

Biological Characteristics and Fungicide Screening of Bipolaris oryzae Causing Leaf Spot on Banana in China


Abstract
1. Introduction
2. Materials and Methods
2.1. Pathogenic Fungal Strain, Inoculum Preparation, and Fungicides
2.2. The Effect of the Medium on the Mycelial Growth and Sporulation
2.3. The Effect of the Carbon and Nitrogen Sources on the Mycelial Growth and Sporulation
2.4. The Effect of Temperature on the Mycelial Growth and Sporulation
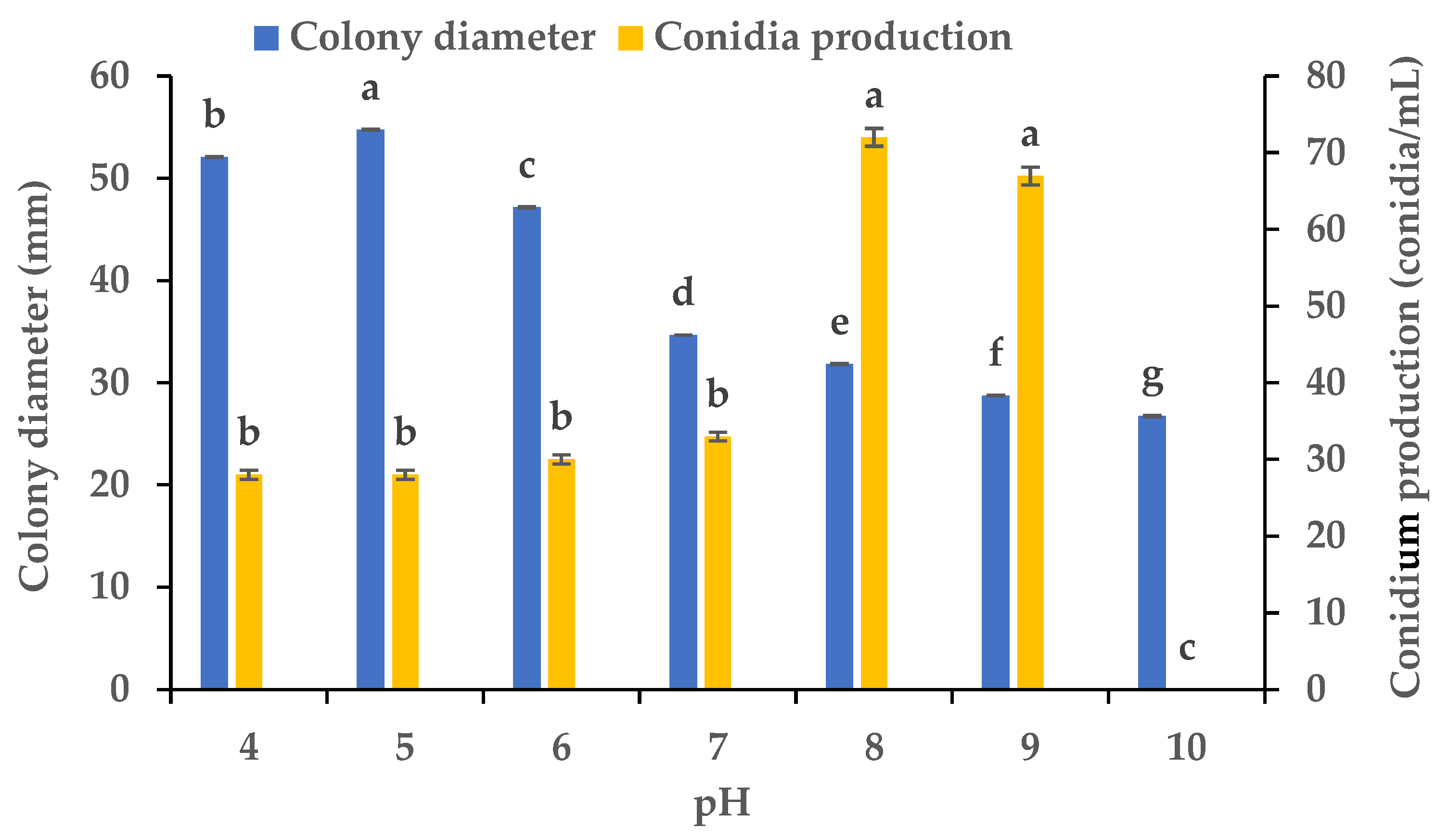
2.5. The Effect of pH on the Mycelial Growth and Sporulation
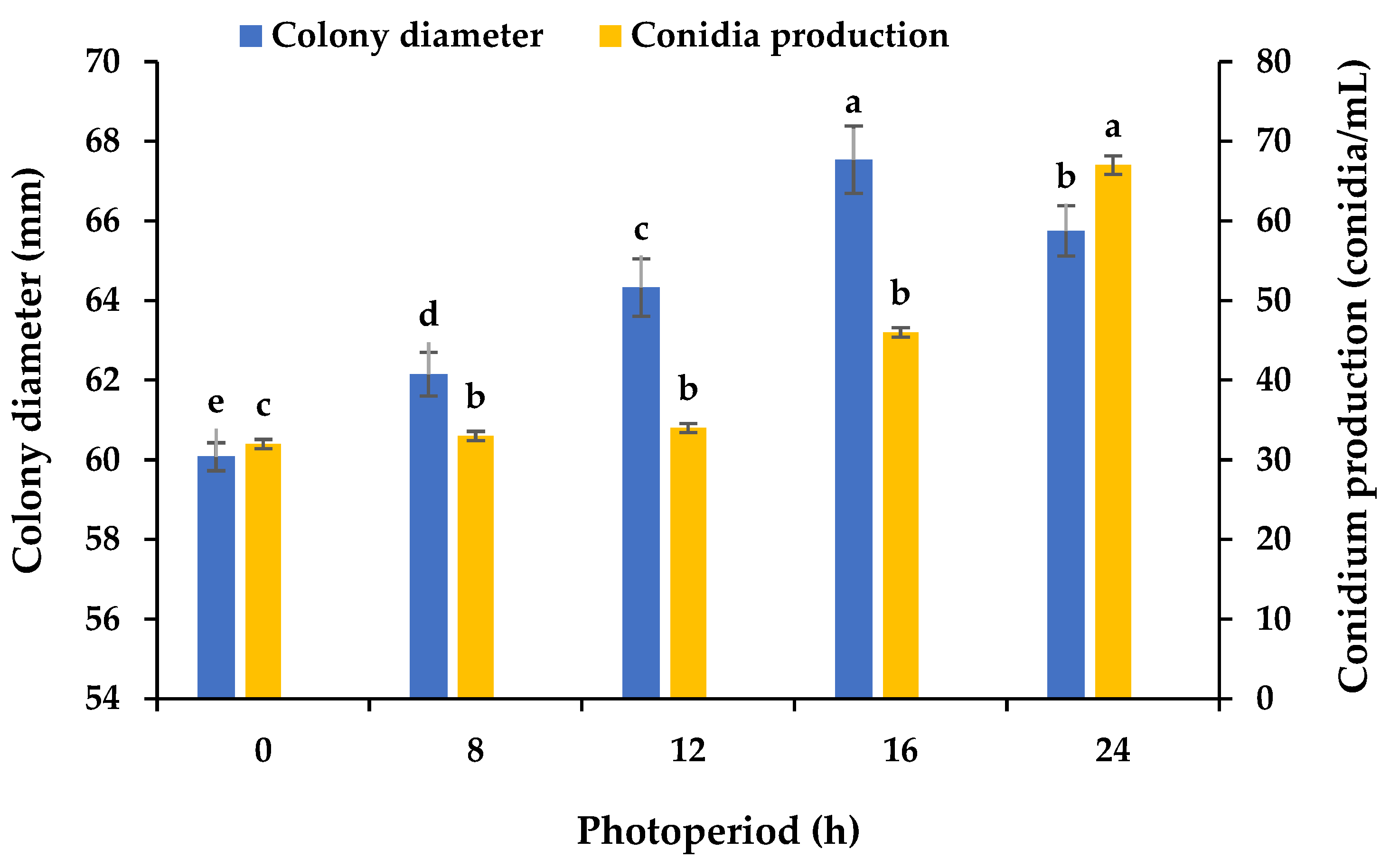
2.6. The Effect of Photoperiod on the Mycelial Growth and Sporulation
2.7. Determination of the Mycelial Lethal Temperature
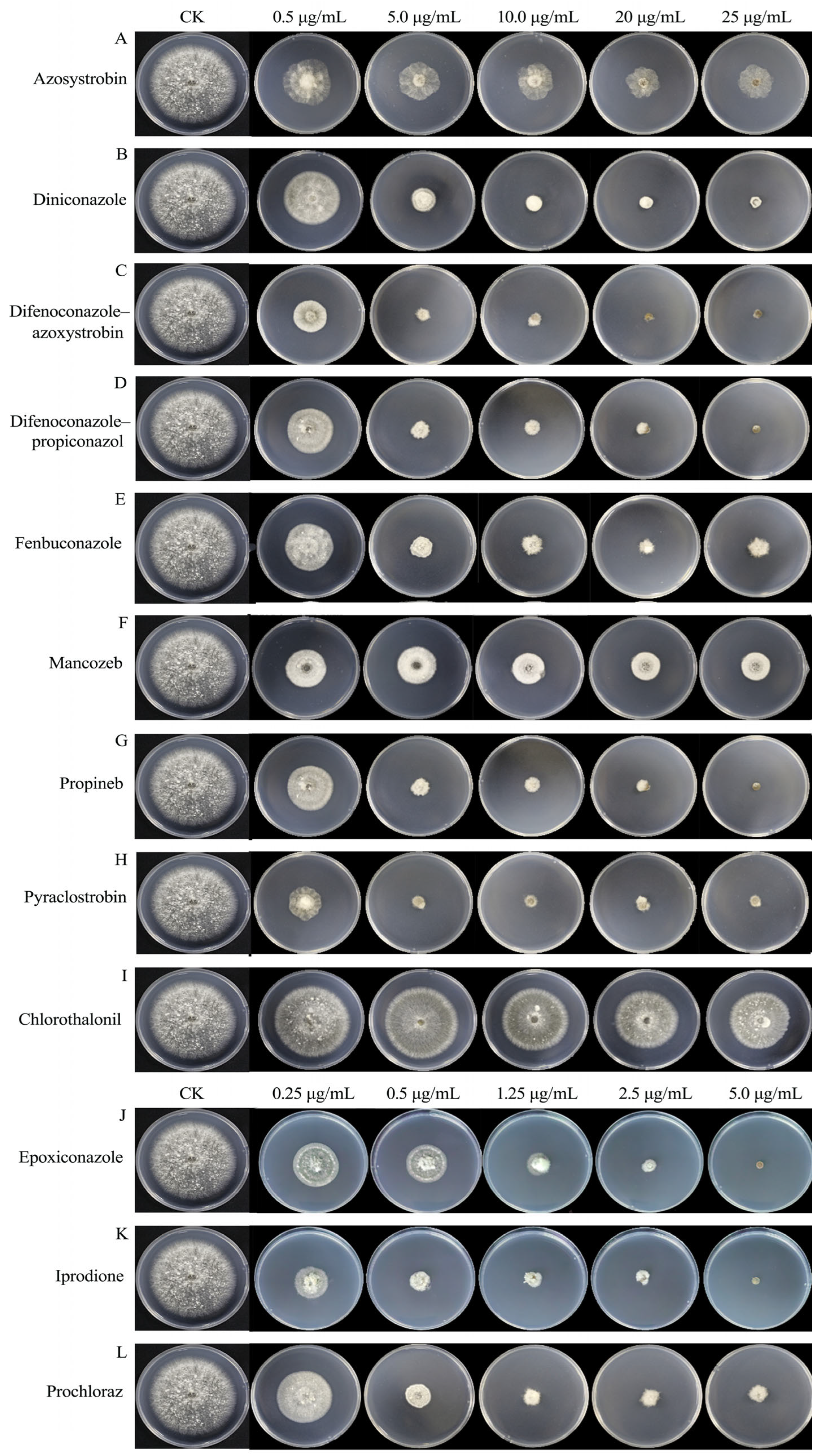
2.8. Screening of Indoor Fungicides
2.9. Control Efficacy of Fungicides
2.10. Statistical Analysis
3. Results
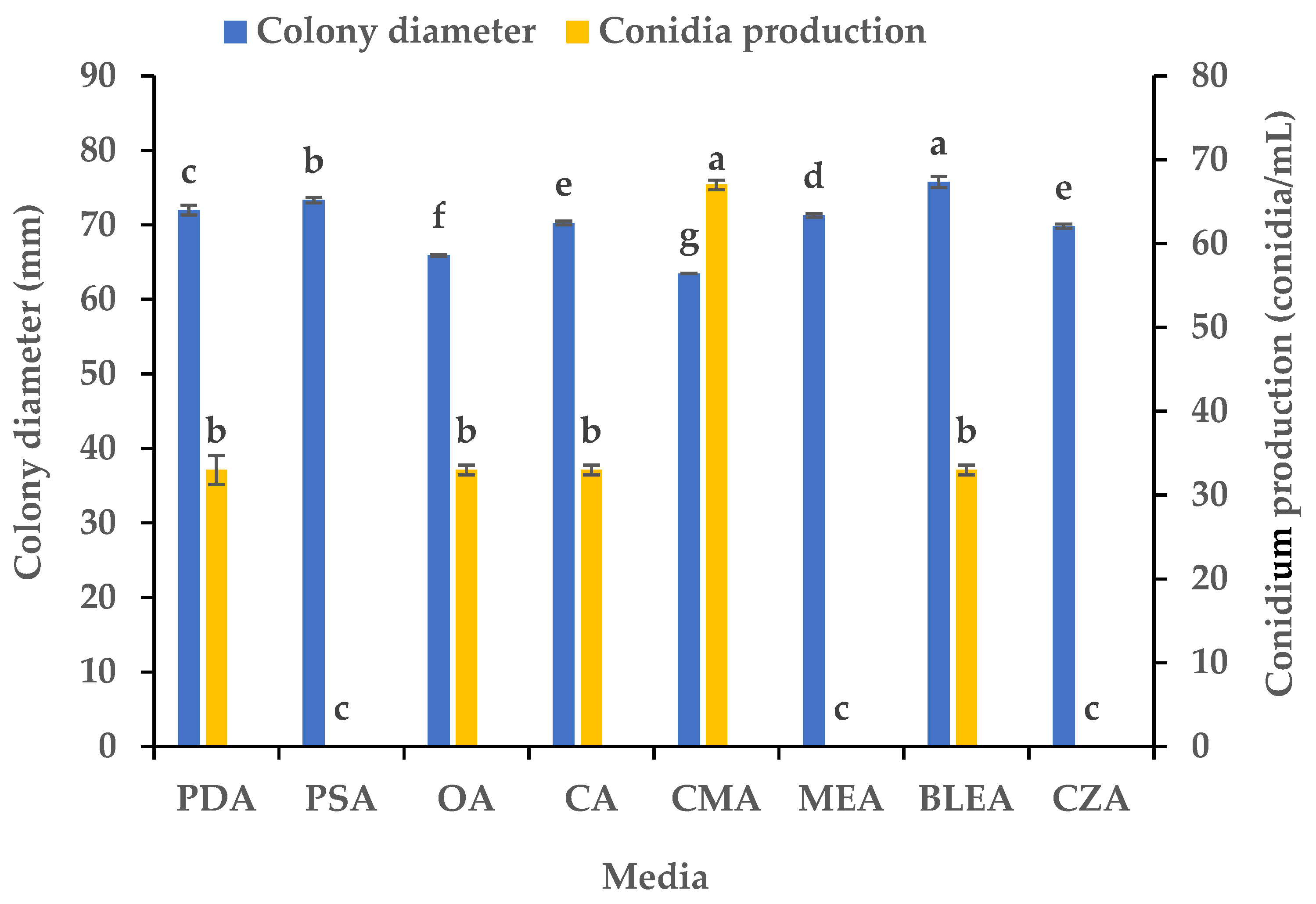
3.1. Effects of the Media on the Mycelial Growth and Sporulation
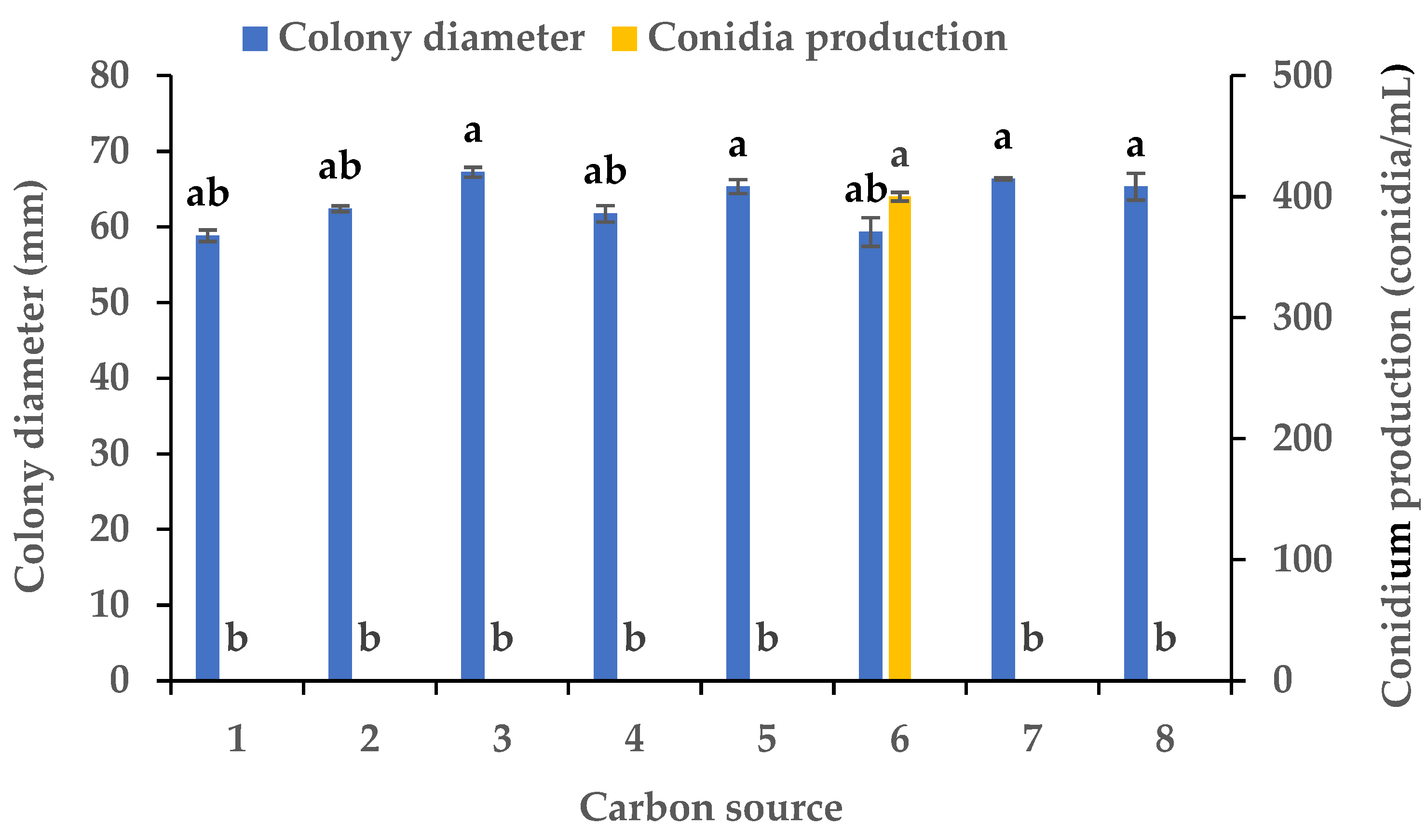
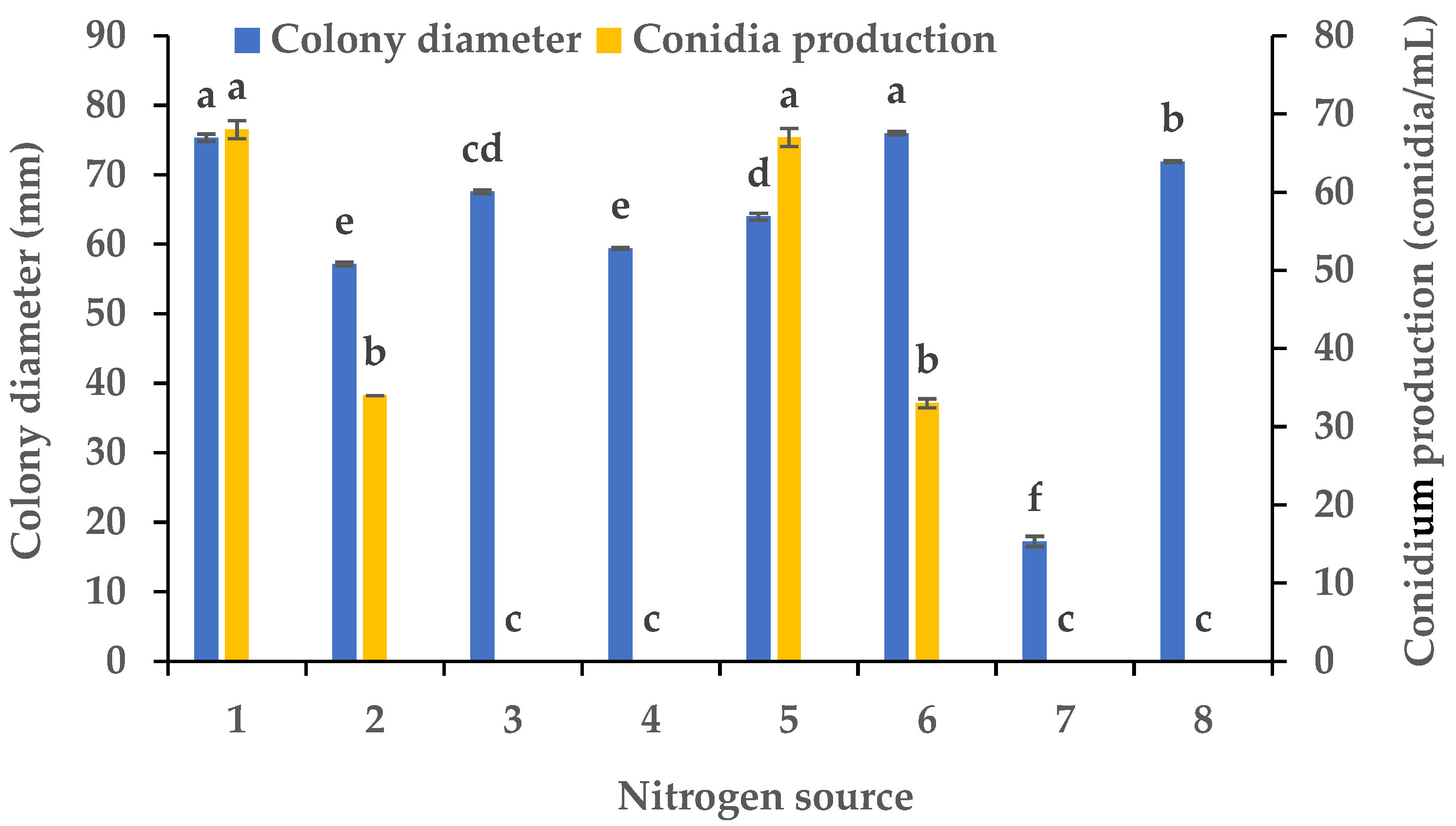
3.2. Effects of Carbon and Nitrogen Sources on the Mycelial Growth and Sporulation
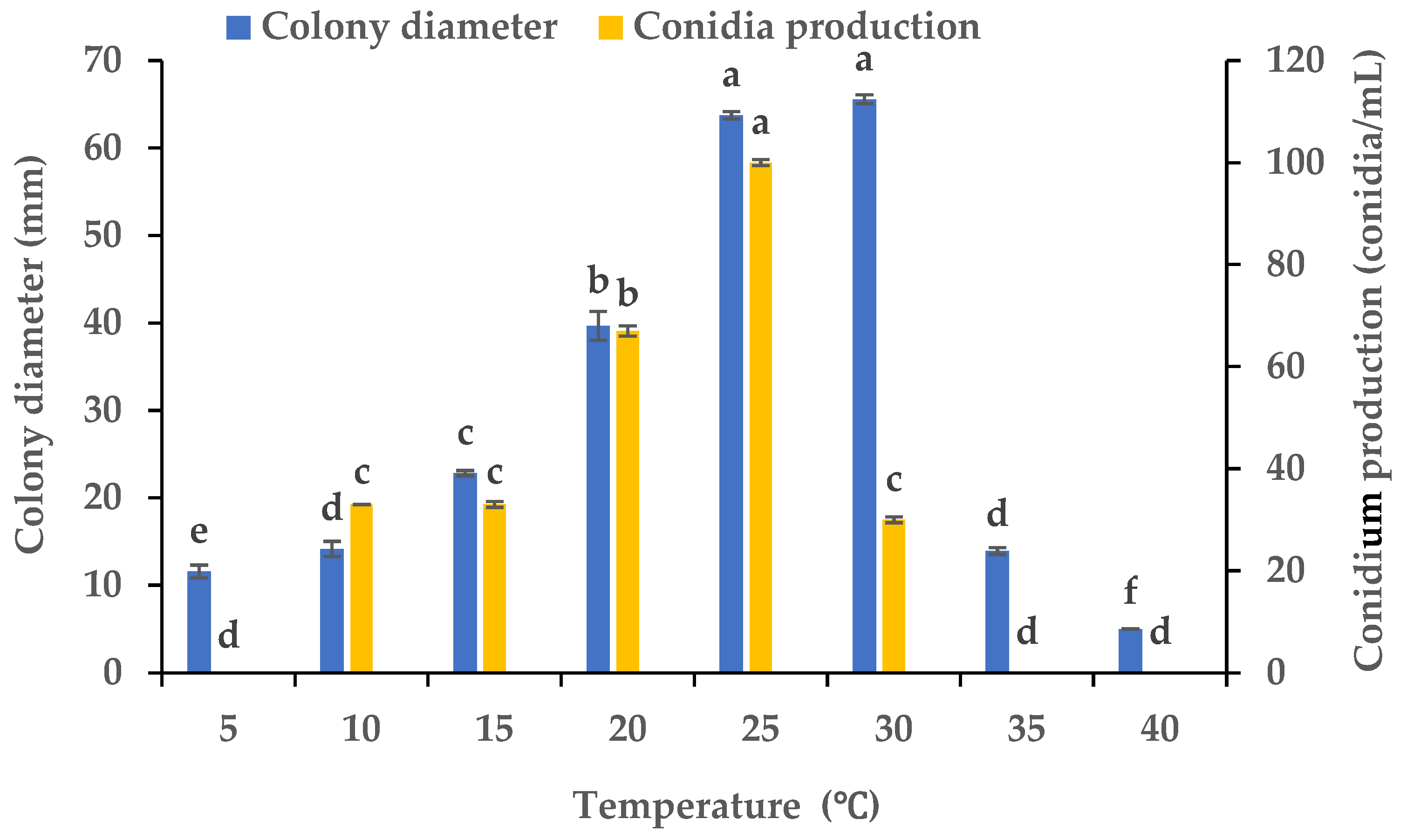
3.3. Effects of Various Temperatures on the Mycelial Growth and Sporulation
3.4. Effects of Various pH Values on the Mycelial Growth and Sporulation
3.5. Effects of Various Photoperiods on the Mycelial Growth and Sporulation
3.6. Determination of the Lethal Temperature for Mycelia
3.7. Antifungal Activities of Fungicides
3.8. Protective and Curative Activity of Fungicides
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crous, P.; Carlier, J.; Roussel, V.; Groenewald, J.Z. Pseudocercospora and allied genera associated with leaf spots of banana (Musa spp.). Fungal Syst. Evol. 2016, 7, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.Y.; Wu, Y.L.; Huang, B.Z. Research and utilization progress in banana germplasm resources in China. J. Fruit Sci. 2023, 40, 2446–2465. [Google Scholar]
- Jones, D.R. Handbook of Diseases of Banana, Abacá and Enset; CABI: Boston, USA, 2018; pp. 41–206. [Google Scholar]
- Zhao, H.; Qi, Y.X.; Zhang, X. First report of Bipolaris oryzae causing leaf spot on banana in China. J. Plant Pathol. 2023, 105, 1717–1718. [Google Scholar] [CrossRef]
- Gou, Y.Y.; Wu, K.M.; Chen, W.Q. Crop Disease and Insect Pests in China; China Agriculture Press: Beijing, China, 2015; pp. 250–252, 255–256. [Google Scholar]
- Deng, Y.Z.; Qu, Z.W.; He, Y.L.; Naqvi, N.I. Sorting nexin Snx41 is essential for conidiation and nediates glutathione-based antioxidant defense during invasive growth in Magnaporthe oryazae. Autophagy 2012, 8, 1058–1070. [Google Scholar] [CrossRef]
- Li, P.; Dai, X.; Wang, S.Y.; Luo, Q.; Tang, Q.Q.; Xu, Z.J.; Zhao, W.G.; Wu, F. Biological characteristics and fungicide screening of Colletotrichum fructicola causing mulberry anthracnose. Microorganisms 2024, 12, 2386. [Google Scholar] [CrossRef]
- Fan, F.; Li, L.; Li, G.Q.; Luo, C.X. Occurrence of fungicide resistance in Botrytis cinerea from greenhouse tomato in Hubei province, China. Plant Dis. 2016, 100, 2414–2421. [Google Scholar] [CrossRef]
- Ma, L.J.; Liu, X.; Guo, L.W.; Luo, Y.; Zhang, B.B.; Cui, X.X.; Yang, K.; Cai, J.; Liu, F.; Ma, N.; et al. Discovery of plant chemical defense mediated by a two-component system involving β-glucosidase in Panax species. Nat. Commun. 2024, 15, 602. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.Y.; Zhang, Z.Q.; Chen, L.L.; He, L.M.; Lu, H.B.; Ren, Y.P.; Mu, W.; Liu, F. Baseline sensitivity of Botrytis cinerea to the succinate dehydrogenase inhibitor isopyrazam and efficacy of this fungicide. Plant Dis. 2016, 100, 1314–1320. [Google Scholar] [CrossRef]
- Manamgoda, D.S.; Rossman, A.Y.; Castlebury, L.A.; Crous, P.W.; Chukeatirote, E.; Hyde, K.D. The genus Bipolaris. Stud. Mycol. 2014, 79, 221–228. [Google Scholar] [CrossRef]
- Huang, W.Q.; Li, Q.; Qin, Z.H.; Wang, M.H. Isolation and identification of the pathogen causing leaf spot of Zizania latifolia. Plant Protect. 2020, 46, 129–134. [Google Scholar]
- Li, S.; Cui, X.W.; Gao, M.; Wang, N.; Wang, Z.Y. First report of a leaf spot caused by Bipolaris oryzae on peanut (Arachis hypogaea) in China. Plant Dis. 2017, 101, 1821. [Google Scholar] [CrossRef]
- Fajolu, O.L. Characterization of Bipolaris species, Their Effects on Switchgrass Biomass Yield and Chemical Components. Ph.D. Thesis, University of Tennessee, Knoxville, TN, USA, 2012. [Google Scholar]
- Sanahuja, G.; Palmateer, A.J. First report of Bipolaris oryzae causing leaf spot on Aechmea tayoensis in Florida. Plant Dis. 2016, 100, 2329. [Google Scholar] [CrossRef]
- Huang, L.; Zhu, Y.N.; Yang, J.Y.; Li, D.W.; Li, Y.; Bian, L.M.; Ye, J.R. Shoot blight on Chinese fir (Cunninghamia lanceolata) is caused by Bipolaris oryzae. Plant Dis. 2018, 102, 500–506. [Google Scholar] [CrossRef]
- Rebolledo-Prudencio, O.G.; Chan-Cupul, W.; Moreno-Zúñiga, G.; Cruz-Jiménez, C.A.; Sánchez-Rangel, J.C. Bipolaris oryzae associated agent at the leaf spot disease in Cocos nucifera hybrid “Brazilian Green Dwarf”. Mex. J. Phytopathology 2025, 43, 55. [Google Scholar] [CrossRef]
- Sunder, S.; Singh, R.; Agarwal, R. Brown spot of rice: An overview. Indian Phytopath. 2014, 67, 201–205. [Google Scholar]
- Silva, B.B.; Gianfranco, R.; Antonio, J.A. Chitosan in the Preservation of Agricultural Commodities; Academic Press: Cambridge, MA, USA, 2016; pp. 115–125. [Google Scholar]
- Peng, C.M.; Wang, R.; Chen, H.L.; Wang, L.W.; Yuan, Y.; Guo, S.W. Cultural characteristics of pathogenic fungi of rice brown spot. Jiangsu J. Agr. Sci. 2014, 30, 503–507. [Google Scholar]
- Channakeshava, C.; Pankaja, N.S. Effect of media, temperature, light, pH and nutrient source on growth and development of Bipolaris oryzae causing brown leaf spot of paddy. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1713–1722. [Google Scholar]
- Jiang, D.Y.; Chen, X.J.; Chen, Y.F.; Chen, C.; Zhang, Z.P.; Wei, L.H. Pathogen identification of cane shoots leaf spot and determination of its sensitivity of five fungicides. China Veg. 2022, 35, 34–41. [Google Scholar]
- Dallagnol, L.J.; Rodrigues, F.A.; Da Matta, F.M. Brown spot of rice as affected by phyton irradiance and temperature. J. Phytopathol. 2011, 159, 630–634. [Google Scholar] [CrossRef]
- Kulkarni, S.; Ramakrishnan, K.; Hegde, R.K. Epidemiology and control of brown leaf spot of rice caused by Drechslera oryzae (Breda de Haan) Subram. and Jain in Karnataka. II. Survival of Drechslera oryzae in nature. Curr. Res. 1984, 10, 67. [Google Scholar]
- Feng, A.Q.; Zhu, X.Y.; Wang, C.Y.; Chen, K.L.; Feng, J.Q.; Yang, J.Y.; Lin, X.P.; Chen, X. Efficacy of thirteen fungicides for controlling rice brown spot disease. Plant Protect. 2022, 48, 352–360. [Google Scholar]
- Dong, Y.F.; Yang, M.F.; Peng, H.; Ye, Z.H.; Tang, J.T.; Li, Y.P.; Song, R.Q.; Zhang, Y.F. Screening and evaluation of effective fungicides for the control of leaf spot disease of Zizania latifolia. Acta Phytopathol. Sin. 2024. [Google Scholar] [CrossRef]
- Barúa, M.; Quintana, L.; Ortiz, A. Chemical control of rice brown spot (Bipolaris oryzae) in Paraguay. Trop. Plant Res. 2019, 6, 148–151. [Google Scholar] [CrossRef]
- Karan, R.; Renganathan, P.; Balabaskar, P. In-vitro evaluation of selected fungicides against Bipolaris oryzae causing leaf spot of rice. Res. J. Agril. Sci. 2021, 12, 1583–1584. [Google Scholar]
- Nargave, S.; Gehlot, J.; Burri, R.; Jakhar, M.; Damor, J.S.; Jain, S. Biological and chemical management strategy to control brown spot disease in rice caused by Bipolaris oryzae. Int. J. Plant Soil Sci. 2023, 35, 531–537. [Google Scholar] [CrossRef]
- Li, Q. Research on the Sporulation Characteristics and Mycoviruses in Brown Spot Disease of Zizania latifolia. Master Thesis, Hubei Minzu University, Enshi, China, 2020. [Google Scholar]
- Wu, S.G.; Xing, C.Y.; Zhu, J.J. Analysis of climate characteristics in Hainan Island. J. Trop. Biol. 2022, 13, 315–323. [Google Scholar]
- Zhang, X.; Zeng, F.Y.; Xie, Y.X.; Peng, J.; Qi, Y.X. Preliminary investigation on occurrence dynamics of freckle disease of banana. China Trop. Agric. 2022, 109, 65–69. [Google Scholar]
- Zhang, J.P.; Zhou, Y.J.; Han, C.; Lu, Y.L.; Yu, L.Q. Biological characteristics of Bipolaris oryzae from Leersia hexandra. Chinese Agr. Sci. Bull. 2014, 30, 124–127. [Google Scholar]
- Li, N.P.; He, P.B.; Liu, Y.L.; He, P.F.; Wu, Y.X.; Tang, P.; Wang, Z.Q.; Munire, S.; Kong, B.H.; HE, Y.Q. Identification and host range test of leaf spot pathogen of Axonopus compressus. Pratacultural Sci. 2024, 41, 2604–2611. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungicide | Manufacturer | Concentration (μg/mL) | Inhibitory Rates (%) |
|---|---|---|---|
| 250 g/L of azosystrobin SC | Syngenta Nantong Crop Protection Co., Ltd. (Nantong, China) | 0.5 | 42.35 |
| 5 | 56.34 | ||
| 10 | 59.4 | ||
| 20 | 62.61 | ||
| 25 | 64.3 | ||
| CK | — | ||
| 12.5% diniconazole WP | ADAMA Huifeng (Jiangsu) Co., Ltd. (Yancheng, China) | 0.5 | 35.29 |
| 5 | 72.03 | ||
| 10 | 84.58 | ||
| 20 | 89.94 | ||
| 25 | 91.44 | ||
| CK | — | ||
| 30% difenoconazole-azoxystrobin SC | Sipcam Agro USA (Shanghai, China) | 0.5 | 63.14 |
| 5 | 82.75 | ||
| 10 | 88.63 | ||
| 20 | 89.61 | ||
| 25 | 91.58 | ||
| CK | — | ||
| 30% difenoconazole-propiconazol EC | Sichuan Runer Technology Co., Ltd. (Jianyang, China) | 0.5 | 46.27 |
| 5 | 77.26 | ||
| 10 | 81.18 | ||
| 20 | 89.80 | ||
| 25 | 94.38 | ||
| CK | — | ||
| 24% fenbuconazole SC | Kedihua Agricultural Technology Co., Ltd. (Beijing, China) | 0.5 | 46.54 |
| 5 | 68.07 | ||
| 10 | 74.51 | ||
| 20 | 78.56 | ||
| 25 | 80.47 | ||
| CK | — | ||
| 43% mancozeb SC | Henan Xinnong Chemical Co., Ltd. (Zhengzhou, China) | 0.5 | 6.40 |
| 5 | 50.76 | ||
| 10 | 71.80 | ||
| 20 | 69.51 | ||
| 25 | 88.11 | ||
| CK | — | ||
| 70% propineb WP | Bayer Crop Science (China) Co., Ltd. (Beijing, China) | 0.5 | 8.24 |
| 5 | 49.15 | ||
| 10 | 85.23 | ||
| 20 | 89.54 | ||
| 25 | 92.83 | ||
| CK | — | ||
| 25% pyraclostrobin SC | Nanjing Huazhou Pharmaceutical Co., Ltd. (Nanjing, China) | 0.5 | 58.43 |
| 5 | 81.70 | ||
| 10 | 83.92 | ||
| 20 | 88.47 | ||
| 25 | 89.15 | ||
| CK | — | ||
| 40% chlorothalonil SC | SDS Biotech K. K. (Shanghai, China) | 0.5 | 6.54 |
| 5 | 11.63 | ||
| 10 | 23.92 | ||
| 20 | 28.89 | ||
| 25 | 32.03 | ||
| CK | — | ||
| 125 g/L of epoxiconazole SC | Shandong Jophne Biotechnology Co., Ltd. (Jinan, China) | 0.25 | 45.23 |
| 0.5 | 54.64 | ||
| 1.25 | 70.59 | ||
| 2.5 | 83.53 | ||
| 5 | 89.41 | ||
| CK | — | ||
| 500 g/L of iprodione SC | Guangdong Zhuoyue Biotechnology Co., Ltd. (Nanxiong, China) | 0.25 | 61.31 |
| 0.5 | 76.34 | ||
| 1.25 | 82.61 | ||
| 2.5 | 86.08 | ||
| 5 | 100 | ||
| CK | — | ||
| 450 g/L of prochloraz EW | Zhejiang Tianfeng Bioscience Co., Ltd. (Jinhua, China) | 0.25 | 38.43 |
| 0.5 | 73.86 | ||
| 1.25 | 75.1 | ||
| 2.5 | 78.04 | ||
| 5 | 83.06 | ||
| CK | — |
| Fungicide | Regression Equation | Correlation Coefficient | EC50 (μg/mL) | 95% Confidence Intervals (μg/mL) |
|---|---|---|---|---|
| 250 g/L of azoxystrobin SC | y = 0.3247 x + 4.9126 | 0.9984 | 1.86 | 0.00–850.65 |
| 12.5% diniconazole WP | y = 1.0399 x + 4.9222 | 0.9981 | 1.19 | 0.09–14.88 |
| 30% difenoconazole-azoxystrobin SC | y = 0.6054 x + 5.5285 | 0.9937 | 0.13 | 0.00–387.124 |
| 30% difenoconazole-propiconazol EC | y = 0.9122 x + 5.1318 | 0.9802 | 0.72 | 0.02–21.01 |
| 24% fenbuconazole SC | y = 0.5551 x + 5.0835 | 0.9994 | 0.71 | 0.00–125.18 |
| 43% mancozeb SC | y = 1.4597 x + 3.9541 | 0.9785 | 5.21 | 1.62–16.78 |
| 70% propineb WP | y = 1.7118 x + 4.0682 | 0.9859 | 3.50 | 1.05–11.73 |
| 25% pyraclostrobin SC | y = 0.6009 x + 5.4159 | 0.9954 | 0.20 | 0.00–251.10 |
| 40% chlorothalonil SC | y = 0.6396 x + 3.5974 | 0.9549 | 155.98 | 0.11–221,352.16 |
| 125 g/L of epoxiconazole SC | y = 1.0358 x + 5.7245 | 0.9475 | 0.20 | 0.02–2.22 |
| 500 g/L of iprodione SC | y = 0.7636 x + 5.8348 | 0.9658 | 0.08 | 0.00–2.31 |
| 450 g/L of prochloraz EW | y = 0.6961 x + 5.1897 | 0.9753 | 0.53 | 0.02–20.90 |
| Treatment | Protective Activity | Curative Activity | ||
|---|---|---|---|---|
| Mean Lesion Diameter (mm) | Control Efficacy (%) | Mean Lesion Diameter (mm) | Control Efficacy (%) | |
| 125 g/L of epoxiconazole SC (250 μg/mL) | 0 ± 0 b | 100 ± 0 a | 2.25 ± 0.56 b | 64 ± 5.09 a |
| 500 g/L of iprodione SC (1667 μg/mL) | 0 ± 0 b | 100 ± 0 a | 2.5 ± 0.77 c | 60 ± 4.86 a |
| 30% difenoconazole-azoxystrobin SC (250 μg/mL) | 0 ± 0 b | 100 ± 0 a | 4 ± 1.14 c | 36 ± 2.57 b |
| CK | 9.25 ± 3.3 a | – | 6.25 ± 1.54 a | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, Y.; Zhao, H.; Zhang, Z.; Ouyang, Y.; Zhang, X. Biological Characteristics and Fungicide Screening of Bipolaris oryzae Causing Leaf Spot on Banana in China. Microorganisms 2025, 13, 1285. https://doi.org/10.3390/microorganisms13061285
Qi Y, Zhao H, Zhang Z, Ouyang Y, Zhang X. Biological Characteristics and Fungicide Screening of Bipolaris oryzae Causing Leaf Spot on Banana in China. Microorganisms. 2025; 13(6):1285. https://doi.org/10.3390/microorganisms13061285
Chicago/Turabian StyleQi, Yanxiang, Hong Zhao, Zhaojing Zhang, Yanfei Ouyang, and Xin Zhang. 2025. "Biological Characteristics and Fungicide Screening of Bipolaris oryzae Causing Leaf Spot on Banana in China" Microorganisms 13, no. 6: 1285. https://doi.org/10.3390/microorganisms13061285
APA StyleQi, Y., Zhao, H., Zhang, Z., Ouyang, Y., & Zhang, X. (2025). Biological Characteristics and Fungicide Screening of Bipolaris oryzae Causing Leaf Spot on Banana in China. Microorganisms, 13(6), 1285. https://doi.org/10.3390/microorganisms13061285