The Availability of Iron Is Involved in the Murine Experimental Toxoplasma gondii Infection Outcome

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Parasites
2.3. Experimental Design
2.4. Iron Staining in the Small Intestine, Lung, and Liver of Infected Mice
2.5. Iron Levels in Serum Samples
2.6. Serum Levels of Pyruvic Transaminase
2.7. Histological Analysis
2.8. Immunohistochemical Analysis for Detection of Tissue Parasitism
2.9. Relative Quantification of mRNA by qPCR
2.10. Relative Quantification of Intestinal Bacteria by qPCR
2.11. Cytometric Bead Array Assay
2.12. Statistical Analysis
3. Results
3.1. Toxoplasma gondii Infection Induces Iron Accumulation in the Organs of Infected Animals
3.2. Infection with T. gondii Interferes in the Expression Genes Involved in the Iron Absorption by the Small Intestine
3.3. T. gondii Infection Increases the Iron Concentration Systemically on Day 8 of Infection
3.4. Iron Availability is Involved in the Parasite Growth and Inflammation in the Small Intestine
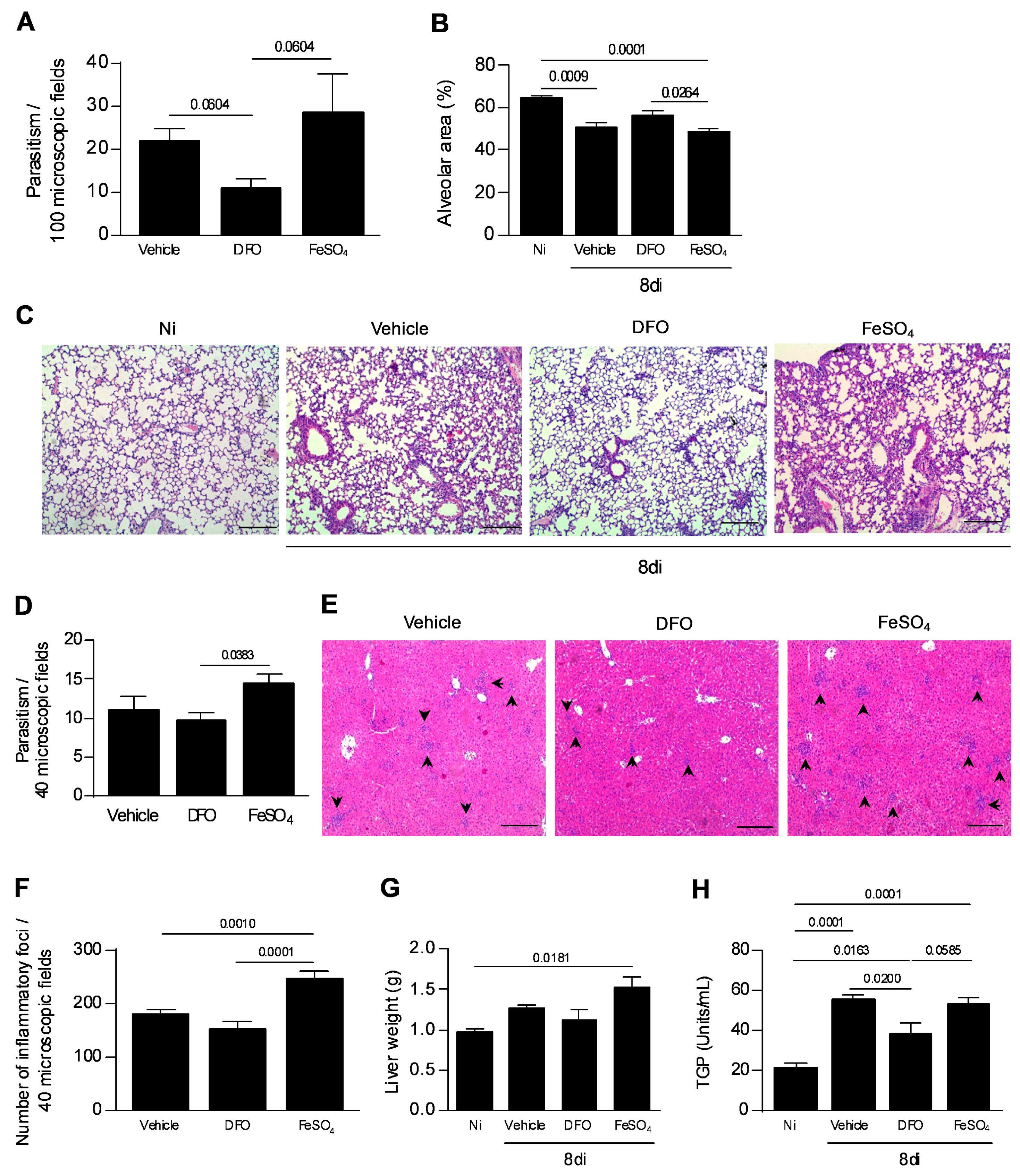
3.5. Iron Availability Is Involved in the T. gondii Proliferation and Spread to the Other Organs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ganz, T.; Nemeth, E. Iron homeostasis in host defence and inflammation. Nat. Rev. Immunol. 2015, 15, 500–510. [Google Scholar] [CrossRef]
- McKie, A.T.; Barrow, D.; Latunde-Dada, G.O.; Rolfs, A.; Sager, G.; Mudaly, E.; Mudaly, M.; Richardson, C.; Barlow, D.; Bomford, A.; et al. An iron-regulated ferric reductase associated with the absorption of dietary iron. Science 2001, 291, 1755–1759. [Google Scholar] [CrossRef] [PubMed]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Abboud, S.; Haile, D.J. A novel mammalian iron-regulated protein involved in intracellular iron metabolism. J. Biol. Chem. 2000, 275, 19906–19912. [Google Scholar] [CrossRef] [PubMed]
- Donovan, A.; Brownlie, A.; Zhou, Y.; Shepard, J.; Pratt, S.J.; Moynihan, J.; Paw, B.H.; Drejer, A.; Barut, B.; Zapata, A.; et al. Positional cloning of zebrafish ferroportin1 identifies a conserved vertebrate iron exporter. Nature 2000, 403, 776–781. [Google Scholar] [CrossRef]
- McKie, A.T.; Marciani, P.; Rolfs, A.; Brennan, K.; Wehr, K.; Barrow, D.; Miret, S.; Bomford, A.; Peters, T.J.; Farzaneh, F.; et al. A novel duodenal iron-regulated transporter, IREG1, implicated in the basolateral transfer of iron to the circulation. Mol. Cell 2000, 5, 299–309. [Google Scholar] [CrossRef]
- Mitchell, C.J.; Shawki, A.; Ganz, T.; Nemeth, E.; Mackenzie, B. Functional properties of human ferroportin, a cellular iron exporter reactive also with cobalt and zinc. Am. J. Physiol. Cell Physiol. 2014, 306, 450–459. [Google Scholar] [CrossRef]
- Huebers, H.; Huebers, E.; Rummel, W.; Crichton, R.R. Isolation and characterization of iron-binding proteins from rat intestinal mucosa. Eur. J. Biochem. 1976, 66, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Farnaud, S.; Evans, R.W. Lactoferrin—A multifunctional protein with antimicrobial properties. Mol. Immunol. 2003, 40, 395–405. [Google Scholar] [CrossRef]
- Schneider, B.D.; Leibold, E.A. Regulation of mammalian iron homeostasis. Curr. Opin. Clin. Nutr. Metab. Care 2000, 3, 267–273. [Google Scholar] [CrossRef]
- Muckenthaler, M.U.; Galy, B.; Hentze, M.W. Systemic iron homeostasis and the iron-responsive element/iron-regulatory protein (IRE/IRP) regulatory network. Annu. Rev. Nutr. 2008, 28, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [PubMed]
- Peyssonnaux, C.; Zinkernagel, A.S.; Datta, V.; Lauth, X.; Johnson, R.S.; Nizet, V. TLR4-dependent hepcidin expression by myeloid cells in response to bacterial pathogens. Blood 2006, 107, 3727–3732. [Google Scholar] [CrossRef] [PubMed]
- Skaar, E.P. The battle for iron between bacterial pathogens and their vertebrate hosts. PLoS Pathog. 2010, 6, e1000949. [Google Scholar] [CrossRef] [PubMed]
- Huynh, C.; Sacks, D.L.; Andrews, N.W. A Leishmania amazonensis ZIP family iron transporter is essential for parasite replication within macrophage phagolysosomes. J. Exp. Med. 2006, 203, 2363–2375. [Google Scholar] [CrossRef]
- Huynh, C.; Andrews, N.W. Iron acquisition within host cells and the pathogenicity of Leishmania. Cell. Microbiol. 2008, 10, 293–300. [Google Scholar] [CrossRef]
- Wilson, M.E.; Vorhies, R.W.; Andersen, K.A.; Britigan, B.E. Acquisition of iron from transferrin and lactoferrin by the protozoan Leishmania chagasi. Infect. Immun. 1994, 62, 3262–3269. [Google Scholar] [CrossRef]
- Lehker, M.W.; Alderete, J.F. Iron regulates growth of Trichomonas vaginalis and the expression of immunogenic trichomonad proteins. Mol. Microbiol. 1992, 6, 123–132. [Google Scholar] [CrossRef]
- Loo, V.G.; Lalonde, R.G. Role of iron in intracellular growth of Trypanosoma cruzi. Infect. Immun. 1984, 45, 726–730. [Google Scholar] [CrossRef]
- Dimier, I.H.; Bout, D.T. Interferon-gamma-activated primary enterocytes inhibit Toxoplasma gondii replication: A role for intracellular iron. Immunology 1998, 94, 488–495. [Google Scholar] [CrossRef]
- Dziadek, B.; Dziadek, J.; Dlugonska, H. Identification of Toxoplasma gondii proteins binding human lactoferrin: A new aspect of rhoptry proteins function. Exp. Parasitol. 2007, 115, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.P.O.; Ferro, E.A.V.; Briceño, M.P.P.; Oliveira, M.C.; Barbosa, B.F.; Silva, N.M. Susceptibility of human villous (BeWo) and extravillous (HTR-8/SVneo) trophoblast cells to Toxoplasma gondii infection is modulated by intracellular iron availability. Parasitol. Res. 2019, 118, 1559–1572. [Google Scholar] [CrossRef] [PubMed]
- Portugal, S.; Carret, C.; Recker, M.; Armitage, A.E.; Gonçalves, L.A.; Epiphanio, S.; Sullivan, D.; Roy, C.; Newbold, C.I.; Drakesmith, H.; et al. Host-mediated regulation of superinfection in malaria. Nat. Med. 2011, 17, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.S. Effect of deferoxamine alone and combined with pyrimethamine on acute toxoplasmosis in mice. J. Egypt. Soc. Parasitol. 1999, 29, 791–803. [Google Scholar]
- Dubey, J.P.; Ferreira, L.R.; Martins, J.; Mcleod, R. Oral oocyst-induced mouse model of toxoplasmosis: Effect of infection with Toxoplasma gondii strains of different genotypes, dose, and mouse strains (transgenic, out-bred, in-bred) on pathogenesis and mortality. Parasitology 2012, 139, 1–13. [Google Scholar] [CrossRef]
- Benevides, L.; Milanezi, C.M.; Yamauchi, L.M.; Benjamim, C.F.; Silva, J.S.; Silva, N.M. CCR2 receptor is essential to activate microbicidal mechanisms to control Toxoplasma gondii infection in the central nervous system. Am. J. Pathol. 2008, 173, 741–751. [Google Scholar] [CrossRef]
- Araujo, E.C.; Barbosa, B.F.; Coutinho, L.B.; Barenco, P.V.; Sousa, L.A.; Milanezi, C.M.; Bonfá, G.; Pavanelli, W.R.; Silva, J.S.; Ferro, E.A.; et al. Heme oxygenase-1 activity is involved in the control of Toxoplasma gondii infection in the lung of BALB/c and C57BL/6 and in the small intestine of C57BL/6 mice. Vet. Res. 2013, 44, 1–14. [Google Scholar] [CrossRef]
- Barenco, P.V.; Lourenço, E.V.; Cunha-Júnior, J.P.; Almeida, K.C.; Roque-Barreira, M.C.; Silva, D.A.; Araújo, E.C.; Coutinho, L.B.; Oliveira, M.C.; Mineo, T.W.; et al. Toxoplasma gondii 70 kDa heat shock protein: Systemic detection is associated with the death of the parasites by the immune response and its increased expression in the brain is associated with parasite replication. PLoS ONE 2014, 9, e96527. [Google Scholar] [CrossRef]
- Guo, X.; Xia, X.; Tang, R.; Zhou, J.; Zhao, H.; Wang, K. Development of a real-time PCR method for Firmicutes and Bacteroidetes in faeces and its application to quantify intestinal population of obese and lean pigs. Lett. Appl. Microbiol. 2008, 47, 367–373. [Google Scholar] [CrossRef]
- Barman, M.; Unold, D.; Shifley, K.; Amir, E.; Hung, K.; Bos, N.; Salzman, N. Enteric salmonellosis disrupts the microbial ecology of the murine gastrointestinal tract. Infect. Immun. 2008, 76, 907–915. [Google Scholar] [CrossRef]
- Jiang, Q.; Fu, B.; Chen, Y.; Wang, Y.; Liu, H. Quantification of viable but nonculturable bacterial pathogens in anaerobic digested sludge. Appl. Microbiol. Biotechnol. 2013, 97, 6043–6050. [Google Scholar] [CrossRef] [PubMed]
- Mitsunaga, T.; Norose, K.; Aosai, F.; Horie, H.; Ohnuma, N.; Yano, A. Infection dynamics of Toxoplasma gondii in gut-associated tissues after oral infection: The role of Peyer’s patches. Parasitol. Int. 2019, 68, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Bereswill, S.; Muñoz, M.; Fischer, A.; Plickert, R.; Haag, L.M.; Otto, B.; Kühl, A.A.; Loddenkemper, C.; Göbel, U.B.; Heimesaat, M.M. Anti-inflammatory effects of resveratrol, curcumin and simvastatin in acute small intestinal inflammation. PLoS ONE 2010, 5, e15099. [Google Scholar] [CrossRef] [PubMed]
- Raetz, M.; Hwang, S.H.; Wilhelm, C.L.; Kirkland, D.; Benson, A.; Sturge, C.R.; Mirpuri, J.; Vaishnava, S.; Hou, B.; Defranco, A.L.; et al. Parasite-induced TH1 cells and intestinal dysbiosis cooperate in IFN-γ-dependent elimination of Paneth cells. Nat. Immunol. 2013, 14, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Dogruman-al, F.; Fidan, I.; Celebi, B.; Yesilyurt, E.; Erdal, B.; Babur, C.; Kustimur, S. Cytokine profile in murine toxoplasmosis. Asian Pac. J. Trop. Med. 2011, 4, 16–19. [Google Scholar] [CrossRef]
- Sibley, L.D.; Messina, M.; Niesman, I.R. Stable DNA transformation in the obligate intracellular parasite Toxoplasma gondii by complementation of tryptophan auxotrophy. Proc. Natl. Acad. Sci. USA 1994, 91, 5508–5512. [Google Scholar] [CrossRef]
- Kirk, K.; Horner, H.A.; Kirk, J. Glucose uptake in Plasmodium falciparum-infected erythrocytes is an equilibrative not an active process. Mol. Biochem. Parasitol. 1996, 82, 195–205. [Google Scholar] [CrossRef]
- Coppens, I.; Sinai, A.P.; Joiner, K.A. Toxoplasma gondii exploits host low-density lipoprotein receptor-mediated endocytosis for cholesterol acquisition. J. Cell Biol. 2000, 149, 167–180. [Google Scholar] [CrossRef]
- Tanaka, T.; Abe, Y.; Inoue, N.; Kim, W.S.; Kumura, H.; Nagasawa, H.; Igarashi, I.; Shimazaki, K. The detection of bovine lactoferrin binding protein on Trypanosoma brucei. J. Vet. Med. Sci. 2004, 66, 619–625. [Google Scholar] [CrossRef]
- Peterson, K.M.; Alderete, J.F. Iron uptake and increased intracellular enzyme activity follow host lactoferrin binding by Trichomonas vaginalis receptors. J. Exp. Med. 1984, 160, 398–410. [Google Scholar] [CrossRef]
- León-Sicairos, N.; Reyes-López, M.; Canizalez-Román, A.; Bermúdez-Cruz, R.M.; Serrano-Luna, J.; Arroyo, R.; de la Garza, M. Human hololactoferrin: Endocytosis and use as an iron source by the parasite Entamoeba histolytica. Microbiology 2005, 151, 3859–3871. [Google Scholar] [CrossRef] [PubMed]
- Schell, D.; Borowy, N.K.; Overath, P. Transferrin is a growth factor for the bloodstream form of Trypanosoma brucei. Parasitol. Res. 1991, 77, 558–560. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Lindsay, D.S.; Speer, C.A. Structures of Toxoplasma gondii tachyzoites, bradyzoites, and sporozoites and biology and development of tissue cysts. Clin. Microbiol. Rev. 1998, 11, 267–299. [Google Scholar] [CrossRef] [PubMed]
- Blader, I.J.; Manger, I.D.; Boothroyd, J.C. Microarray analysis reveals previously unknown changes in Toxoplasma gondii-infected human cells. J. Biol. Chem. 2001, 276, 24223–24231. [Google Scholar] [CrossRef] [PubMed]
- Gail, M.; Gross, U.; Bohne, W. Transcriptional profile of Toxoplasma gondii-infected human fibroblasts as revealed by gene-array hybridization. Mol. Genet. Genom. 2001, 265, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Gail, M.; Gross, U.; Bohne, W. Transferrin receptor induction in Toxoplasma gondii-infected HFF is associated with increased iron-responsive protein 1 activity and is mediated by secreted factors. Parasitol. Res. 2004, 94, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Dziadek, B.; Dzitko, K.; Dlugonska, H. Toxoplasma gondii binds human lactoferrin but not transferrin. Exp. Parasitol. 2005, 110, 165–167. [Google Scholar] [CrossRef]
- Deschemin, J.C.; Vaulont, S. Role of hepcidin in the setting of hypoferremia during acute inflammation. PLoS ONE 2013, 8, e61050. [Google Scholar] [CrossRef]
- Anderson, G.J.; Frazer, D.M.; Wilkins, S.J.; Becker, E.M.; Millard, K.N.; Murphy, T.L.; McKie, A.T.; Vulpe, C.D. Relationship between intestinal iron-transporter expression, hepatic hepcidin levels and the control of iron absorption. Biochem. Soc. Trans. 2002, 30, 724–726. [Google Scholar] [CrossRef]
- Nicolas, G.; Bennoun, M.; Devaux, I.; Beaumont, C.; Grandchamp, B.; Kahn, A.; Vaulont, S. Lack of hepcidin gene expression and severe tissue iron overload in upstream stimulatory factor 2 (USF2) knockout mice. Proc. Natl. Acad. Sci. USA 2001, 98, 8780–8785. [Google Scholar] [CrossRef]
- Park, C.H.; Valore, E.V.; Waring, A.J.; Ganz, T. Hepcidin, a urinary antimicrobial peptide synthesized in the liver. J. Biol. Chem. 2001, 276, 7806–7810. [Google Scholar] [CrossRef] [PubMed]
- Rachidi, S.; Coudray, C.; Baret, P.; Gelon, G.; Pierre, J.L.; Favier, A. Inhibition of lipid peroxidation by a new family of iron chelators. Comparison with desferrioxamine. Biol. Trace Elem. Res. 1994, 41, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Vermylen, C. What is new in iron overload? Eur. J. Pediatr. 2008, 167, 377–381. [Google Scholar] [CrossRef]
- Hershko, C.; Peto, T.E. Deferoxamine inhibition of malaria is independent of host iron status. J. Exp. Med. 1988, 168, 375–387. [Google Scholar] [CrossRef]
- Arantes, J.M.; Francisco, A.F.; de Abreu Vieira, P.M.; Silva, M.; Araújo, M.S.; de Carvalho, A.T.; Pedrosa, M.L.; Carneiro, C.M.; Tafuri, W.L.; Martins-Filho, O.A.; et al. Trypanosoma cruzi: Desferrioxamine decreases mortality and parasitemia in infected mice through a trypanostatic effect. Exp. Parasitol. 2011, 128, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Paiva, C.N.; Feijó, D.F.; Dutra, F.F.; Carneiro, V.C.; Freitas, G.B.; Alves, L.S.; Mesquita, J.; Fortes, G.B.; Figueiredo, R.T.; Souza, H.S.; et al. Oxidative stress fuels Trypanosoma cruzi infection in mice. J. Clin. Investig. 2012, 122, 2531–2542. [Google Scholar] [CrossRef]
- Kurashima, Y.; Yamamoto, D.; Nelson, S.; Uematsu, S.; Ernst, P.B.; Nakayama, T.; Kiyono, H. Mucosal Mesenchymal Cells: Secondary Barrier and Peripheral Educator for the Gut Immune System. Front. Immunol. 2017, 8, 1787. [Google Scholar] [CrossRef]
- Bevins, C.L.; Salzman, N.H. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat. Rev. Microbiol. 2011, 9, 356–368. [Google Scholar] [CrossRef]
- Villeret, B.; Brault, L.; Couturier-Maillard, A.; Robinet, P.; Vasseur, V.; Secher, T.; Dimier-Poisson, I.; Jacobs, M.; Zheng, S.G.; Quesniaux, V.F.; et al. Blockade of IL-1R signaling diminishes Paneth cell depletion and Toxoplasma gondii induced ileitis in mice. Am. J. Clin. Exp. Immunol. 2013, 2, 107–116. [Google Scholar]
- Burger, E.; Araujo, A.; López-Yglesias, A.; Rajala, M.W.; Geng, L.; Levine, B.; Hooper, L.V.; Burstein, E.; Yarovinsky, F. Loss of Paneth Cell Autophagy Causes Acute Susceptibility to Toxoplasma gondii-Mediated Inflammation. Cell Host Microbe 2018, 23, 177–190. [Google Scholar] [CrossRef]
- Courret, N.; Darche, S.; Sonigo, P.; Milon, G.; Buzoni-Gâtel, D.; Tardieux, I. CD11c- and CD11b-expressing mouse leukocytes transport single Toxoplasma gondii tachyzoites to the brain. Blood 2006, 107, 309–331. [Google Scholar] [CrossRef]
- Rishi, G.; Subramaniam, V.N. The liver in regulation of iron homeostasis. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Denkers, E.Y.; Gazzinelli, R.T. Regulation and function of T-cell-mediated immunity during Toxoplasma gondii infection. Clin. Microbiol. Rev. 1998, 11, 569–588. [Google Scholar] [CrossRef]
- Sibley, L.D.; Adams, L.B.; Fukutomi, Y.; Krahenbuhl, J.L. Tumor necrosis factor-alpha triggers antitoxoplasmal activity of IFN-gamma primed macrophages. J. Immunol. 1991, 147, 2340–2345. [Google Scholar] [PubMed]
- Langermans, J.A.; van der Hulst, M.E.; Nibbering, P.H.; van Furth, R. Endogenous tumor necrosis factor alpha is required for enhanced antimicrobial activity against Toxoplasma gondii and Listeria monocytogenes in recombinant gamma interferon-treated mice. Infect. Immun. 1992, 60, 5107–5112. [Google Scholar] [CrossRef] [PubMed]
- Liesenfeld, O.; Kosek, J.; Remington, J.S.; Suzuki, Y. Association of CD4+ T cell-dependent, interferon-gamma-mediated necrosis of the small intestine with genetic susceptibility of mice to peroral infection with Toxoplasma gondii. J. Exp. Med. 1996, 184, 597–607. [Google Scholar] [CrossRef]
- Liesenfeld, O.; Kang, H.; Park, D.; Nguyen, T.A.; Parkhe, C.V.; Watanabe, H.; Abo, T.; Sher, A.; Remington, J.S.; Suzuki, Y. TNF-alpha, nitric oxide and IFN-gamma are all critical for development of necrosis in the small intestine and early mortality in genetically susceptible mice infected perorally with Toxoplasma gondii. Parasite Immunol. 1999, 21, 365–376. [Google Scholar] [CrossRef]
- Okada, T.; Sawada, T.; Kubota, K. Deferoxamine enhances anti-proliferative effect of interferon-gamma against hepatocellular carcinoma cells. Cancer Lett. 2007, 248, 24–31. [Google Scholar] [CrossRef]
- Akira, S.; Taga, T.; Kishimoto, T. Interleukin-6 in biology and medicine. Adv. Immunol. 1993, 54, 1–78. [Google Scholar] [CrossRef]
- Suzuki, Y.; Rani, S.; Liesenfeld, O.; Kojima, T.; Lim, S.; Nguyen, T.A.; Dalrymple, S.A.; Murray, R.; Remington, J.S. Impaired resistance to the development of toxoplasmic encephalitis in interleukin-6-deficient mice. Infect. Immun. 1997, 65, 2339–2345. [Google Scholar] [CrossRef]
- Jebbari, H.; Roberts, C.W.; Ferguson, D.J.; Bluethmann, H.; Alexander, J. A protective role for IL-6 during early infection with Toxoplasma gondii. Parasite Immunol. 1998, 20, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Whitmarsh, R.J.; Gray, C.M.; Gregg, B.; Christian, D.A.; May, M.J.; Murray, P.J.; Hunter, C.A. A critical role for SOCS3 in innate resistance to Toxoplasma gondii. Cell Host Microbe 2011, 10, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Gazzinelli, R.T.; Wysocka, M.; Hieny, S.; Scharton-Kersten, T.; Cheever, A.; Kühn, R.; Müller, W.; Trinchieri, G.; Sher, A. In the absence of endogenous IL-10, mice acutely infected with Toxoplasma gondii succumb to a lethal immune response dependent on CD4+ T cells and accompanied by overproduction of IL-12, IFN-gamma and TNF-alpha. J. Immunol. 1996, 157, 798–805. [Google Scholar] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, M.C.; Coutinho, L.B.; Almeida, M.P.O.; Briceño, M.P.; Araujo, E.C.B.; Silva, N.M. The Availability of Iron Is Involved in the Murine Experimental Toxoplasma gondii Infection Outcome. Microorganisms 2020, 8, 560. https://doi.org/10.3390/microorganisms8040560
Oliveira MC, Coutinho LB, Almeida MPO, Briceño MP, Araujo ECB, Silva NM. The Availability of Iron Is Involved in the Murine Experimental Toxoplasma gondii Infection Outcome. Microorganisms. 2020; 8(4):560. https://doi.org/10.3390/microorganisms8040560
Chicago/Turabian StyleOliveira, Mário Cézar, Loyane Bertagnolli Coutinho, Marcos Paulo Oliveira Almeida, Marisol Pallete Briceño, Ester Cristina Borges Araujo, and Neide Maria Silva. 2020. "The Availability of Iron Is Involved in the Murine Experimental Toxoplasma gondii Infection Outcome" Microorganisms 8, no. 4: 560. https://doi.org/10.3390/microorganisms8040560
APA StyleOliveira, M. C., Coutinho, L. B., Almeida, M. P. O., Briceño, M. P., Araujo, E. C. B., & Silva, N. M. (2020). The Availability of Iron Is Involved in the Murine Experimental Toxoplasma gondii Infection Outcome. Microorganisms, 8(4), 560. https://doi.org/10.3390/microorganisms8040560