Lysogeny in Streptococcus pneumoniae


Abstract
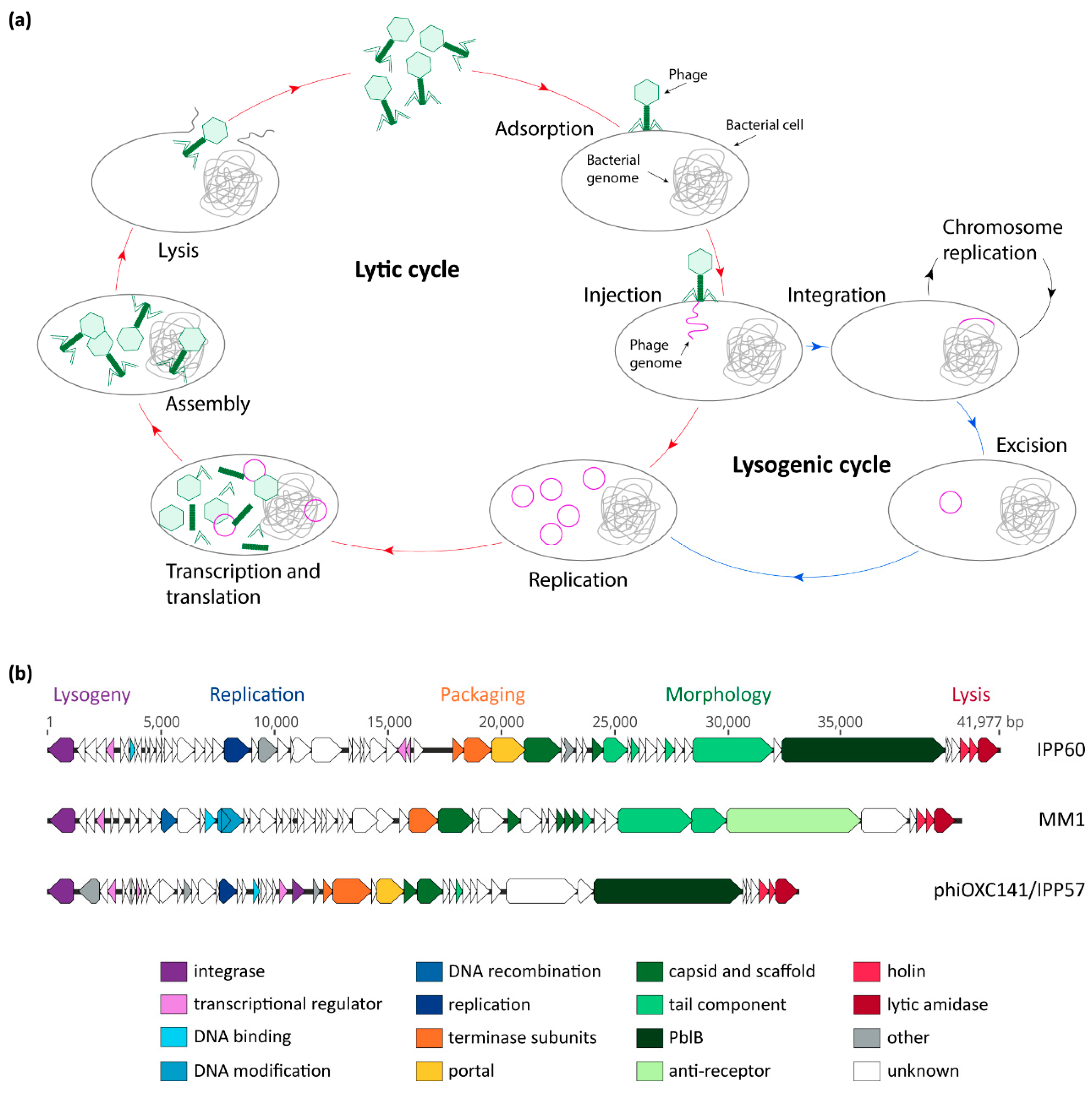
1. Introduction
2. Pneumococcal Bacteriophage Genetic Modules
3. Phage Groups and Chromosomal Integration Loci
4. Polylysogeny
5. The Phage and the Host: Contribution to Virulence and Colonization
6. Phages, Biofilms, and Natural Competence for Transformation
7. Phage-Related Chromosomal Islands
8. Perspectives
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Phage Name | Phage Accession Number | Genome Accession Number |
|---|---|---|
| IPP12 | KY065454.1 | NC_012467.1 |
| IPP39 | KY065479.1 | AGPM01000004.1 |
| phiSpn_200/IPP42 | KY065482.1 | NC_014494.1 |
| IPP43 | KY065483.1 | NZ_UHGQ01000001.1 |
| IPP45 | KY065485.1 | CP002176.1 |
| IPP46 | KY065486.1 | CP002176.1 |
| IPP48 | KY065487.1 | CP003357.2 |
| IPP51 | KY065489.1 | AGPN01000004.1 |
| IPP53 | KY065491.1 | AGOL01000005.1 |
| phiOXC141/IPP57 | KY065494.1 | NC_017592.1 |
| IPP58 | KY065495.1 | AGQI01000002.1 |
| IPP60 | KY065496.1 | NZ_FYOS01000005.1 |
| IPP67 | KY065503.1 | NZ_ABWQ01000001.1 |
| MM1 | NC_003050.2 | FM211187.1 |
| phi34117 | FR671407.1 | Not available |
| phi8140 | FR671410.1 | Not available |
| phiNTCC11888 | Used genome | NZ_UHFY01000001.1 |
| phiNTCC13735 | Used genome | NZ_UHGD01000001.1 |
| phiSMRU1311 | Used genome | NZ_CRHW01000001.1 |
| phiSMRU2522 | Used genome | NZ_CLJV01000005.1 |
| phiSMRU3096 | Used genome | NZ_CMLU01000003.1 |
| phiSPNH_2 | Used genome | CP000936.1 |
| phiUoS0268 | Used genome | NZ_FYHY01000007.1 |
| phiUoS4388 | Used genome | NZ_FYVE01000006.1 |
| SPN195_1 | Used genome | ABGE01000002.1 |
| SPN195_2 | Used genome | ABGE01000013.1 |
References
- Suttle, C.A. Viruses in the sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Dion, M.B.; Oechslin, F.; Moineau, S. Phage diversity, genomics and phylogeny. Nat. Rev. Microbiol. 2020, 18, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Keen, E.C. A century of phage research: Bacteriophages and the shaping of modern biology. Bioessays 2015, 37, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Salmond, G.P.; Fineran, P.C. A century of the phage: Past, present and future. Nat. Rev. Microbiol. 2015, 13, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Fiers, W.; Contreras, R.; Duerinck, F.; Haegeman, G.; Iserentant, D.; Merregaert, J.; Min Jou, W.; Molemans, F.; Raeymaekers, A.; Van den Berghe, A.; et al. Complete nucleotide sequence of bacteriophage MS2 RNA: Primary and secondary structure of the replicase gene. Nature 1976, 260, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Sanger, F.; Air, G.M.; Barrell, B.G.; Brown, N.L.; Coulson, A.R.; Fiddes, C.A.; Hutchison, C.A.; Slocombe, P.M.; Smith, M. Nucleotide sequence of bacteriophage phi X174 DNA. Nature 1977, 265, 687–695. [Google Scholar] [CrossRef]
- Penadés, J.R.; Christie, G.E. The Phage-Inducible Chromosomal Islands: A Family of Highly Evolved Molecular Parasites. Annu. Rev. Virol. 2015, 2, 181–201. [Google Scholar] [CrossRef]
- Feiner, R.; Argov, T.; Rabinovich, L.; Sigal, N.; Borovok, I.; Herskovits, A.A. A new perspective on lysogeny: Prophages as active regulatory switches of bacteria. Nat. Rev. Microbiol. 2015, 13, 641–650. [Google Scholar] [CrossRef]
- Freeman, V.J. Studies on the virulence of bacteriophage-infected strains of Corynebacterium diphtheriae. J. Bacteriol. 1951, 61, 675–688. [Google Scholar] [CrossRef]
- Plunkett, G., 3rd; Rose, D.J.; Durfee, T.J.; Blattner, F.R. Sequence of Shiga toxin 2 phage 933W from Escherichia coli O157:H7: Shiga toxin as a phage late-gene product. J. Bacteriol. 1999, 181, 1767–1778. [Google Scholar] [CrossRef]
- Waldor, M.K.; Mekalanos, J.J. Lysogenic conversion by a filamentous phage encoding cholera toxin. Science 1996, 272, 1910–1914. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, M.; Lain, R.; Tomasz, A. “Diplophage”: A bacteriophage of Diplococcus pneumoniae. Virology 1975, 63, 577–582. [Google Scholar] [CrossRef]
- Tiraby, J.G.; Tiraby, E.; Fox, M.S. Pneumococcal bacteriophages. Virology 1975, 68, 566–569. [Google Scholar] [CrossRef]
- Twort, F.W. An investigation on the nature of ultra-microscopic viruses. Lancet 1915, 2, 1241–1243. [Google Scholar] [CrossRef]
- d’Hérelle, F. Sur un microbe invisible antagonistic des bacilles dysenterique. Acad. des Sci. de Paris 1917, 165, 373–375. [Google Scholar]
- Bernheimer, H.P. Lysogeny in pneumococci freshly isolated from man. Science 1977, 195, 66–68. [Google Scholar] [CrossRef]
- Bernheimer, H.P. Lysogenic pneumococci and their bacteriophages. J. Bacteriol. 1979, 138, 618–624. [Google Scholar] [CrossRef]
- López, R.; García, E. Recent trends on the molecular biology of pneumococcal capsules, lytic enzymes, and bacteriophage. FEMS Microbiol. Rev. 2004, 28, 553–580. [Google Scholar] [CrossRef]
- Ramirez, M.; Severina, E.; Tomasz, A. A high incidence of prophage carriage among natural isolates of Streptococcus pneumoniae. J. Bacteriol. 1999, 181, 3618–3625. [Google Scholar] [CrossRef]
- Brueggemann, A.B.; Harrold, C.L.; Rezaei Javan, R.; van Tonder, A.J.; McDonnell, A.J.; Edwards, B.A. Pneumococcal prophages are diverse, but not without structure or history. Sci. Rep. 2017, 7, 42976. [Google Scholar] [CrossRef]
- Croucher, N.J.; Coupland, P.G.; Stevenson, A.E.; Callendrello, A.; Bentley, S.D.; Hanage, W.P. Diversification of bacterial genome content through distinct mechanisms over different timescales. Nat. Commun. 2014, 5, 5471. [Google Scholar] [CrossRef] [PubMed]
- Croucher, N.J.; Mostowy, R.; Wymant, C.; Turner, P.; Bentley, S.D.; Fraser, C. Horizontal DNA Transfer Mechanisms of Bacteria as Weapons of Intragenomic Conflict. PLoS Biol. 2016, 14, e1002394. [Google Scholar] [CrossRef] [PubMed]
- Obregón, V.; García, J.L.; García, E.; Lopez, R.; García, P. Genome organization and molecular analysis of the temperate bacteriophage MM1 of Streptococcus pneumoniae. J. Bacteriol. 2003, 185, 2362–2368. [Google Scholar] [CrossRef] [PubMed]
- Gindreau, E.; Lopez, R.; Garcia, P. MM1, a temperate bacteriophage of the type 23F Spanish/USA multiresistant epidemic clone of Streptococcus pneumoniae: Structural analysis of the site-specific integration system. J. Virol. 2000, 74, 7803–7813. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Obregón, V.; García, P.; López, R.; García, J.L. Molecular and biochemical analysis of the system regulating the lytic/lysogenic cycle in the pneumococcal temperate phage MM1. FEMS Microbiol. Lett. 2003, 222, 193–197. [Google Scholar] [CrossRef]
- Romero, P.; Croucher, N.J.; Hiller, N.L.; Hu, F.Z.; Ehrlich, G.D.; Bentley, S.D.; García, E.; Mitchell, T.J. Comparative genomic analysis of ten Streptococcus pneumoniae temperate bacteriophages. J. Bacteriol. 2009, 191, 4854–4862. [Google Scholar] [CrossRef]
- Camilli, R.; Bonnal, R.J.; Del Grosso, M.; Iacono, M.; Corti, G.; Rizzi, E.; Marchetti, M.; Mulas, L.; Iannelli, F.; Superti, F.; et al. Complete genome sequence of a serotype 11A, ST62 Streptococcus pneumoniae invasive isolate. BMC Microbiol. 2011, 11, 25. [Google Scholar] [CrossRef]
- Casjens, S.R.; Hendrix, R.W. Bacteriophage lambda: Early pioneer and still relevant. Virology 2015, 479–480, 310–330. [Google Scholar] [CrossRef]
- Oppenheim, A.B.; Kobiler, O.; Stavans, J.; Court, D.L.; Adhya, S. Switches in bacteriophage lambda development. Annu. Rev. Genet. 2005, 39, 409–429. [Google Scholar] [CrossRef]
- Erill, I.; Campoy, S.; Barbe, J. Aeons of distress: An evolutionary perspective on the bacterial SOS response. FEMS Microbiol. Rev. 2007, 31, 637–656. [Google Scholar] [CrossRef]
- Prudhomme, M.; Attaiech, L.; Sanchez, G.; Martin, B.; Claverys, J.P. Antibiotic stress induces genetic transformability in the human pathogen Streptococcus pneumoniae. Science 2006, 313, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Furi, L.; Crawford, L.A.; Rangel-Pineros, G.; Manso, A.S.; De Ste Croix, M.; Haigh, R.D.; Kwun, M.J.; Engelsen Fjelland, K.; Gilfillan, G.D.; Bentley, S.D.; et al. Methylation Warfare: Interaction of Pneumococcal Bacteriophages with Their Host. J. Bacteriol. 2019, 201, e00370-19. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.; García, P.; Castanié, M.P.; Claverys, J.P. The recA gene of Streptococcus pneumoniae is part of a competence-induced operon and controls lysogenic induction. Mol. Microbiol. 1995, 15, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Tunjungputri, R.N.; Mobegi, F.M.; Cremers, A.J.; van der Gaast-de Jongh, C.E.; Ferwerda, G.; Meis, J.F.; Roeleveld, N.; Bentley, S.D.; Pastura, A.S.; van Hijum, S.A.; et al. Phage-Derived Protein Induces Increased Platelet Activation and Is Associated with Mortality in Patients with Invasive Pneumococcal Disease. mBio 2017, 8, e01984-16. [Google Scholar] [CrossRef] [PubMed]
- López, E.; Domenech, A.; Ferrándiz, M.J.; Frias, M.J.; Ardanuy, C.; Ramirez, M.; García, E.; Liñares, J.; de la Campa, A.G. Induction of prophages by fluoroquinolones in Streptococcus pneumoniae: Implications for emergence of resistance in genetically-related clones. PLoS ONE 2014, 9, e94358. [Google Scholar]
- Rao, V.B.; Feiss, M. The bacteriophage DNA packaging motor. Annu. Rev. Genet. 2008, 42, 647–681. [Google Scholar] [CrossRef]
- García, P.; Martín, A.C.; López, R. Bacteriophages of Streptococcus pneumoniae: A molecular approach. Microb. Drug Resist. 1997, 3, 165–176. [Google Scholar] [CrossRef]
- Romero, P.; García, E.; Mitchell, T.J. Development of a prophage typing system and analysis of prophage carriage in Streptococcus pneumoniae. Appl. Environ. Microbiol. 2009, 75, 1642–1649. [Google Scholar] [CrossRef]
- Bensing, B.A.; Siboo, I.R.; Sullam, P.M. Proteins PblA and PblB of Streptococcus mitis, which promote binding to human platelets, are encoded within a lysogenic bacteriophage. Infect. Immun. 2001, 69, 6186–6192. [Google Scholar] [CrossRef]
- López, R.; García, E.; García, P.; Ronda, C.; Tomasz, A. Choline-containing bacteriophage receptors in Streptococcus pneumoniae. J. Bacteriol. 1982, 151, 1581–1590. [Google Scholar] [CrossRef]
- Mitchell, J.; Siboo, I.R.; Takamatsu, D.; Chambers, H.F.; Sullam, P.M. Mechanism of cell surface expression of the Streptococcus mitis platelet binding proteins PblA and PblB. Mol. Microbiol. 2007, 64, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Bernheimer, H.P.; Tiraby, J.G. Inhibition of phage infection by pneumococcus capsule. Virology 1976, 73, 308–309. [Google Scholar] [CrossRef]
- Leprohon, P.; Gingras, H.; Ouennane, S.; Moineau, S.; Ouellette, M. A genomic approach to understand interactions between Streptococcus pneumoniae and its bacteriophages. BMC Genom. 2015, 16, 972. [Google Scholar] [CrossRef] [PubMed]
- Manso, A.S.; Chai, M.H.; Atack, J.M.; Furi, L.; De Ste Croix, M.; Haigh, R.; Trappetti, C.; Ogunniyi, A.D.; Shewell, L.K.; Boitano, M.; et al. A random six-phase switch regulates pneumococcal virulence via global epigenetic changes. Nat. Commun. 2014, 5, 5055. [Google Scholar] [CrossRef]
- Cundell, D.R.; Weiser, J.N.; Shen, J.; Young, A.; Tuomanen, E.I. Relationship between colonial morphology and adherence of Streptococcus pneumoniae. Infect. Immun. 1995, 63, 757–761. [Google Scholar] [CrossRef]
- Weiser, J.N.; Austrian, R.; Sreenivasan, P.K.; Masure, H.R. Phase variation in pneumococcal opacity: Relationship between colonial morphology and nasopharyngeal colonization. Infect. Immun. 1994, 62, 2582–2589. [Google Scholar] [CrossRef]
- Wang, I.N.; Smith, D.L.; Young, R. Holins: The protein clocks of bacteriophage infections. Annu. Rev. Microbiol. 2000, 54, 799–825. [Google Scholar] [CrossRef]
- Morales, M.; Martín-Galiano, A.J.; Domenech, M.; García, E. Insights into the Evolutionary Relationships of LytA Autolysin and Ply Pneumolysin-Like Genes in Streptococcus pneumoniae and Related Streptococci. Genome Biol. Evol. 2015, 7, 2747–2761. [Google Scholar] [CrossRef]
- Romero, A.; López, R.; García, P. Sequence of the Streptococcus pneumoniae bacteriophage HB-3 amidase reveals high homology with the major host autolysin. J. Bacteriol. 1990, 172, 5064–5070. [Google Scholar] [CrossRef]
- García, P.; García, E.; Ronda, C.; López, R.; Tomasz, A. A phage-associated murein hydrolase in Streptococcus pneumoniae infected with bacteriophage Dp-1. J. Gen. Microbiol. 1983, 129, 489–497. [Google Scholar] [CrossRef]
- Frias, M.J.; Melo-Cristino, J.; Ramirez, M. The autolysin LytA contributes to efficient bacteriophage progeny release in Streptococcus pneumoniae. J. Bacteriol. 2009, 191, 5428–5440. [Google Scholar] [CrossRef] [PubMed]
- Browall, S.; Norman, M.; Tangrot, J.; Galanis, I.; Sjostrom, K.; Dagerhamn, J.; Hellberg, C.; Pathak, A.; Spadafina, T.; Sandgren, A.; et al. Intraclonal variations among Streptococcus pneumoniae isolates influence the likelihood of invasive disease in children. J. Infect. Dis. 2014, 209, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Croucher, N.J.; Harris, S.R.; Fraser, C.; Quail, M.A.; Burton, J.; van der Linden, M.; McGee, L.; von Gottberg, A.; Song, J.H.; Ko, K.S.; et al. Rapid pneumococcal evolution in response to clinical interventions. Science 2011, 331, 430–434. [Google Scholar] [CrossRef]
- Botstein, D. A theory of modular evolution for bacteriophages. Ann. N. Y. Acad. Sci. 1980, 354, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.M. Preferential orientation of natural lambdoid prophages and bacterial chromosome organization. Theor. Popul. Biol. 2002, 61, 503–507. [Google Scholar] [CrossRef]
- Canchaya, C.; Fournous, G.; Brussow, H. The impact of prophages on bacterial chromosomes. Mol. Microbiol. 2004, 53, 9–18. [Google Scholar] [CrossRef]
- Bobay, L.M.; Rocha, E.P.; Touchon, M. The adaptation of temperate bacteriophages to their host genomes. Mol. Biol. Evol. 2013, 30, 737–751. [Google Scholar] [CrossRef]
- Groth, A.C.; Calos, M.P. Phage integrases: Biology and applications. J. Mol. Biol. 2004, 335, 667–678. [Google Scholar] [CrossRef]
- Lewis, J.A.; Hatfull, G.F. Control of directionality in integrase-mediated recombination: Examination of recombination directionality factors (RDFs) including Xis and Cox proteins. Nucleic Acids Res. 2001, 29, 2205–2216. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- DeBardeleben, H.K.; Lysenko, E.S.; Dalia, A.B.; Weiser, J.N. Tolerance of a phage element by Streptococcus pneumoniae leads to a fitness defect during colonization. J. Bacteriol. 2014, 196, 2670–2680. [Google Scholar] [CrossRef] [PubMed]
- Muschiol, S.; Erlendsson, S.; Aschtgen, M.S.; Oliveira, V.; Schmieder, P.; de Lichtenberg, C.; Teilum, K.; Boesen, T.; Akbey, U.; Henriques-Normark, B. Structure of the competence pilus major pilin ComGC in Streptococcus pneumoniae. J. Biol. Chem. 2017, 292, 14134–14146. [Google Scholar] [CrossRef]
- Càmara, J.; Cubero, M.; Martín-Galiano, A.J.; García, E.; Grau, I.; Nielsen, J.B.; Worning, P.; Tubau, F.; Pallarés, R.; Domínguez, M.A.; et al. Evolution of the beta-lactam-resistant Streptococcus pneumoniae PMEN3 clone over a 30 year period in Barcelona, Spain. J. Antimicrob. Chemother. 2018, 73, 2941–2951. [Google Scholar] [CrossRef]
- Bondy-Denomy, J.; Qian, J.; Westra, E.R.; Buckling, A.; Guttman, D.S.; Davidson, A.R.; Maxwell, K.L. Prophages mediate defense against phage infection through diverse mechanisms. ISME J. 2016, 10, 2854–2866. [Google Scholar] [CrossRef]
- Kameyama, L.; Fernández, L.; Calderón, J.; Ortiz-Rojas, A.; Patterson, T.A. Characterization of wild lambdoid bacteriophages: Detection of a wide distribution of phage immunity groups and identification of a nus-dependent, nonlambdoid phage group. Virology 1999, 263, 100–111. [Google Scholar] [CrossRef][Green Version]
- Owen, S.V.; Canals, R.; Wenner, N.; Hammarlof, D.L.; Kroger, C.; Hinton, J.C.D. A window into lysogeny: Revealing temperate phage biology with transcriptomics. Microb. Genom. 2020, 6, e000330. [Google Scholar] [CrossRef]
- Ricker, N.; Qian, H.; Fulthorpe, R.R. The limitations of draft assemblies for understanding prokaryotic adaptation and evolution. Genomics 2012, 100, 167–175. [Google Scholar] [CrossRef]
- Jorgensen, T.S.; Kiil, A.S.; Hansen, M.A.; Sorensen, S.J.; Hansen, L.H. Current strategies for mobilome research. Front. Microbiol. 2014, 5, 750. [Google Scholar]
- Carrolo, M.; Frias, M.J.; Pinto, F.R.; Melo-Cristino, J.; Ramirez, M. Prophage spontaneous activation promotes DNA release enhancing biofilm formation in Streptococcus pneumoniae. PLoS ONE 2010, 5, e15678. [Google Scholar] [CrossRef] [PubMed]
- Rezaei Javan, R.; Ramos-Sevillano, E.; Akter, A.; Brown, J.; Brueggemann, A.B. Prophages and satellite prophages are widespread in Streptococcus and may play a role in pneumococcal pathogenesis. Nat. Commun. 2019, 10, 4852. [Google Scholar] [CrossRef] [PubMed]
- Croucher, N.J.; Walker, D.; Romero, P.; Lennard, N.; Paterson, G.K.; Bason, N.C.; Mitchell, A.M.; Quail, M.A.; Andrew, P.W.; Parkhill, J.; et al. Role of conjugative elements in the evolution of the multidrug-resistant pandemic clone Streptococcus pneumoniae Spain23F ST81. J. Bacteriol. 2009, 191, 1480–1489. [Google Scholar] [CrossRef] [PubMed]
- Hiller, N.L.; Eutsey, R.A.; Powell, E.; Earl, J.P.; Janto, B.; Martin, D.P.; Dawid, S.; Ahmed, A.; Longwell, M.J.; Dahlgren, M.E.; et al. Differences in genotype and virulence among four multidrug-resistant Streptococcus pneumoniae isolates belonging to the PMEN1 clone. PLoS ONE 2011, 6, e28850. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.M.; Stroeher, U.H.; Ogunniyi, A.D.; Smith-Vaughan, H.C.; Leach, A.J.; Paton, J.C. A variable region within the genome of Streptococcus pneumoniae contributes to strain-strain variation in virulence. PLoS ONE 2011, 6, e19650. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.M.; Trappetti, C.; Mahdi, L.K.; Wang, H.; McAllister, L.J.; Scalvini, A.; Paton, A.W.; Paton, J.C. The Variable Region of Pneumococcal Pathogenicity Island 1 Is Responsible for Unusually High Virulence of a Serotype 1 Isolate. Infect. Immun. 2016, 84, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.M.; Loman, N.J.; Ebruke, C.; Musher, D.M.; Adegbola, R.A.; Pallen, M.J.; Weinstock, G.M.; Antonio, M. Genome analysis of a highly virulent serotype 1 strain of Streptococcus pneumoniae from West Africa. PLoS ONE 2012, 7, e26742. [Google Scholar] [CrossRef]
- Bensing, B.A.; Rubens, C.E.; Sullam, P.M. Genetic loci of Streptococcus mitis that mediate binding to human platelets. Infect. Immun. 2001, 69, 1373–1380. [Google Scholar] [CrossRef]
- Hsieh, Y.C.; Lin, T.L.; Lin, C.M.; Wang, J.T. Identification of PblB mediating galactose-specific adhesion in a successful Streptococcus pneumoniae clone. Sci. Rep. 2015, 5, 12265. [Google Scholar] [CrossRef]
- Loeffler, J.M.; Fischetti, V.A. Lysogeny of Streptococcus pneumoniae with MM1 phage: Improved adherence and other phenotypic changes. Infect. Immun. 2006, 74, 4486–4495. [Google Scholar] [CrossRef]
- Hiller, N.L.; Sá-Leão, R. Puzzling Over the Pneumococcal Pangenome. Front. Microbiol. 2018, 9, 2580. [Google Scholar] [CrossRef] [PubMed]
- Cowley, L.A.; Petersen, F.C.; Junges, R.; Jimson, D.J.M.; Morrison, D.A.; Hanage, W.P. Evolution via recombination: Cell-to-cell contact facilitates larger recombination events in Streptococcus pneumoniae. PLoS Genet. 2018, 14, e1007410. [Google Scholar] [CrossRef] [PubMed]
- Lees, J.A.; Croucher, N.J.; Goldblatt, D.; Nosten, F.; Parkhill, J.; Turner, C.; Turner, P.; Bentley, S.D. Genome-wide identification of lineage and locus specific variation associated with pneumococcal carriage duration. eLife 2017, 6, e26255. [Google Scholar] [CrossRef]
- Novick, R.P.; Christie, G.E.; Penades, J.R. The phage-related chromosomal islands of Gram-positive bacteria. Nat. Rev. Microbiol. 2010, 8, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Rubio, R.; Quiles-Puchalt, N.; Martí, M.; Humphrey, S.; Ram, G.; Smyth, D.; Chen, J.; Novick, R.P.; Penadés, J.R. Phage-inducible islands in the Gram-positive cocci. ISME J. 2017, 11, 1029–1042. [Google Scholar] [CrossRef]
- Tormo-Mas, M.A.; Mir, I.; Shrestha, A.; Tallent, S.M.; Campoy, S.; Lasa, I.; Barbe, J.; Novick, R.P.; Christie, G.E.; Penades, J.R. Moonlighting bacteriophage proteins derepress staphylococcal pathogenicity islands. Nature 2010, 465, 779–782. [Google Scholar] [CrossRef]
- Deshpande, N.P.; Riordan, S.M.; Castano-Rodriguez, N.; Wilkins, M.R.; Kaakoush, N.O. Signatures within the esophageal microbiome are associated with host genetics, age, and disease. Microbiome 2018, 6, 227. [Google Scholar] [CrossRef] [PubMed]
- Willner, D.; Furlan, M.; Haynes, M.; Schmieder, R.; Angly, F.E.; Silva, J.; Tammadoni, S.; Nosrat, B.; Conrad, D.; Rohwer, F. Metagenomic analysis of respiratory tract DNA viral communities in cystic fibrosis and non-cystic fibrosis individuals. PLoS ONE 2009, 4, e7370. [Google Scholar] [CrossRef] [PubMed]
- Duerkop, B.A. Bacteriophages shift the focus of the mammalian microbiota. PLoS Pathog. 2018, 14, e1007310. [Google Scholar] [CrossRef]
- Keen, E.C.; Dantas, G. Close Encounters of Three Kinds: Bacteriophages, Commensal Bacteria, and Host Immunity. Trends Microbiol. 2018, 26, 943–954. [Google Scholar] [CrossRef]
- Man, W.H.; de Steenhuijsen Piters, W.A.; Bogaert, D. The microbiota of the respiratory tract: Gatekeeper to respiratory health. Nat. Rev. Microbiol. 2017, 15, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Garriss, G.; Nannapaneni, P.; Simoes, A.S.; Browall, S.; Subramanian, K.; Sa-Leao, R.; Goossens, H.; de Lencastre, H.; Henriques-Normark, B. Genomic Characterization of the Emerging Pathogen Streptococcus pseudopneumoniae. mBio 2019, 10, e01286-19. [Google Scholar] [CrossRef] [PubMed]
- Bikard, D.; Hatoum-Aslan, A.; Mucida, D.; Marraffini, L.A. CRISPR interference can prevent natural transformation and virulence acquisition during in vivo bacterial infection. Cell Host Microbe 2012, 12, 177–186. [Google Scholar] [CrossRef]
- Pride, D.T.; Sun, C.L.; Salzman, J.; Rao, N.; Loomer, P.; Armitage, G.C.; Banfield, J.F.; Relman, D.A. Analysis of streptococcal CRISPRs from human saliva reveals substantial sequence diversity within and between subjects over time. Genome Res. 2011, 21, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Pride, D.T.; Salzman, J.; Relman, D.A. Comparisons of clustered regularly interspaced short palindromic repeats and viromes in human saliva reveal bacterial adaptations to salivary viruses. Environ. Microbiol. 2012, 14, 2564–2576. [Google Scholar] [CrossRef]
- Park, B.; Nizet, V.; Liu, G.Y. Role of Staphylococcus aureus catalase in niche competition against Streptococcus pneumoniae. J. Bacteriol. 2008, 190, 2275–2278. [Google Scholar] [CrossRef]
- Regev-Yochay, G.; Trzcinski, K.; Thompson, C.M.; Malley, R.; Lipsitch, M. Interference between Streptococcus pneumoniae and Staphylococcus aureus: In vitro hydrogen peroxide-mediated killing by Streptococcus pneumoniae. J. Bacteriol. 2006, 188, 4996–5001. [Google Scholar] [CrossRef]
- Selva, L.; Viana, D.; Regev-Yochay, G.; Trzcinski, K.; Corpa, J.M.; Lasa, I.; Novick, R.P.; Penadés, J.R. Killing niche competitors by remote-control bacteriophage induction. Proc. Natl. Acad. Sci. USA 2009, 106, 1234–1238. [Google Scholar] [CrossRef]
- Chen, Y.; Golding, I.; Sawai, S.; Guo, L.; Cox, E.C. Population fitness and the regulation of Escherichia coli genes by bacterial viruses. PLoS Biol. 2005, 3, e229. [Google Scholar] [CrossRef]
- Hernandez-Doria, J.D.; Sperandio, V. Bacteriophage Transcription Factor Cro Regulates Virulence Gene Expression in Enterohemorrhagic Escherichia coli. Cell Host Microbe 2018, 23, 607–617. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garriss, G.; Henriques-Normark, B. Lysogeny in Streptococcus pneumoniae. Microorganisms 2020, 8, 1546. https://doi.org/10.3390/microorganisms8101546
Garriss G, Henriques-Normark B. Lysogeny in Streptococcus pneumoniae. Microorganisms. 2020; 8(10):1546. https://doi.org/10.3390/microorganisms8101546
Chicago/Turabian StyleGarriss, Geneviève, and Birgitta Henriques-Normark. 2020. "Lysogeny in Streptococcus pneumoniae" Microorganisms 8, no. 10: 1546. https://doi.org/10.3390/microorganisms8101546
APA StyleGarriss, G., & Henriques-Normark, B. (2020). Lysogeny in Streptococcus pneumoniae. Microorganisms, 8(10), 1546. https://doi.org/10.3390/microorganisms8101546