias, C.R. Description and characterization of the digestive gland microbiome in the freshwater mussel Villosa nebulosa (Bivalvia: Unionidae). J. Molluscan Stud. 2018, 84, 240–246. [Google Scholar] [CrossRef]
- Hollants, J.; Leroux, O.; Leliaert, F.; Decleyre, H.; De Clerck, O.; Willems, A. Who is in there? Exploration of endophytic bacteria within the siphonous green seaweed Bryopsis (Bryopsidales, Chlorophyta). PLoS ONE 2011, 6, e26458. [Google Scholar] [CrossRef]
- Hollants, J.; Leliaert, F.; Verbruggen, H.; Willems, A.; De Clerck, O. Permanent residents or temporary lodgers: Characterizing intracellular bacterial communities in the siphonous green alga Bryopsis. Proc. R. Soc. B. 2013, 280, 20122659. [Google Scholar] [CrossRef]
- Cicala, F.; Cisterna-Céliz, J.A.; Moore, J.D.; Rocha-Olivares, A. Structure, dynamics and predicted functional role of the gut microbiota of the blue (Haliotis fulgens) and yellow (H. corrugata) abalone from Baja California Sur, Mexico. PeerJ 2018, 6, e5830. [Google Scholar] [CrossRef]
- Aronson, H.S.; Zellmer, A.J.; Goffredi, S.K. The specific and exclusive microbiome of the deep-sea bone-eating snail, Rubyspira osteovora. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed]
- Pierce, M.L.; Ward, J.E. Gut microbiomes of the eastern oyster (Crassostrea virginica) and the blue mussel (mytilus edulis): Temporal variation and the influence of marine aggregate-associated microbial communities. MSphere 2019, 4, e00730-19. [Google Scholar] [CrossRef] [PubMed]
- Razin, S.; Yogev, D.; Naot, Y. Molecular biology and pathogenicity of mycoplasmas. Microbiol. Mol. Biol. Rev. 1998, 62, 1094–1156. [Google Scholar] [CrossRef]
- Register, K.B.; Thole, L.; Rosenbush, R.F.; Minion, F.C. Multilocus sequence typing of Mycoplasma bovis reveals host-specific genotypes in cattle versus bison. Vet. Microbiol. 2015, 175, 92–98. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, J.M.; Wang, S.L.; Gao, Z.M.; Zhang, A.Q.; Danchin, A.; He, L.S. Genomic characterization of symbiotic mycoplasmas from the stomach of deep-sea isopod bathynomus sp. Environ. Microbiol. 2016, 18, 2646–2659. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.R.; Zacher, L.A.; Wendland, L.D.; Brown, M.R. Emerging mycoplasmoses in wildlife. In Mycoplasmas Molecular Biology, Pathogenicity and Strategies for Control, 1st ed.; Blanchard, A., Browing, G., Eds.; Horizon Bioscience: Norwich, UK, 2005; pp. 383–414. [Google Scholar]
- Ludwig, W.; Euzéby, J.; Whitman, W.B. Road map of the phyla Bacteroidetes, Spirochaetes, Tenericutes (Mollicutes), Acidobacteria, Fibrobacteres, Fusobacteria, Dictyoglomi, Gemmatimonadetes, Lentisphaerae, Verrucomicrobia, Chlamydiae, and Planctomycetes. In Bergey’s Manual of Systematic Bacteriology; Krieg, N.R., Ludwig, W., Whitman, W., Hedlund, B.P., Paster, B.J., Staley, J.T., Ward, N., Brown, D., Parte, A., Eds.; Springer: New York, NY, USA, 2010; Volume 4, pp. 1–19. [Google Scholar]
- Fraune, S.; Zimmer, M. Host-specificity of environmentally transmitted mycoplasma-like isopod symbionts. Environ. Microbiol. 2008, 10, 2497–2504. [Google Scholar] [CrossRef] [PubMed]
- Duperron, S.; Pottier, M.A.; Leger, N.; Gaudron, S.M.; Puillandre, N.; Le Prieur, S.; Sigwart, J.D.; Ravaux, J.; Zbinden, M. A tale of two chitons: Is habitat specialisation linked to distinct associated bacterial communities? FEMS Microbiol. Ecol. 2013, 83, 552–567. [Google Scholar] [CrossRef]
- Henrich, B.; Feldmann, R.C.; Hadding, U. Cytoadhesins of Mycoplasma hominis. Infect. Immun. 1993, 61, 2945–2951. [Google Scholar] [CrossRef]
- Severi, E.; Hood, D.W.; Thomas, G.H. Sialic acid utilization by bacterial pathogens. Microbiology 2007, 153, 2817–2822. [Google Scholar] [CrossRef]
- Pérez-Acosta, J.; Martínez-Porchas, M.; Gollas-Galván, T.; Martínez-Córdova, L.R.; Gutiérrez-Millán, L.E.; López-Torre, M. Proteínas transmembranales de organismos tipo rickettsia (OTR) en animales acuáticos: Factores de adherencia, invasión e infección. Rev. Biol. Mar. Oceanogr. 2017, 52, 19–32. [Google Scholar] [CrossRef]
- Li, H.; Walker, D.H. rOmpA is a critical protein for the adhesion of Rickettsia rickettsii to host cells. Microb. Pathog. 1998, 24, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Palmer, G.H.; Noh, S.M. Rickettsial entry into host cells: Finding the keys to unlock the doors. Infect. Immun. 2012, 80, 3746–3747. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.K. Probiotics and immunity: A fish perspective. Fish Shellfish Immunol. 2010, 29, 2–14. [Google Scholar] [CrossRef]
- Catalán, N.; Villasante, A.; Wacyk, J.; Ramírez, C.; Romero, J. Fermented Soybean Meal Increases Lactic Acid Bacteria in Gut Microbiota of Atlantic Salmon (Salmon salar). Probiotics Antimicrob. Proteins 2018, 10, 566–576. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Sun, Y.Z.; Wang, A.; Zhou, Z. Probiotics as Means of Diseases Control in Aquaculture, a Review of Current Knowledge and Future Perspectives. Front. Microbiol. 2018, 9, 2429. [Google Scholar] [CrossRef]
- Ringø, E. Probiotics in shellfish aquaculture. Aquac. Fish. 2020, 5, 1–27. [Google Scholar] [CrossRef]
- Martínez, P.; Ibáñez, A.L.; Monroy, O.A.; Ramírez, H.C. Use of probiotics in aquaculture. ISRN Microbiol. 2012, 2012, 916845. [Google Scholar] [CrossRef]
- Mohapatra, S.; Chakraborty, T.; Kumar, V.; DeBoeck, G.; Mohanta, K.N. Aquaculture and stress management: A review of probiotic intervention. J. Anim. Physiol. Anim. Nutr. 2013, 97, 405–430. [Google Scholar] [CrossRef]
- Wang, Y.B.; Li, J.R.; Lin, J. Probiotics in aquaculture: Challenges and outlook. Aquaculture 2008, 281, 1–4. [Google Scholar] [CrossRef]
- Pandiyan, P.; Balaraman, D.; Thirunavukkarasu, R.; Jothi, E.D.; Subaramaniyan, K.; Manikkam, S.; Sadayappan, B. Probiotics in aquaculture. Drug Invent. Today 2013, 5, 55–59. [Google Scholar] [CrossRef]
- Lulijwa, R.; Rupia, E.J.; Alfaro, A.C. Antibiotic use in aquaculture, policies and regulation, health and environmental risk: A review of the top 15 major producers. Rev. Aquacult. 2019, 12, 640–663. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villasante, A.; Catalán, N.; Rojas, R.; Lohrmann, K.B.; Romero, J. Microbiota of the Digestive Gland of Red Abalone (Haliotis rufescens) Is Affected by Withering Syndrome. Microorganisms 2020, 8, 1411. https://doi.org/10.3390/microorganisms8091411
Villasante A, Catalán N, Rojas R, Lohrmann KB, Romero J. Microbiota of the Digestive Gland of Red Abalone (Haliotis rufescens) Is Affected by Withering Syndrome. Microorganisms. 2020; 8(9):1411. https://doi.org/10.3390/microorganisms8091411
Chicago/Turabian StyleVillasante, Alejandro, Natalia Catalán, Rodrigo Rojas, Karin B. Lohrmann, and Jaime Romero. 2020. "Microbiota of the Digestive Gland of Red Abalone (Haliotis rufescens) Is Affected by Withering Syndrome" Microorganisms 8, no. 9: 1411. https://doi.org/10.3390/microorganisms8091411
APA StyleVillasante, A., Catalán, N., Rojas, R., Lohrmann, K. B., & Romero, J. (2020). Microbiota of the Digestive Gland of Red Abalone (Haliotis rufescens) Is Affected by Withering Syndrome. Microorganisms, 8(9), 1411. https://doi.org/10.3390/microorganisms8091411