

Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 Supplementation: An Exploratory, Randomized, Placebo-Controlled Trial of Endocannabinoid and Inflammatory Responses in Female Dancers

 ,
,  ,
,  , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
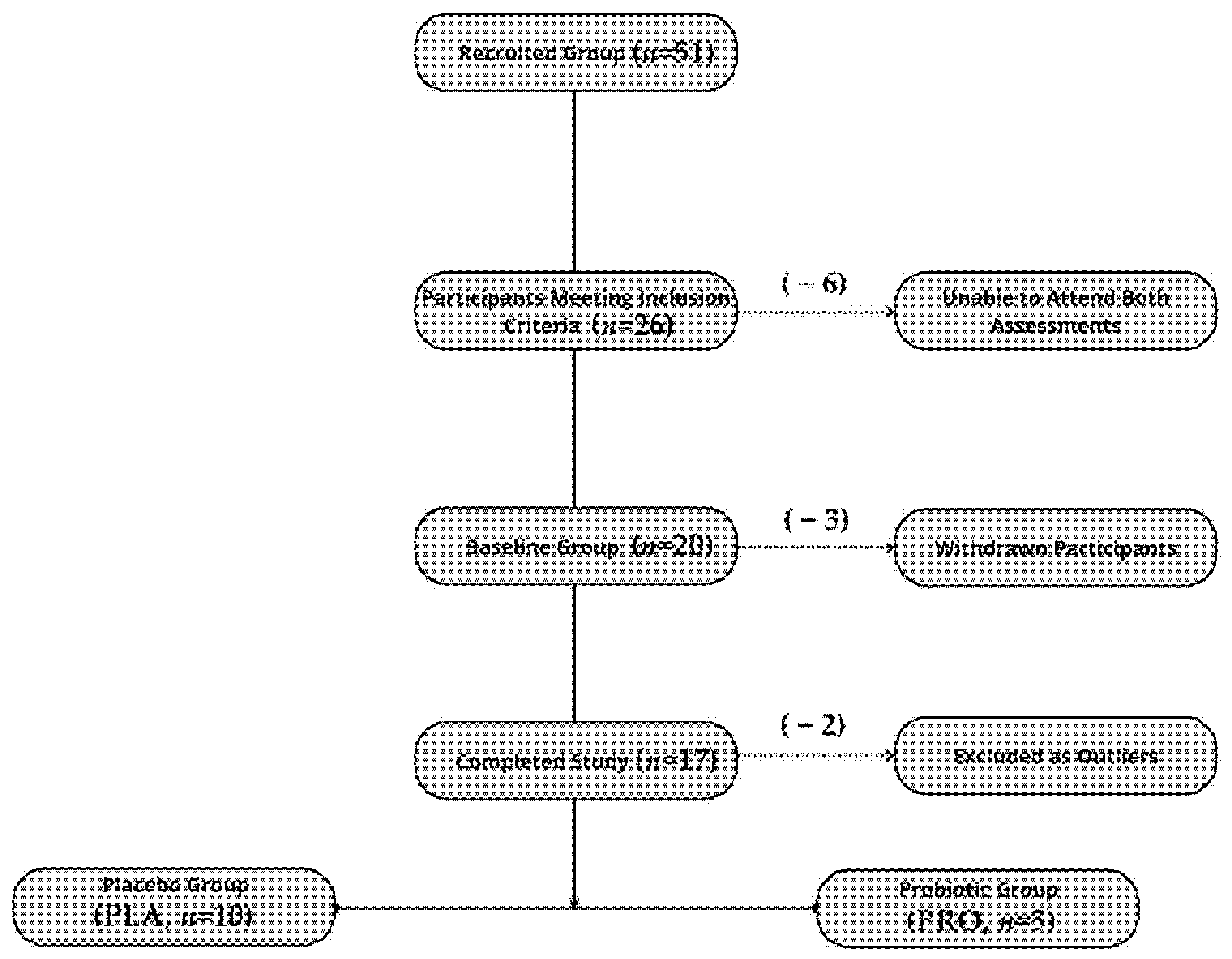
2.1. Participants
2.2. Intervention
2.3. Study Protocol
2.4. Methods
2.4.1. Outcome Measures
2.4.2. Statistical Analysis
3. Results
3.1. No Baseline Differences in Anthropometric or Dietary Variables Between the Groups
3.2. No Statistically Significant Changes in AEA, LPS, or Cytokine Levels Following Supplementation
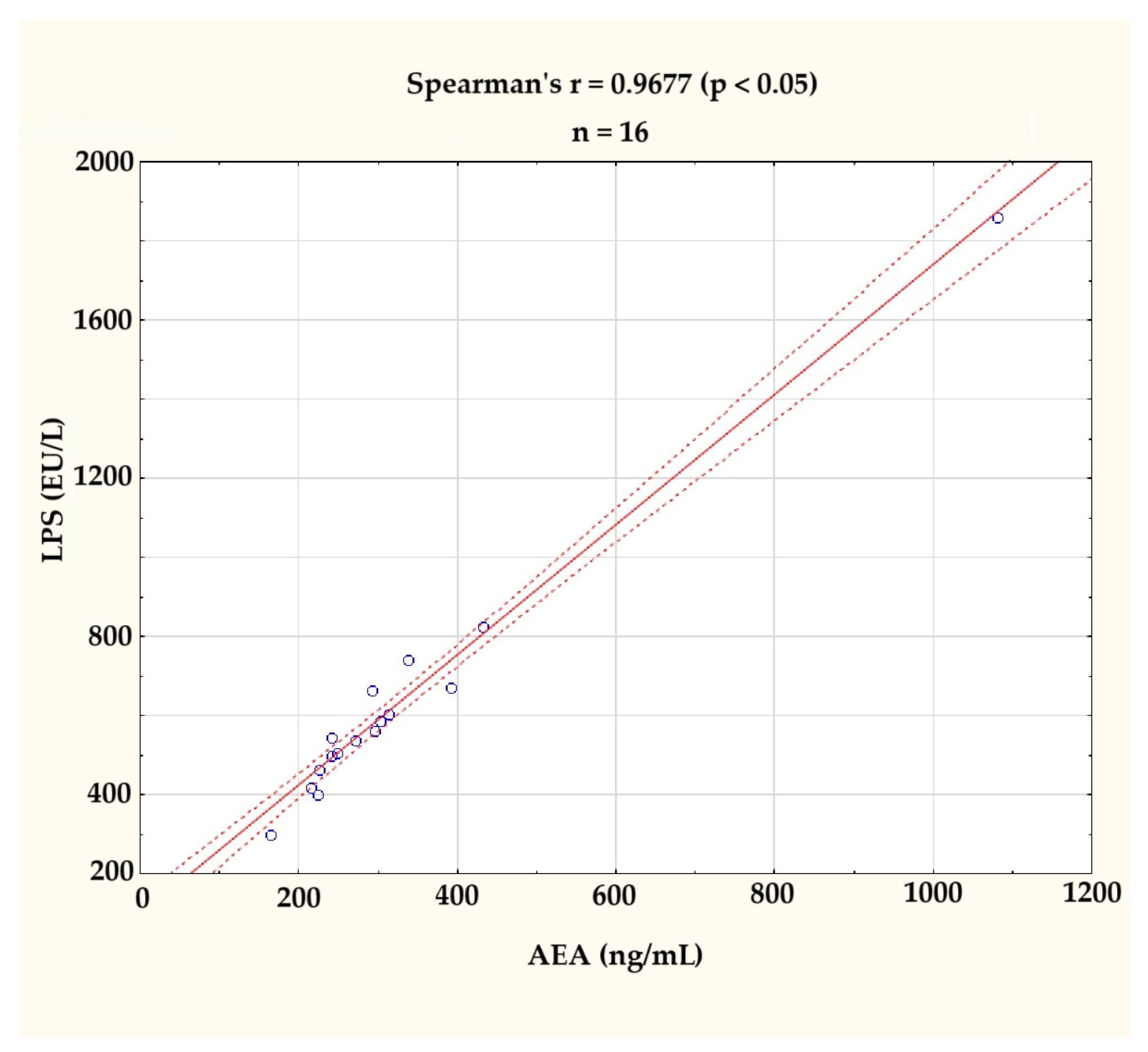
3.2.1. Strong Positive Correlation Between AEA and LPS at the Baseline
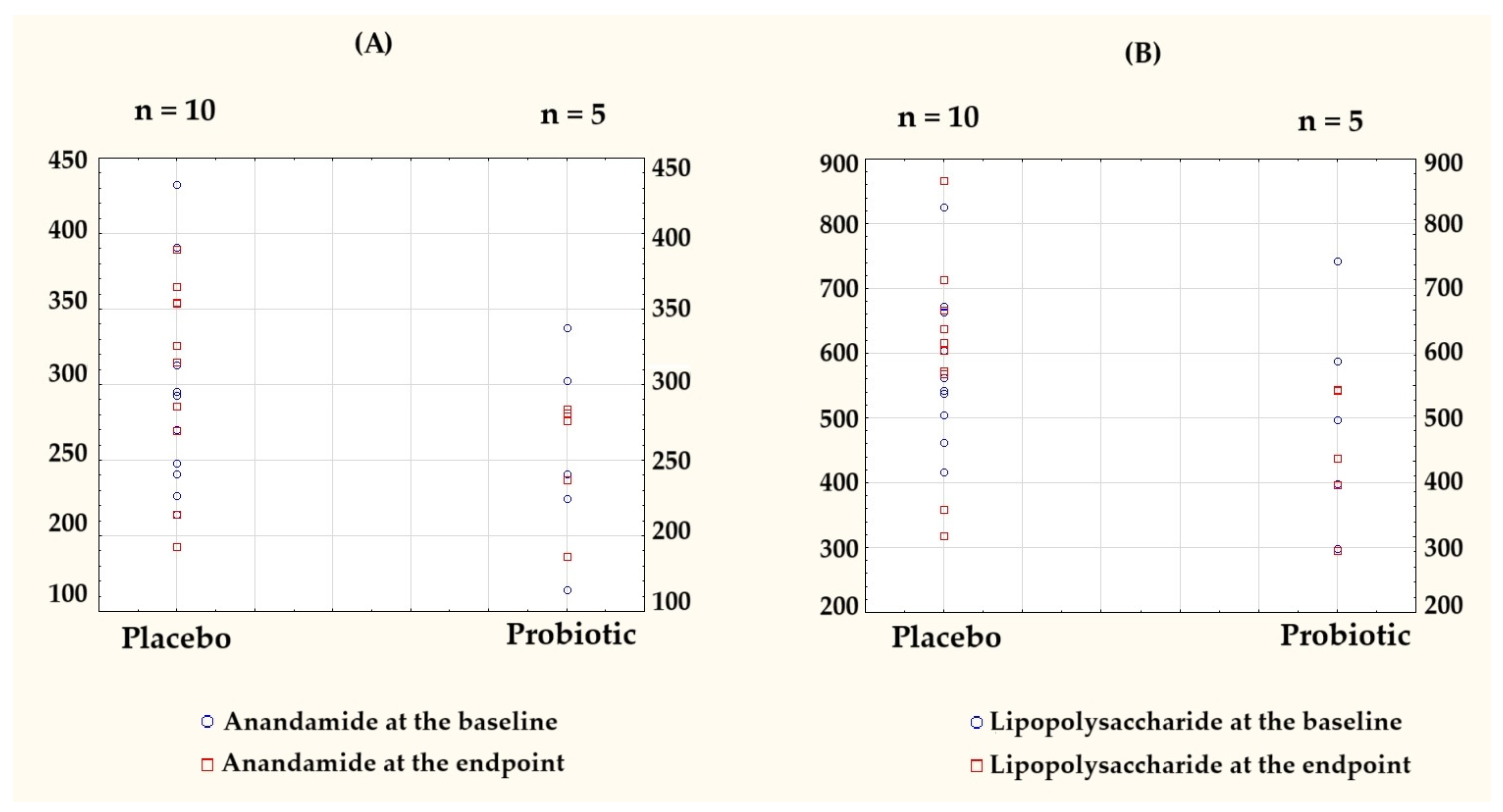
3.2.2. Directional Trends Observed, but Between- and Within-Group Changes Were Not Significant
3.3. No Effect of Probiotic Supplementation on Stress-Coping Strategies
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AEA | Anandamide |
| AFUs | Active fluorescent units |
| ANOVA | Analysis of variance |
| BDNF | Brain-derived neurotrophic factor |
| BIA | Bioelectrical impedance analysis |
| BMI | Body mass index |
| CB1 | Cannabinoid receptor type 1 |
| CB2 | Cannabinoid receptor type 2 |
| CFUs | Colony forming units |
| CONSORT | Consolidated Standards of Reporting Trials |
| ECS | Endocannabinoid system |
| ELISA | Enzyme-linked immunosorbent assay |
| EU/L | Endotoxin units per liter |
| GI | Gastrointestinal |
| HPA axis | Hypothalamic–pituitary–adrenal axis |
| IBS | Irritable bowel syndrome |
| IL-1β | Interleukin-1 beta |
| IL-10 | Interleukin-10 |
| LPS | Lipopolysaccharide |
| MDAT | Mediterranean Diet Assessment Tool |
| Mini-COPE | Mini-Coping Orientation to Problems Experienced |
| MDD | Major depressive disorder |
| NF-κB | Nuclear Factor Kappa-light-chain-enhancer of Activated B Cells |
| SCFA | Short-chain fatty acids |
| SD | Standard deviation |
| SHIME® | Simulator of the Human Intestinal Microbial Ecosystem |
| SMD | Standardized mean difference |
| TNF-α | Tumor necrosis factor-alpha |
| URTI | Upper respiratory tract infection |
| WBC | White blood cell |
References
- Pertwee, R.G. Cannabinoid pharmacology: The first 66 years. Br. J. Pharmacol. 2006, 147 (Suppl 1), S163–S171. [Google Scholar] [CrossRef] [PubMed]
- Ohno-Shosaku, T.; Tanimura, A.; Hashimotodani, Y.; Kano, M. Endocannabinoids and retrograde modulation of synaptic transmission. Neuroscientist 2012, 18, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Ligresti, A.; Cascio, M.G.; Di Marzo, V. Endocannabinoid metabolic pathways and enzymes. Curr. Drug Targets CNS Neurol. Disord. 2005, 4, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Katona, I.; Freund, T.F. Multiple functions of endocannabinoid signaling in the brain. Annu. Rev. Neurosci. 2012, 35, 529–558. [Google Scholar] [CrossRef]
- Tantimonaco, M.; Ceci, R.; Sabatini, S.; Catani, M.V.; Rossi, A.; Gasperi, V.; Maccarrone, M. Physical activity and the endocannabinoid system: An overview. Cell Mol. Life Sci. 2014, 71, 2681–2698. [Google Scholar] [CrossRef]
- Matei, D.; Trofin, D.; Iordan, D.A.; Onu, I.; Condurache, I.; Ionite, C.; Buculei, I. The Endocannabinoid System and Physical Exercise. Int. J. Mol. Sci. 2023, 24, 1989. [Google Scholar] [CrossRef]
- Monda, V.; Villano, I.; Messina, A.; Valenzano, A.; Esposito, T.; Moscatelli, F.; Viggiano, A.; Cibelli, G.; Chieffi, S.; Monda, M.; et al. Exercise Modifies the Gut Microbiota with Positive Health Effects. Oxid. Med. Cell Longev. 2017, 2017, 3831972. [Google Scholar] [CrossRef]
- Dziewiecka, H.; Buttar, H.S.; Kasperska, A.; Ostapiuk–Karolczuk, J.; Domagalska, M.; Cichoń, J.; Skarpańska-Stejnborn, A. Physical activity induced alterations of gut microbiota in humans: A systematic review. BMC Sports Sci. Med. Rehabil. 2022, 14, 122. [Google Scholar] [CrossRef]
- Clark, A.; Mach, N. Exercise-induced stress behavior, gut-microbiota-brain axis and diet: A systematic review for athletes. J. Int. Soc. Sports Nutr. 2016, 13, 43. [Google Scholar] [CrossRef]
- McBride, C.; Bronner, S. Injury characteristics in professional modern dancers: A 15-year analysis of work-related injury rates and patterns. J. Sports Sci. 2022, 40, 821–837. [Google Scholar] [CrossRef]
- Nunes, A.C.; Mendes, L.A.; Mota, L.A.; Lima, P.O.; Almeida, G.P. Training Load, Pain Intensity, and Functioning Can Explain Injuries in Dancers: A Classification and Regression Tree (CART) Analysis. Med. Probl. Perform. Art. 2022, 37, 73–77. [Google Scholar] [CrossRef] [PubMed]
- van Winden, D.; van Rijn, R.M.; Savelsbergh, G.J.P.; Oudejans, R.R.D.; Stubbe, J.H. The Association Between Stress. and Injury: A Prospective Cohort Study Among 186 First-Year Contemporary Dance Students. Front. Psychol. 2021, 12, 770494. [Google Scholar] [CrossRef] [PubMed]
- Berndt, C.; Strahler, J.; Kirschbaum, C.; Rohleder, N. Lower stress system activity and higher peripheral inflammation in competitive ballroom dancers. Biol. Psychol. 2012, 91, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Hendry, D.; Campbell, A.; Smith, A.; Hopper, L.; Straker, L.; O’Sullivan, P. Movement quantity and quality: How do they relate to pain and disability in dancers? PLoS ONE 2022, 17, e0268444. [Google Scholar] [CrossRef]
- Cani, P.D.; Plovier, H.; Van Hul, M.; Geurts, L.; Delzenne, N.M.; Druart, C.; Everard, A. Endocannabinoids—At the crossroads between the gut microbiota and host metabolism. Nat. Rev. Endocrinol. 2016, 12, 133–143. [Google Scholar] [CrossRef]
- Rousseaux, C.; Thuru, X.; Gelot, A.; Barnich, N.; Neut, C.; Dubuquoy, L.; Dubuquoy, C.; Merour, E.; Geboes, K.; Chamaillard, M.; et al. Lactobacillus acidophilus modulates intestinal pain and induces opioid and cannabinoid receptors. Nat. Med. 2007, 13, 35–37. [Google Scholar] [CrossRef]
- Brierley, S.M.; Greenwood-Van Meerveld, B.; Sarnelli, G.; Sharkey, K.A.; Storr, M.; Tack, J. Targeting the endocannabinoid system for the treatment of abdominal pain in irritable bowel syndrome. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 5–25. [Google Scholar] [CrossRef]
- Malek, N.; Popiolek-Barczyk, K.; Mika, J.; Przewlocka, B.; Starowicz, K. Anandamide, Acting via CB2 Receptors, Alleviates LPS-Induced Neuroinflammation in Rat Primary Microglial Cultures. Neural Plast. 2015, 2015, 130639. [Google Scholar] [CrossRef]
- Berg, B.B.; Linhares, A.F.S.; Martins, D.M.; Rachid, M.A.; Cau, S.B.d.A.; de Souza, G.G.; de Carvalho, J.C.S.; Sorgi, C.A.; Romero, T.R.L.; Pinho, V.; et al. Anandamide reduces the migration of lymphocytes to the intestine by CB2 activation and reduces TNF-α in the target organs, protecting mice from graft-versus-host disease. Eur. J. Pharmacol. 2023, 956, 175932. [Google Scholar] [CrossRef]
- Vercelli, C.A.; Aisemberg, J.; Billi, S.; Wolfson, M.L.; Franchi, A.M. Endocannabinoid system and nitric oxide are involved in the deleterious effects of lipopolysaccharide on murine decidua. Placenta 2009, 30, 579–584. [Google Scholar] [CrossRef]
- Liu, J.; Batkai, S.; Pacher, P.; Harvey-White, J.; Wagner, J.A.; Cravatt, B.F.; Gao, B.; Kunos, G. Lipopolysaccharide induces anandamide synthesis in macrophages via CD14/MAPK/phosphoinositide 3-kinase/NF-kappaB independently of platelet-activating factor. J. Biol. Chem. 2003, 278, 45034–45039. [Google Scholar] [CrossRef] [PubMed]
- Molina-Holgado, F.; Lledó, A.; Guaza, C. Anandamide suppresses nitric oxide and TNF-alpha responses to Theiler’s virus or endotoxin in astrocytes. Neuroreport 1997, 8, 1929–1933. [Google Scholar] [CrossRef] [PubMed]
- Jansma, J.; Brinkman, F.; van Hemert, S.; El Aidy, S. Targeting the endocannabinoid system with microbial interventions to improve gut integrity. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 106, 110169. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M.; De Petrocellis, L.; Bari, M.; Fezza, F.; Salvati, S.; Di Marzo, V.; Finazzi-Agrò, A. Lipopolysaccharide downregulates fatty acid amide hydrolase expression and increases anandamide levels in human peripheral lymphocytes. Arch. Biochem. Biophys. 2001, 393, 321–328. [Google Scholar] [CrossRef]
- Khan, R.N.; Maner-Smith, K.; Owens, J.A.; Barbian, M.E.; M Jones, R.; Naudin, C.R. At the heart of microbial conversations: Endocannabinoids and the microbiome in cardiometabolic risk. Gut Microbes 2021, 13, 1911572. [Google Scholar] [CrossRef]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: A Door to the Body. Front. Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef]
- Rodes, L.; Khan, A.; Paul, A.; Coussa-Charley, M.; Marinescu, D.; Tomaro-Duchesneau, C.; Shao, W.; Kahouli, I.; Prakash, S. Effect of probiotics Lactobacillus and Bifidobacterium on gut-derived lipopolysaccharides and inflammatory cytokines: An in vitro study using a human colonic microbiota model. J. Microbiol. Biotechnol. 2013, 23, 518–526. [Google Scholar] [CrossRef]
- Wang, H.; Lee, I.S.; Braun, C.; Enck, P. Effect of Probiotics on Central Nervous System Functions in Animals and Humans: A Systematic Review. J. Neurogastroenterol. Motil. 2016, 22, 589–605. [Google Scholar] [CrossRef]
- Mohammadi, G.; Dargahi, L.; Peymani, A.; Mirzanejad, Y.; Alizadeh, S.A.; Naserpour, T.; Nassiri-Asl, M. The Effects of Probiotic Formulation Pretreatment (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) on a Lipopolysaccharide Rat Model. J. Am. Coll. Nutr. 2019, 38, 209–217. [Google Scholar] [CrossRef]
- Mohammadi, G.; Dargahi, L.; Naserpour, T.; Mirzanejad, Y.; Alizadeh, S.A.; Peymani, A.; Nassiri-Asl, M. Probiotic mixture of Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 attenuates hippocampal apoptosis induced by lipopolysaccharide in rats. Int. Microbiol. 2019, 22, 317–323. [Google Scholar] [CrossRef]
- Partrick, K.A.; Rosenhauer, A.M.; Auger, J.; Arnold, A.R.; Ronczkowski, N.M.; Jackson, L.M.; Lord, M.N.; Abdulla, S.M.; Chassaing, B.; Huhman, K.L. Ingestion of probiotic (Lactobacillus helveticus and Bifidobacterium longum) alters intestinal microbial structure and behavioral expression following social defeat stress. Sci. Rep. 2021, 11, 3763. [Google Scholar] [CrossRef] [PubMed]
- Ait-Belgnaoui, A.; Payard, I.; Rolland, C.; Harkat, C.; Braniste, V.; Théodorou, V.; Tompkins, T.A. Bifidobacterium longum and Lactobacillus helveticus Synergistically Suppress Stress-related Visceral Hypersensitivity Through Hypothalamic-Pituitary-Adrenal Axis Modulation. J. Neurogastroenterol. Motil. 2018, 24, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Myles, E.M.; O’Leary, M.E.; Smith, R.; MacPherson, C.W.; Oprea, A.; Melanson, E.H.; Tompkins, T.A.; Perrot, T.S. Supplementation with Combined Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 Across Development Reveals Sex Differences in Physiological and Behavioural Effects of Western Diet in Long-Evans Rats. Microorganisms 2020, 8, 1527. [Google Scholar] [CrossRef] [PubMed]
- Esposito, P.; Kearns, M.M.; Smith, K.B.; MacPherson, C.W.; Oprea, A.; Melanson, E.H.; Tompkins, T.A.; Perrot, T.S. The effects of antimicrobials and lipopolysaccharide on acute immune responsivity in pubertal male and female CD1 mice. Compr. Psychoneuroendocrinology 2022, 11, 100147. [Google Scholar] [CrossRef]
- Misera, A.; Liśkiewicz, P.; Łoniewski, I.; Skonieczna-Żydecka, K.; Samochowiec, J. Effect of Psychobiotics on Psychometric Tests and Inflammatory Markers in Major Depressive Disorder: Meta-Analysis of Randomized Controlled Trials with Meta-Regression. Pharmaceuticals 2021, 14, 952. [Google Scholar] [CrossRef]
- Gil-Hernández, E.; Ruiz-González, C.; Rodriguez-Arrastia, M.; Ropero-Padilla, C.; Rueda-Ruzafa, L.; Sánchez-Labraca, N.; Roman, P. Effect of gut microbiota modulation on sleep: A systematic review and meta-analysis of clinical trials. Nutr. Rev. 2023, 81, 1556–1570. [Google Scholar] [CrossRef]
- Rode, J.; Edebol Carlman, H.M.T.; König, J.; Repsilber, D.; Hutchinson, A.N.; Thunberg, P.; Andersson, P.; Persson, J.; Kiselev, A.; Stern, L.L.; et al. Probiotic Mixture Containing Lactobacillus helveticus, Bifidobacterium longum and Lactiplantibacillus plantarum Affects Brain Responses Toward an Emotional Task in Healthy Subjects: A Randomized Clinical Trial. Front. Nutr. 2022, 9, 827182. [Google Scholar] [CrossRef]
- Varghese, S.; Rao, S.; Khattak, A.; Zamir, F.; Chaari, A. Physical Exercise and the Gut Microbiome: A Bidirectional Relationship Influencing Health and Performance. Nutrients 2024, 16, 3663. [Google Scholar] [CrossRef]
- Mohr, A.E.; Pugh, J.; O’Sullivan, O.; Black, K.; Townsend, J.R.; Pyne, D.B.; Wardenaar, F.C.; West, N.P.; Whisner, C.M.; McFarland, L.V. Best Practices for Probiotic Research in Athletic and Physically Active Populations: Guidance for Future Randomized Controlled Trials. Front. Nutr. 2022, 9, 809983. [Google Scholar] [CrossRef]
- Aykut, M.N.; Erdoğan, E.N.; Çelik, M.N.; Gürbüz, M. An Updated View of the Effect of Probiotic Supplement on Sports Performance: A Detailed Review. Curr. Nutr. Rep. 2024, 13, 251–263. [Google Scholar] [CrossRef]
- Liu, M.; Tandorost, A.; Moludi, J.; Dey, P. Prebiotics Plus Probiotics May Favorably Impact on Gut Permeability, Endocannabinoid Receptors, and Inflammatory Biomarkers in Patients with Coronary Artery Diseases: A Clinical Trial. Food Sci. Nutr. 2023, 12, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, Z.; Tang, P.; Wu, Y.; Zhang, A.; Li, D.; Wang, C.-Z.; Wan, J.-Y.; Yao, H.; Yuan, C.-S. Probiotics fortify intestinal barrier function: A systematic review and meta-analysis of randomized trials. Front. Immunol. 2023, 14, 1143548. [Google Scholar] [CrossRef] [PubMed]
- Milajerdi, A.; Mousavi, S.M.; Sadeghi, A.; Salari-Moghaddam, A.; Parohan, M.; Larijani, B.; Esmaillzadeh, A. The effect of probiotics on inflammatory biomarkers: A meta-analysis of randomized clinical trials. Eur. J. Nutr. 2020, 59, 633–649. [Google Scholar] [CrossRef] [PubMed]
- Łagowska, K.; Bajerska, J. Probiotic Supplementation and Respiratory Infection and Immune Function in Athletes: Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Athl. Train. 2021, 56, 1213–1223. [Google Scholar] [CrossRef]
- Johnson, D.; Letchumanan, V.; Thum, C.C.; Thurairajasingam, S.; Lee, L.H. A Microbial-Based Approach to Mental Health: The Potential of Probiotics in the Treatment of Depression. Nutrients 2023, 15, 1382. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Lutz, B.; Ruiz de Azua, I. The Microbiome and Gut Endocannabinoid System in the Regulation of Stress Responses and Metabolism. Front. Cell Neurosci. 2022, 16, 867267. [Google Scholar] [CrossRef]
- Messaoudi, M.; Lalonde, R.; Violle, N.; Javelot, H.; Desor, D.; Nejdi, A.; Bisson, J.F.; Rougeot, C.; Pichelin, M.; Cazaubiel, M.; et al. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br. J. Nutr. 2011, 105, 755–764. [Google Scholar] [CrossRef]
- Romijn, A.R.; Rucklidge, J.J.; Kuijer, R.G.; Frampton, C. A double-blind, randomized, placebo-controlled trial of Lactobacillus helveticus and Bifidobacterium longum for the symptoms of depression. Aust. N. Z. J. Psychiatry 2017, 51, 810–821. [Google Scholar] [CrossRef]
- Kazemi, A.; Noorbala, A.A.; Azam, K.; Eskandari, M.H.; Djafarian, K. Effect of probiotic and prebiotic vs placebo on psychological outcomes in patients with major depressive disorder: A randomized clinical trial. Clin. Nutr. 2019, 38, 522–528. [Google Scholar] [CrossRef]
- De Oliveira, F.L.; Salgaço, M.K.; de Oliveira, M.T.; Mesa, V.; Sartoratto, A.; Peregrino, A.M.; Ramos, W.S.; Sivieri, K. Exploring the Potential of Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 as Promising Psychobiotics Using SHIME. Nutrients 2023, 15, 1521. [Google Scholar] [CrossRef]
- Heimer, M.; Teschler, M.; Schmitz, B.; Mooren, F.C. Health Benefits of Probiotics in Sport and Exercise—Non-existent or a Matter of Heterogeneity? A Systematic Review. Front. Nutr. 2022, 9, 1051918, Erratum in Front. Nutr. 2022, 9, 804046. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probiotic (n = 5) | Placebo (n = 11) | Total (n = 16) | Placebo vs. Probiotic (p Value) | |
|---|---|---|---|---|
| Mean ± SD (Min–Max) | Mean ± SD (Min–Max) | Mean ± SD (Min–Max) | ||
| Dancing activity [hours per week] | 16.00 ± 9.77 (9.50–29.00) | 17.11 ± 6.98 (8.00–33.00) | 16.77 ± 7.63 (8.00–33.00) | 0.69 b |
| Age [years] | 20.00 ± 1.30 (19–22) | 20.55 ± 1.04 (19–22) | 20.44 ± 1.09 (19–22) | 0.55 b |
| Body mass [kg] | 60.10 ± 7.31 (48.60–68.30) | 58.07 ± 6.95 (49.40–68.70) | 58.08 ± 6.81 (48.60–68.70) | 0.99 a |
| BMI (body mass index) [kg/m2] | 20.80 ± 2.29 (18.10–25.10) | 21.05 ± 2.18 (17.70–23.40) | 21.02 ± 2.13 (17.70–25.10) | 0.93 a |
| Fat [% body mass] | 27 ± 3 (25–31) | 27 ± 4 (21–31) | 27 ± 3 (21–31) | 0.84 a |
| WBC [×109/L] | 5.94 ± 1.17 (4.5–6.6) | 5.54 ± 0.84 (4.0–7.5) | 5.66 ± 1.07 (4.0–7.5) | 0.50 a |
| Lymphocytes [×109/L] | 2.60 ± 0.46 (1.6–3.3) | 2.34 ± 0.79 (1.8–3.4) | 2.42 ± 0.57 (1.6–3.4) | 0.41 a |
| Energy [kcal] | 2325.54 ± 425.00 (1835.0–2842.6) | 1999.23 ± 279.81 (1588.93–2578.45) | 2101.20 ± 353.22 (1588.93–2842.6) | 0.26 a |
| Protein [g] | 100.47 ± 21.09 (79.42–130.92) | 85.29 ± 30.13 (49.52–154.26) | 90.03 ± 27.87 (49.52–154.26) | 0.51 a |
| Fat [g] | 90.47 ± 18.72 (67.02–115.73) | 74.56 ± 13.93 (57.69–95.74) | 79.54 ± 16.76 (57.69–115.73) | 0.41 a |
| Cholesterol [mg] | 330.21 ± 140.77 (178.32–490.73) | 216.73 ± 104.36 (9–372.03) | 252.19 ± 124.49 (9–490.73) | 0.40 a |
| Carbohydrates [g] | 298.79 ± 57.98 (240.54–376.75) | 271.12 ± 53.14 (194.89–359.42) | 279.77 ± 54.36 (194.89–376.75) | 0.75 a |
| Fiber [g] | 21.36 ± 12.67 (15.03–27.63) | 28.96 ± 15.13 (16.69–47.68) | 26.59 ± 14.44 (15.03–47.68) | 0.91 b |
| Mediterranean diet adherence [0–14] | 5.46 ± 1.86 (3.00–10.00) | 6.20 ± 2.49 (3.00–10.00) | 5.69 ± 2.02 (3.00–10.00) | 0.42 a |
| Mean | Probiotic Group (n = 5) | Placebo Group (n = 10) | (Independent t-Test p) | ||
|---|---|---|---|---|---|
| Baseline (Mean ± SD) (Min–Max) | (Shapiro–Wilk p) | Baseline (Mean ± SD) (Min–Max) | (Shapiro–Wilk p) | ||
| LPS [EU/L*] | 505.05 ± 170.77 299.35–742.17 | 0.9796 | 579.36 ± 118.50 416.63–826.21 | 0.6987 | 0.3391 |
| AEA [ng/mL] | 253.99 ± 68,01 164.05–337.67 | 0.8996 | 292.59 ± 70.98 214.40–432.66 | 0.1966 | 0.3329 |
| TNF-α [pg/mL] | 87.52 ± 24.40 56.75–117.74 | 0.9029 | 105.89 ± 22.23 72.16–139.53 | 0.8120 | 0.1671 |
| IL-1β [pg/mL] | 1300.42 ± 298.48 982.44−1689.51 | 0.4774 | 1475.79 ± 333.89 940.79–2090.87 | 0.9272 | 0.3402 |
| IL-10 [pg/mL] | 490.03 ± 110.89 393.35–679.28 | 0.1302 | 515.46 ± 96.86 397.89–709.14 | 0.6118 | 0.6546 |
| Mean | Probiotic Group (n = 5) | Placebo Group (n = 10) | (Independent t-Test p) | ||
|---|---|---|---|---|---|
| Endpoint (Mean ± SD) (Min–Max) | (Shapiro–Wilk p) | Endpoint (Mean ± SD) (Min–Max) | (Shapiro–Wilk p) | ||
| LPS [EU/L] | 508.53 ± 89.83 382.19–594.96 | 0.4817 | 636.34 ± 136.85 401.08–871.60 | 0.4620 | 0.1210 |
| AEA [ng/mL] | 252.83 ± 41.75 186.22–283.97 | 0.1123 | 306.67 ± 65.45 192.98–389.58 | 0.4332 | 0.0831 |
| TNF-α [pg/mL] | 97.81 ± 17.51 68.48–115.42 | 0.1965 | 113.04 ± 22.80 76.75–146.56 | 0.7815 | 0.2144 |
| IL-1β [pg/mL] | 1346.94 ± 209.28 1001.15–1509.79 | 0.1242 | 1580.40 ± 325.51 1077.65–2183.22 | 0.8382 | 0.1717 |
| IL-10 [pg/mL] | 556.20 ± 31.95 500.76–583.24 | 0.0387 | 540.40 ± 81.99 395.87–663.44 | 0.9436 | 1.0000 * |
| Mean | Probiotic Group (n = 5) | Placebo Group (n = 10) | ||
|---|---|---|---|---|
| Delta | Paired t-Test (t, p) | Delta | Paired t-Test (t; p) | |
| LPS [EU/L] | 3.48 | 0.0853; 0.9361 | 56.98 | 2.0605; 0.0694 |
| AEA [ng/mL] | −1.11 | −0.0616; 0.9538 | 14.08 | 0.7457; 0.4749 |
| TNF-α [pg/mL] | 10.28 | 1.8887; 0.1320 | 7.15 | 1.2768; 0.2336 |
| IL-1β [pg/mL] | 46.52 | 0.5646; 0.6025 | 104.61 | 1.9067; 0.0889 |
| IL-10 [pg/mL] | 66.17 | 0.9439; 0.3452 * | 24.95 | 1.1912; 0.2640 |
| Probiotic Group (n = 5) Mean ± SD | Placebo Group (n = 10) Mean ± SD | |||||||
|---|---|---|---|---|---|---|---|---|
| Pre (Mean ± SD) (Min–Max) | Shapiro–Wilk p | Post (Mean ± SD) (Min–Max) | Paired t-Test (t; p) | Pre (Mean ± SD) (Min–Max) | Shapiro–Wilk p | Post (Mean ± SD) (Min–Max) | Paired t-Test (t; p) | |
| Active Coping [0–18] | 13 ± 4.53 8–18 | 0.3946 | 12 ± 3.94 9–18 | 0.9535; 0.3943 | 12.8 ± 3.71 6–18 | 0.5996 | 12 ± 3.97 4–17 | 0.7530; 0.4707 |
| Avoidance Behaviors [0–30] | 14.6 ± 2.61 13–19 | 0.0214 | 13 ± 5 6–20 | 0.3652; 0.7150 * | 14.8 ± 4.73 8–22 | 0.7620 | 13.2 ± 5.07 4–20 | 0.9130; 0.3850 |
| Seeking Support/Emotion-Focused Coping [0–36] | 16.4 ± 1.67 14–18 | 0.3140 | 16 ± 2.92 14–21 | 0.4313; 0.6885 | 16.7 ± 4.45 11–24 | 0.3767 | 15.5 ± 4.06 9–23 | 1.1078; 0.2967 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiącek, J.; Skonieczna-Żydecka, K.; Łoniewski, I.; Deli, C.K.; Fatouros, I.G.; Jamurtas, A.Z.; Moszczyńska, D.; Karolkiewicz, J. Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 Supplementation: An Exploratory, Randomized, Placebo-Controlled Trial of Endocannabinoid and Inflammatory Responses in Female Dancers. Microorganisms 2025, 13, 1284. https://doi.org/10.3390/microorganisms13061284
Wiącek J, Skonieczna-Żydecka K, Łoniewski I, Deli CK, Fatouros IG, Jamurtas AZ, Moszczyńska D, Karolkiewicz J. Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 Supplementation: An Exploratory, Randomized, Placebo-Controlled Trial of Endocannabinoid and Inflammatory Responses in Female Dancers. Microorganisms. 2025; 13(6):1284. https://doi.org/10.3390/microorganisms13061284
Chicago/Turabian StyleWiącek, Jakub, Karolina Skonieczna-Żydecka, Igor Łoniewski, Chariklia K. Deli, Ioannis G. Fatouros, Athanasios Z. Jamurtas, Dominika Moszczyńska, and Joanna Karolkiewicz. 2025. "Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 Supplementation: An Exploratory, Randomized, Placebo-Controlled Trial of Endocannabinoid and Inflammatory Responses in Female Dancers" Microorganisms 13, no. 6: 1284. https://doi.org/10.3390/microorganisms13061284
APA StyleWiącek, J., Skonieczna-Żydecka, K., Łoniewski, I., Deli, C. K., Fatouros, I. G., Jamurtas, A. Z., Moszczyńska, D., & Karolkiewicz, J. (2025). Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 Supplementation: An Exploratory, Randomized, Placebo-Controlled Trial of Endocannabinoid and Inflammatory Responses in Female Dancers. Microorganisms, 13(6), 1284. https://doi.org/10.3390/microorganisms13061284