Abstract
Pine wilt disease (PWD), caused by the pine wood nematode (PWN, Bursaphelenchus xylophilus), poses a significant threat to global pine forests and calls for the development of innovative management strategies. Microbial control emerges as an effective, cost-efficient, and environmentally sustainable approach to eliminate the damage from PWD. This review consolidates molecular mechanisms in the microbiological control of PWD, which focus on three core strategies: microbial control activity against PWN, biological control of vector insects, and the enhancement of host tree resistance to nematode infections. The review thoroughly evaluates integrated control strategies in which microbial control is used in traditional management practices. Recent studies have pinpointed promising microbial agents for PWN control, such as nematophagous microorganisms, nematicidal metabolites, parasitic fungi that target vector insects, and microbes that boost plant resistance. In particular, the control potential of volatile organic compounds (VOCs) produced by microorganisms against PWN and the enhancement of pine resistance to PWN by microorganisms were emphasized. Moreover, we assessed the challenges and opportunities associated with the field application of microbiological control agents. We emphasized the feasibility of multi-strategy microbial integrated control, which provides a framework for future studies on microbial-based PWD control strategies.
1. Introduction
Pine wilt disease (PWD) caused by the pine wood nematode (PWN), Bursaphelenchus xylophilus, which causes the rapid death of infected pine trees within a few weeks, has been a threat to pine forests worldwide (Figure 1) [1,2,3]. As a devastating disease to pine, PWN has invaded large-scale pine forests in China, Japan, Korea, and Europe, resulting in severe economic and ecological losses [4,5,6,7]. In East Asia, PWN first attacked Japan in 1905 and spread to China in 1982 [5,8,9,10]. A targeted monitoring of PWN based on the PWD risk assessment framework showed that such pine forest disease has been distributed in the southern, eastern, northeastern, northwestern, and central regions of China, with unsaturated geographical distribution and rapid spread [11]. Furthermore, global climate change accelerates in PWN distribution at high-latitude regions [12].
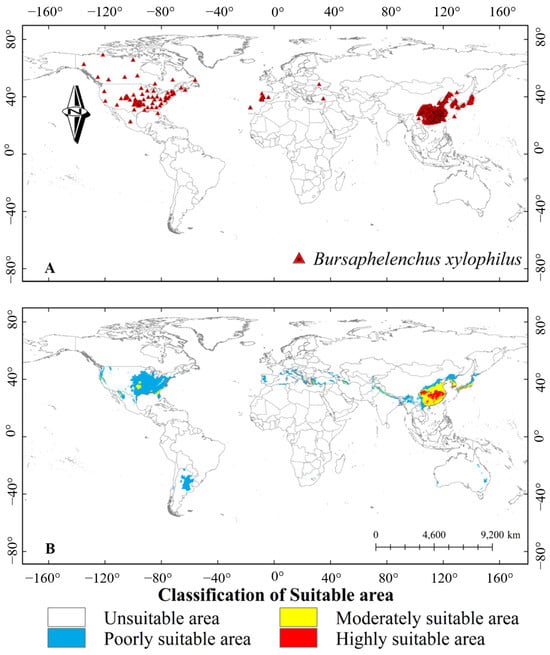
Figure 1.
The global distribution and predicted distribution of pine wood nematode (PWN). (A) Global geographic distribution of the occurrence points of PWN; (B) Predicted distribution of PWN suitable areas under current climatic conditions. Note: In the horizontal direction, the negative value is the west longitude, and the positive value is the east longitude; In the vertical direction, negative values are south latitudes and positive values are north latitudes. Adapted from Xiao et al. (2023) [3].
Currently, the control of PWD generally relies on a combination of physical and chemical measures. Physical measures, including fumigation of infected wood and black light trapping of vector insects, are highly expensive and labor-intensive [13]. The chemical agents, including avermectin, thiacloprid, milbemectin, alkaloids, terpenes, phenylpropanes, coumarins, and flavones [14,15], are rising concerns of serious environmental pollution [16]. Meanwhile, long-term application of chemical agents can induce resistance in PWN populations [17]. Biological control is an emerging approach that uses living organisms or their metabolites to reduce populations of PWN in pine forests [18], which is an emerging research hotspot [19,20]. Biological control measures, such as the application of insects like Scleroderma guani, have shown great effect in killing PWN [21,22]. Given their efficiency and environmental sustainability, the emerging microbial control measures demonstrate greater application potential due to their high diversity, rapid reproduction, and strong environmental adaptability [23,24].
The control of PWD using microorganisms primarily through three approaches: biological control of PWN populations, biological control of vector insects, and the enhancement of host tree resistance to nematode infections [24]. For instance, several biocontrol microorganisms showed high potential in the biological control of PWD due to their production of diverse nematicidal metabolites, which have been successfully developed into biological control agents (BCAs) [25]. Meanwhile, a crucial approach to PWN management involves controlling its primary vector, Monochamus alternatus (Coleoptera: Cerambycidae), which facilitates rapid PWN dissemination throughout pine forest ecosystems [26,27]. Compared to conventional biological control strategies, such as the release of natural enemies of M. alternatus, the application of entomopathogenic fungi, such as Metarhizium spp. and Beauveria bassiana, has emerged as an especially promising approach [22,28,29]. Furthermore, microorganism-mediated enhancement of pine resistance represents an emerging control strategy, operating through both constitutive improvement of host defenses and induced systemic resistance mechanisms. While the molecular and physiological mechanisms underlying these protective effects remain incompletely characterized, this approach holds considerable potential for integrated PWN management strategies.
To understand the molecular interactions between PWD and biological control microorganisms, we comprehensively reviewed the molecular mechanisms of microbial control of PWN through three fundamental aspects. First, we examined how microorganisms control the PWN population through two primary pathways: physical attack mechanisms and the production of nematicidal substances, with particular emphasis on volatile organic compounds (VOCs). Second, we summarized the interactions between microorganisms and PWN vectors, exploring how microbial agents can effectively control these vectors to prevent PWN transmission. Third, we evaluated how beneficial microorganisms can enhance pine tree resistance to PWN infection. We discussed these mechanisms in detail, with special attention to the molecular and biochemical pathways through which microorganisms induce enhanced plant resistance. Our work provides a new vision for developing an integrated biological control strategy against PWD.
2. Methodology
This study adopts the systematic literature review approach to investigate the molecular mechanisms of microbial-based control of PWD. The relevant keywords for literature search included “pine wilt disease”, “Bursaphelenchus xylophilus”, “pine wood nematode”, “biocontrol microorganisms”, “nematicidal metabolites”, “molecular mechanism”, “nematode-trapping fungi”, “endoparasitic fungi”, “toxigenic fungi”, “Monochamus alternatus”, “Metarhizium”, “Beauveria”, “vector insects”, “pine resistance”, “field application” alone and in combinations. The search engines included PubMed, Google Scholar, and CNKI. The search spanned from the establishment of each database up to 2024. Inclusion criteria were (1) the research clearly involves the biological control effects of microorganisms on PWN, vector insects, or pine trees; (2) provision of detailed molecular and biochemical mechanisms; (3) including experimental verification. Exclusion criteria were (1) literature with errors, incomplete data, or unavailable for analysis; (2) literature with inaccessible full text.
3. Molecular Mechanisms of Microbiological Control of PWN
A diverse array of microorganisms with potent nematophagous activity against PWN has been identified and characterized as promising biocontrol agents. These can be systematically classified into three major categories based on their control mechanisms: nematode-trapping fungi, which physically capture and kill nematodes; endoparasitic fungi, which parasitize nematodes; and toxigenic fungi, which produce compounds lethal to nematodes [24]. The specific microorganisms with demonstrated effects against PWN are listed in Table S1, providing a foundation for understanding potential biological control strategies.
3.1. Nematode-Trapping Microorganisms
Nematode-trapping fungi were isolated from pine-growing regions, such as Yunnan (China) and Choengdo City (Korea) [30,31]. These fungi are distinguished by their unique nematode-trapping structures, which mark their evolutionary transition from decomposers to predators. The trapping mechanisms can be categorized into two main types: constricting rings and adhesive structures (including adhesive knobs and three-dimensional adhesive networks). The fungal species, such as Drechslerella dactyloides and Arthrobotrys dactyloides, form constricting rings that effectively immobilize PWN [31,32]. Other fungi, including Arthrobotrys cladodes and Arthrobotrys oviformis, develop three-dimensional adhesive networks that bind to the nematode, facilitating subsequent infection and internal colonization [32]. Other fungi exhibiting these trapping capabilities include Arthrobotrys, Dactylella, and Monacrosporium. Recent research has demonstrated the effect of these fungi as biocontrol agents; for instance, Volutella citrinella GUCC2219 achieved a 33% predation rate in vitro, while its fermentation broth induced complete nematode mortality within 72 h [33].
The predation process of nematode-trapping fungi has been extensively studied. In the initial stages, these fungi produce VOCs, such as 2,4-dithiapentane, S-methyl thioacetate, and dimethyl disulfide, which mimic nematode pheromones to attract their prey [34]. Once attracted, the nematodes are recognized by the fungus Arthrobotrys oligospora through lectin–carbohydrate interactions mediated by lectins produced by the fungus [35]. Following recognition, the fungi initiate the formation of trapping structures to capture the nematodes. This process is regulated by in vivo signals [36]. Upon detecting nematodes, specific signaling pathways, including the G protein signaling pathway and Ca2+/calmodulin-dependent protein kinases (CaMKs), trigger the development of these structures [36]. The G protein pathway plays a critical role in trap formation in A. oligospora; for example, mutants lacking the G protein signal regulator Ric8 are unable to form traps [37]. Similarly, CaMKs, a class of serine/threonine (Ser/Thr) kinases, contribute to various regulatory cascades within organisms [38]. In the ΔCaMK B mutant, where the CaMK B gene is knocked out, the formation of trapping structures is significantly delayed [39]. Once the fungi capture or adhere to the PWN, they secrete a suite of extracellular enzymes, including chitinase, serine protease Ac1, and collagenase. These enzymes act synergistically to degrade the nematode cuticle, facilitating fungal penetration and colonization [30,40,41]. Subsequently, A. oligospora penetrates the nematode via penetration tubes, exerting a mechanical force that causes the cuticle to indent and eventually breach [42]. In the final stage, the penetration tube develops into a nutrient mycelium at the infection site, which colonizes and digests the nematode, leading to its death [42,43].
The molecular studies suggested that the efficacy of nematode-trapping fungi relies on their secretion of various hydrolases, with serine protease identified as a particularly crucial enzyme in the trapping process [30,44]. A notable example is Serratia sp. A88copa13, isolated from Portugal, demonstrates high nematicidal activity through the production of a 70 kDa serine protease that actively degrades PWN components [44]. Similarly, Arthrobotrys conoides secretes an extracellular serine protease (Ac1) that immobilizes free-living PWN, showing 40–50% immobilization rates after 24 h of exposure [30].
3.2. Nematophagous Microorganisms
Nematophagous fungi represent a key group of microorganisms that control PWN through spore-based infection [45]. Unlike nematode-trapping fungi that rely on external capture mechanisms, endoparasitic fungi directly colonize the cavity of nematodes and result in rapid death, making them particularly effective as biocontrol agents. Esteya vermicola, an endoparasitic fungus demonstrating efficient infectivity against PWN [46]. This discovery marked a crucial advancement in biological control strategies against pine wilt disease, as E. vermicola exhibited both high specificity and pathogenicity toward PWN.
The infection mechanism of E. vermicola follows a sophisticated process. The fungus initially attracts PWN by producing specific volatile organic compounds (VOCs), including α-pinene, β-pinene, and camphor, which mimic pine tree scents [47]. These compounds were proved to be more attractive to PWN than those from actual pine trees [48]. Once PWN is attracted, E. vermicola initiates a four-day infection cycle [49]. The process begins when lunate conidia adhere to the cuticle of PWN without impeding its movement, enabling infected nematodes to disseminate the fungus throughout the host tree [50]. Within 18–24 h of adhesion, the conidia germinate and penetrate the cuticle of PWN [49] and then consume the internal contents of PWN, progressively reducing its motility and destroying its organs and tissues until death occurs [50]. The cycle concludes as fungal mycelium breaches the cuticle of a dead nematode, producing new lunate conidia that can attract and infect additional PWN in the next cycle [50].
Recent research has expanded our understanding of Esteya spp. and its infection mechanisms. Yin et al. (2020) demonstrated that E. vermicola blastospores exhibit comparable biocontrol efficacy to PWN [51]. Moreover, several strains of E. vermicola have been identified, including ATCC74485, CBS115803, CNU120806, and NKF13222. While Esteya spp. remained monotypic for many years after its discovery [46], Li et al. (2021) identified a second species, Esteya floridanum, which shares similar morphological characteristics and infection mechanisms with E. vermicola, demonstrating comparable nematicidal activity [52].
3.3. Toxigenic Microorganisms
Toxigenic microorganisms show significant potential in biocontrol against PWN due to their ability to produce nematicidal compounds, leading to rapid and effective nematode mortality. These organisms, including various species of fungi, bacteria, and actinomycetes, synthesize diverse classes of nematicidal metabolites, ranging from low-molecular-weight metabolites and volatile organic compounds (VOCs) to complex secondary metabolites and enzymes (Table 1). We specifically reviewed the toxigenic microorganisms identified to date and examined their specific nematicidal compounds, mechanisms of toxicity, and control efficiency against PWN, with particular emphasis on their potential for practical application in pine wilt disease management.
3.3.1. Nematicidal Proteins from Toxigenic Microorganisms
Among toxigenic bacteria, the genus Bacillus has emerged as a source of nematicidal compounds against PWN, producing diverse bioactive molecules, including crystal proteins, guanidine compounds, and alkaline proteases [53,54,55]. Bacillus thuringiensis produces crystal protein App6Aa2, which demonstrates significant toxicity against PWN (LC50 = 49.71 μg·mL−1) by inducing shrinkage and thinning of intestinal cells, contraction of the intestine from the body wall, vacuolization, and the degeneration of appearance [53]. Similarly, Bacillus LYMC-3 synthesizes guanidine compounds with notable nematicidal activity, achieving LC50 values of 113.5 mg·L−1 and 62.5 mg·L−1 at 24 and 48 h, respectively [54]. Recent studies have shown that extracellular proteases from Bacillus cereus NJSZ-13 can degrade the nematode cuticle, progressively diminishing nematode activity [56]. Beyond natural isolates, genetic engineering approaches have yielded promising results, as demonstrated by the engineered strain Bxy19P3GFP, which expresses the Cry6Aa crystal protein and has shown significant nematicidal efficacy in greenhouse trials [57].
Recent studies have identified a novel nematotoxic cytolytic-like protein (CytCo) from the entomopathogenic fungus Conidiobolus obscurus that specifically targets the lipid metabolism of PWN through a distinct molecular mechanism. This protein disrupts the formation and maintenance of lipid droplets, which are crucial cellular organelles governing lipid homeostasis during PWN’s infection cycle. At the molecular level, CytCo operates through dual mechanisms: it interferes with surface phospholipid organization and modulates the expression of key genes involved in lipid metabolism. These combined effects lead to the elimination of large lipid droplets and subsequent disruption of lipid distribution patterns within PWN cells. While the immediate effects of CytCo on lipid metabolism are well-characterized, its broader implications for PWN’s infection cycle and potential applications in biocontrol strategies remain unclear [58]. Moreover, Actinomycetes, particularly Streptomyces species, produce diverse nematicidal compounds that show high toxicity against PWN. A prime example is teleocidin B4, isolated from Streptomyces sp. 680560, which achieves 95% mortality within 48 h while simultaneously inhibiting egg hatching of PWN [59]. The practical potential of these compounds is further evidenced by Streptomyces strain AE170020, whose extracts demonstrate complete PWN control at a minimal concentration of 7.2 mg per pine tree [60].
3.3.2. Nematicidal Metabolites from Toxigenic Microorganisms
Toxigenic microorganisms synthesize diverse secondary metabolites that exhibit significant nematicidal activity against PWN. These nematicidal substances are classified into two primary categories based on their physicochemical properties: non-volatile metabolites and volatile organic compounds (VOCs). Previous investigations have led to the identification and characterization of 91 distinct nematicidal substances, comprising 54 non-volatile metabolites and 37 VOCs (Table 1). These compounds encompass multiple structural classes, including polyketones, lipopeptides, quinones, alkaloids, piperazines, phenols, terpenes, aldehydes, siderophores, and furans.

Table 1.
Nematicidal metabolites against pine wood nematode secreted by microorganisms.
Table 1.
Nematicidal metabolites against pine wood nematode secreted by microorganisms.
| Strain | Substance Class | Bioactive Substance | Reference |
|---|---|---|---|
| Bacteria | |||
| Brevundimonas diminuta LCB-3 | Alcohols | (R)-(-)-2-ethylhexan-1-ol | [61] |
| Serratia marcescens AHPC29 | Alkaloids | salsolinol | [59] |
| Bacillus sp. SMrs28 | Alkenes | 5,8-triene | [62] |
| Streptomyces sp. AN091965 | Antibiotics | Spectinabilin | [63] |
| Streptomyces ahygroscopicus | Antibiotics | tetramycin B3 | [64] |
| Streptomyces sp. AE170020 | Aromatic Compounds | alloaureothin | [60] |
| Streptomyces sp. 680560 | Aromatic Compounds | Teleocidin B4 | [59] |
| Bacillus sp. SMrs28 | Aromatic Compounds | phenylacetamide | [62] |
| Lysinimonas M4 | Aromatic Compounds | 2-coumaranone | [65] |
| Streptomyces sp. AE170020 | Benzopyranones | Aureothin | [60] |
| Bacillus sp. SMrs28 | Cyclic Compounds | 4-Oxabicyclo [3.2.2] nona-1 | [62] |
| Stenotrophomonas maltophilia G2 | Enzymes | serine protease | [66] |
| Bacillus sp. SMrs28 | Ester | methyl elaidate | [62] |
| Bacillus sp. SMrs28 | Fatty Acids | lauric acid | [62] |
| Bacillus sp. SMrs28 | Ketones | 4-dione | [62] |
| Streptomyces avermitilis AVE-H39 | Lactone | 13α-Hydroxymilbemycinβ13 | [67] |
| Streptomyces avermitilis AVE-H39 | Lactone | 26-methyl-13α-hydroxymilbemycin β13 | [67] |
| Bacillus pumilus LYMC-3 | Nitrogen Compounds | 2-{3-[(3S,8aS)-1,4-dioxooctahydropyrrolo [1,2-a] pyrazin-3-yl] propyl} guanidine | [54] |
| Bacillus sp. SMrs28 | Nitrogen Compounds | (3S, 8aS)-hexahydro-3methylpyrro [1,2-a] pyrazine-1 | [62] |
| Bacillus amyloliquefaciens JK-JS3 | Nitrogen Compounds | 2,2-dimethyl-N-phenylpropanethioamide | [68] |
| Bacillus amyloliquefaciens JK-JS3 | Nitrogen Compounds | Hexahydro-5-methyl-1-phenyl-1,3,5-triazine-2-thione | [68] |
| Bacillus amyloliquefaciens JK-JS3 | Nitrogen Compounds | [(4,7,7-trimethyl-3-bicyclo [2.2.1] heptanylidene) amino] urea | [68] |
| Streptomyces sp. C611 | Nitrogen Compounds | Furaltadone | [69] |
| Bacillus sp. SMrs28 | Peptides | cyclo(L-Pro-L-Val) | [62] |
| Lysinimonas M4 | Peptides | cyclo-(Phe-Pro) | [65] |
| Erwinia sp. A41C3 | Siderophores | Catecholate-typesiderophore | [70] |
| Rouxiella sp. Arv20#4.1 | Siderophores | hydroxamate-type siderophore | [70] |
| Streptomyces sp. TCS19-048 | Sulfur compounds | S-3-1 | [71] |
| Pseudoduganella violaceinigra G5-3 | VOCs | 2,5-dimethyl pyrazine | [72] |
| Pseudoduganella violaceinigra G5-3 | VOCs | 4-dimethylaminopyridine | [72] |
| Pseudoduganella violaceinigra G5-3 | VOCs | benzyl acetate | [72] |
| Pseudoduganella violaceinigra G5-3 | VOCs | phenethyl butyrate | [72] |
| Pseudoduganella violaceinigra G5-3 | VOCs | phenethyl alcohol | [72] |
| Stenotrophomonas maltophilia | VOCs | phenol | [73] |
| Bacillus subtilis | VOCs | 2-octanol | [73] |
| Serratia marcescens | VOCs | benzaldchyde | [73] |
| Stenotrophomonas maltophilia | VOCs | benzeneacetaldehyde | [73] |
| Bacillus subtilis | VOCs | decanal | [73] |
| Bacillus subtilis | VOCs | 2-nonanone | [73] |
| Stenotrophomonas maltophilia | VOCs | 2-undecanone | [73] |
| Bacillus subtilis | VOCs | cyclohexene | [73] |
| Stenotrophomonas maltophilia | VOCs | dimethyl disulfide | [73] |
| Vibrio atlanticus S-16 and Pseudoalteromonas marina H-42 | VOCs | dimethyl disulfide | [74] |
| Vibrio atlanticus S-16 and Pseudoalteromonas marina H-42 | VOCs | benzaldehyde | [74] |
| Vibrio atlanticus S-16 and Pseudoalteromonas marina H-42 | VOCs | dimethyl trisulfide | [74] |
| Vibrio atlanticus S-16 | VOCs | tert-butylamine | [74] |
| Vibrio atlanticus S-16 | VOCs | acetone | [74] |
| Pseudoalteromonas marina H-42 | VOCs | Dimethylamine | [74] |
| Pseudoalteromonas marina H-42 | VOCs | N(diisopropylphosphino)methyl- | [74] |
| Fungi | |||
| Geotrichum sp. AL4 | Alcohols | [2,3-dihydro-2-(1-methylethenyl)-1-benzofuran-5-yl] methanol | [75] |
| Alternaria sp. Samif01 | Aromatic Compounds | Alternariol 9-methyl ether | [76] |
| Aspergillus fumigatus | Aromatic Compounds | Fumiquinones A and B | [25] |
| Caryospora callicarpa YMF1.01026 | Aromatic Compounds | 4,8-Dihydroxy-3,4-dihydronaphthalen-1(2H)-one | [77] |
| Caryospora callicarpa YMF1.01026 | Aromatic Compounds | 4,6-dihydroxy-3,4-dihydronaphthalen-1(2H)-one | [77] |
| Caryospora callicarpa YMF1.01026 | Aromatic Compounds | 4,6,8-trihydroxy-3,4-dihydronaphthalen-1(2H)-one) | [77] |
| Caryospora callicarpa YMF1.01026 | Aromatic Compounds | 3,4,6,8-tetrahydroxy-3,4-dihydronaphthalen-1(2H)-one(cis-4-hydroxyscytalone) | [77] |
| Oidiodendron sp. | Aromatic Compounds | 4-Hydroxyphenylacetic acid | [78] |
| Gliocladium roseum YMF1.00133 | Aromatic Compounds | 5-n-heneicosylresorcinol | [79] |
| Geotrichum sp. AL4 | Aromatic Compounds | 1-(2,4-dihydroxyphenyl) ethanone | [75] |
| Caryospora callicarpa YMF1.01026 | Aromatic Compounds | caryospomycins A–C | [80,81] |
| Coelomycetes sp. YMFl.01029 | Aromatic Compounds | Preussomerin C | [82] |
| Coelomycetes sp. YMFl.01029 | Aromatic Compounds | preussomerin E | [82] |
| Coelomycetes sp. YMFl.01029 | Aromatic Compounds | preussomerin D | [82,83] |
| Coelomycetes sp. YMFl.01029 | Aromatic Compounds | 4,6,8-trjhydfoxy-3,4-dihydronaphthalen-1(2H)-one | [82,83] |
| Coelomycetes sp. YMFl.01029 | Aromatic Compounds | (4RS)4,8-dihydroxy-3,4-dihydronaphthalen-1(2H)-one | [82,83] |
| Chaetomium ascotrichoides 1-24-2 | Aromatic Compounds | 4,5,6-trihydroxy-7-methylphthalide | [84] |
| Chaetomium ascotrichoides 1-24-2 | Aromatic Compounds | 2-chlorobenzothiazole | [84] |
| Fusarium oxysporum EF119 | Benzopyranones | Bikaverin | [85] |
| Aspergillus sp. | Carboxylic Acids | 5-Hydroxymethyl-2-furoic acid | [86] |
| Fusarium oxysporum EF119 | Carboxylic Acids | fusaric acid | [85] |
| Pseudohalonectria adversaria YMF1.01019 | Cyclic Compounds | pseudohalonectrin A and B | [80] |
| Fusarium bulbicola | Cyclic Esters | Beauvericin | [87] |
| Beauveria bassiana and Beauveria pseudobassiana | Cyclic Esters | Beauvericin | [87] |
| Paraniesslia sp. YMF1.01400 | Glycosides | (2S,2‘R,3R,3′E,4E,8E)-1-O-(β-D-glucopyranosyl)-3-hydroxyl-2-[N-2′-hydroxyl-3′-eicosadecenoyl] amino-9-methyl-4,8-octadecadiene | [80] |
| Oidiodendron sp. | Lactone | oidiolactone D | [78] |
| Ophioceras dolichostomum YMF1.00988 | Lipids | Ophiocerol | [88] |
| Geotrichum sp. AL4 | Nitrogen Compounds | 1-[(2R*,4S*,5S*)-2-chloro-4-methyl-1,3-oxazinan-5-yl] ethenone | [75] |
| Chaetomium ascotrichoides 1-24-2 | Nitrogen Compounds | O-methylisourea | [84] |
| Gliocladium roseum YMF1.00133 | Peptides | Gliocladin C | [79] |
| Gliocladium roseum 1A | Peptides | Gliocladines A–D | [89] |
| Trichoderma sp. | VOCs | 1β-vinylcyclopentane-1α,3α-diol | [83,90] |
| Trichoderma sp. | VOCs | 6-pentyl-2H-pyran-2-one (2) | [83,90] |
| Annulohypoxylon sp. FPYF3050 | VOCs | 1,8-cineole | [91] |
| Annulohypoxylon sp. FPYF3050 | VOCs | (+)-sativene | [91] |
| Annulohypoxylon sp. FPYF3050 | VOCs | isocaryophyllene | [91] |
The discovery of non-volatile nematicidal metabolites has advanced considerably since the initial characterization of aliphatic extracts from Pseudohalonectria adversaria, Xylaria sp., and Hyphomycete sp. exhibiting nematicidal activity against PWN [92]. Subsequent investigations led to the isolation and structural determination of increasingly potent compounds. Two naphthoquinone derivatives, Fumiquinones A and B, isolated from Aspergillus fumigatus, demonstrated nematicidal activities of 24% and 44%, respectively [25]. A significant advancement in this field was the identification of (R)-(-)-2-ethylhexan-1-ol from Brevundimonas diminuta, which exhibited 73% mortality against PWN in a 48 h bioassay [61]. Further structural diversity was revealed through the isolation of Beauvericin, a cyclohexadepsipeptide from Fusarium bulbicola, which demonstrated substantial nematicidal efficacy [93]. Recently, more efficient nematicidal substances have been identified, such as beauvericin, salsolinol, O-methylisourea, 2-chlorobenzothiazole, 4,5,6-trihydroxy-7-methylphthalide, and tetramycin B3 [64,84,87,94], showed high control efficiency against PWN.
VOCs have emerged as particularly efficacious nematicidal agents, consistently demonstrating promising nematicidal activity compared to non-volatile metabolites, with representative compounds including 2,5-dimethylpyrazine, 4-dimethylaminopyridine, and 1,8-cineole (Table 1). Quantitative bioassays have revealed the exceptional efficacy of bacterial VOCs, with metabolites from Pseudoduganella violaceinigra G5-3 and Novosphingobium pokkalii G8-2 achieving mortality rates of 98.26% and 93.10%, respectively, within 24 h of exposure [72]. Marine microorganisms are known to produce nematicidal VOCs, such as Pseudoalteromonas marina strain H-42 and Vibrio atlanticus strain S-16, which produce nematicidal compounds, including dimethyl trisulphide, benzaldehyde, dimethyl disulphide, and tert-butylamine [72]. Beyond direct nematicidal activity, certain VOCs exhibit developmental inhibition properties, as demonstrated by 1,8-cineole produced by Annulohypoxylon sp. FPYF3050, which significantly impairs PWN embryogenesis [91].
4. Molecular Mechanisms of Microbial Control of Vector Insects
Monochamus alternatus is the primary vector insect of PWN, spreading the nematode by feeding and laying eggs in healthy pine trees [95]. Several microorganisms, especially entomopathogenic fungi such as Metarhizium and Beauveria species, have shown significant potential as BCAs against M. alternatus [96]. This section examines the diverse microorganisms that show promise as BCAs and elucidates their infection mechanisms against M. alternatus.
4.1. Biological Control Agents of M. alternatus
Entomopathogenic fungi, particularly species of Metarhizium and Beauveria, have emerged as efficacious biological control agents against PWN due to their high virulence and demonstrated mortality rates (Table 2). Notably, B. pseudobassiana exhibited 100% mortality against Monochamus galloprovincialis, another PWN vector, at a concentration of 1 × 108 conidia/mL [97]. Recent investigations by Zheng et al. (2024) revealed that M. robertsii GQH6, isolated from the Loess Plateau, achieved 100% corrected mortality against M. alternatus at concentrations of 108 and 109 conidia/mL [98]. Additionally, several other microorganisms have demonstrated promising potential as biological control agents against M. alternatus (Table 2). These include Serratia marcescens AHPC29, S. nematodiphila ZJPC33, Lecanicillium decadeum, Aspergillus austwickii, Scopulariopsis alboflavescens, Aspergillus ruber, Beauveria bassiana, Penicillium citrinum, and Trichoderma dorotheae [99,100]. These organisms have shown significant entomopathogenic activity while maintaining low phytotoxicity against host pine trees.
Climatic factors, such as temperature, humidity, and latitude, determine the distribution of Metarhizium and Beauveria and the pathogenicity effect on the M. alternatus [101]. Temperature has a significant impact on the efficacy of Metarhizium and Beauveria as BCAs [102]. Omuse et al. (2022) established a nonlinear model to simulate the effects of a wide temperature range on the conidial germination and mycelial growth of these fungi, identifying the optimal temperature for their pathogenicity and safety properties. Their results indicated that all fungi germinated best at 26.6–28.1 °C and grew best at 25.9–28.1 °C. These conditions are applicable to the management of most pests in tropical regions [103]. Therefore, establishing models to predict the adaptive growth temperatures of entomopathogenic fungi can help in selecting the most suitable fungi for different temperature regions. McGuire et al. (2022) proposed a qualitative conceptual model to study how latitude influences the specificity of fungi. In high-latitude regions, temperature is the most significant abiotic stressor, and fungi in these areas exhibit stronger environmental adaptability due to broader climate fluctuations. In contrast, fungi in low-latitude regions face greater selection pressure from host insects, making them more host-specific [104].
Under laboratory conditions, B. bassiana and M. anisopliae showed a great effect against M. alternatus. However, field conditions, influenced by environmental factors such as temperature and humidity, require evaluation of their stability in the control of PWN [105]. Notably, the B. bassiana ERL836 fungal powder demonstrated high nematicidal activity in the field. Compared to suspension sprays, which require large amounts of water, the fungal powder has a lower demand and is more applicable [106]. Furthermore, although Kim et al. (2022) suggest that the risk of B. bassiana ERL836-infected M. alternatus adults spreading the fungus to other non-target insects is low, the impact of B. bassiana and M. anisopliae on non-target insects and forest ecosystem security should still be taken seriously [106].
Table 2.
Biocontrol microorganisms against vector insect, Monochamus alternatus.
Table 2.
Biocontrol microorganisms against vector insect, Monochamus alternatus.
| Strain | Bioactive Substance | Killed Insects | Source | Reference |
|---|---|---|---|---|
| B. bassiana | N/A | Monochamus alternatus | Monochamus alternatus | [100,107] |
| B. bassiana F-263 | N/A | Monochamus alternatus | Monochamus alternatus | [108,109,110] |
| B. bassiana ERL836 | N/A | Monochamus alternatus | Entomology Research Laboratory, University of Vermont, USA | [106] |
| B. bassiana B7/B9 | N/A | Monochamus alternatus | Monochamus alternatus | [111] |
| B. brongniartii F-877 | N/A | Monochamus alternatus | Monochamus alternatus | [108] |
| B. brongniartii #879 | N/A | Monochamus alternatus | Psacothea hilaris | [108] |
| B. pseudobassiana | N/A | Monochamus galloprovincialis | Monochamus galloprovincialis | [97] |
| M. anisopliae JEF-279 | Destruxin and protease | Monochamus alternatus | Soil | [112] |
| M. anisopliae 1291 | N/A | Monochamus alternatus | M. alternatus larva | [113] |
| M. anisopliae 1349 | N/A | Monochamus alternatus | M. alternatus adult | [113] |
| M. anisopliae 2049 | N/A | Monochamus alternatus | Cydnid bug adult | [113] |
| M. anisopliae JEF-197 | N/A | Monochamus alternatus | Soil | [114] |
| M. anisopliae JEF-271 | N/A | Monochamus alternatus | Soil | [114] |
| M. anisopliae JEF-279 | N/A | Monochamus alternatus | Soil | [114] |
| M. anisopliae Ma789 | N/A | Monochamus alternatus | Chinese Academy of Forestry | [115] |
| M. anisopliae MaYTTR-03 | N/A | Monochamus alternatus | Soil | [116] |
| M. anisopliae MaYTTR-04 | N/A | Monochamus alternatus | Soil | [116] |
| M. anisopliae MaZPTR-01 | N/A | Monochamus alternatus | Soil | [116] |
| M. anisopliae var. anisopliae | N/A | Monochamus alternatus | Monochamus alternatus | [107] |
| M. anisopliae var. major CQMa117 | N/A | Monochamus alternatus | Monochamus alternatus | [117] |
| M. robertsii GQH6 | N/A | Monochamus alternatus | Soil | [98] |
| Aspergillus austwickii | N/A | Monochamus alternatus | Monochamus alternatus | [100] |
| Aspergillus ruber | N/A | Monochamus alternatus | Monochamus alternatus | [100] |
| Bacillus thuringiensis Cry3Aa | Coleopteran-specific Cry3Aa toxin | Monochamus alternatus | Not mentioned | [118] |
| Lecanicillium attenuatum | N/A | Monochamus alternatus | Monochamus alternatus | [100] |
| Paecilomyces farinosus | N/A | Monochamus alternatus | Monochamus alternatus | [107] |
| Penicillium citrinum | N/A | Monochamus alternatus | Monochamus alternatus | [100] |
| Scopulariopsis alboflavescens | N/A | Monochamus alternatus | Monochamus alternatus | [100] |
| Serratia marcescens | N/A | Monochamus alternatus | Monochamus alternatus | [107] |
| Serratia marcescens AHPC29 | N/A | M. alternatus and M. saltuarius | M. alternatus and M. saltuarius | [99] |
| Trichoderma dorotheae | N/A | Monochamus alternatus | Monochamus alternatus | [100] |
“N/A”: Not Applicable.
4.2. Infection Mechanisms of Metarhizium and Beauveria Against M. alternatus
The molecular mechanisms of the infection process of entomopathogenic fungi against M. alternatus have been well investigated and follow a complex, sequential pathway comprising six distinct stages: (1) conidial adhesion to the insect cuticle followed by germination; (2) enzymatic and mechanical penetration of the host cuticle; (3) proliferation of fungal hyphae within the insect cuticle; (4) production and secretion of insecticidal metabolites; (5) modulation of and response to host immune defenses; (6) fungal colonization culminating in hyphal extrusion and conidiation on the insect cadaver [119,120].
Initial colonization is mediated by specific fungal proteins that facilitate conidial attachment to the host cuticle. Fungal proteins, such as CFEM and Mad1, enable conidia to adhere to the host’s cuticle [121,122]. Mad1 also regulates cytoskeletal organization and activates genes related to the cell cycle [122]. Once attached, conidia produce germ tubes that develop into appressoria, structures enriched with organelles, preparing them to penetrate the host’s integument [123]. Cuticle penetration combines mechanical force and enzymatic breakdown [120]. Fungi like Metarhizium and Beauveria generate pressure through appressoria and secrete hydrolases, including proteases, chitinases, and lipases, to degrade the cuticle’s chitin, proteins, and lipids [119,124]. For instance, M. anisopliae employs the protease Pr1 to dismantle the cuticle of M. alternatus [125,126]. Other factors, such as MrGpa1 in M. robertsii and CYP52X1 in B. bassiana, further improve penetration efficiency [127,128].
Upon reaching the hemocoel, hyphae transform into yeast-like blastospores with a brush-like outer layer [129]. This adaptation helps the fungus evade the host’s immune defenses and spread rapidly within the hemolymph [119]. Fungi produce insecticidal metabolites, such as destruxins (DTXs) and proteases, which kill the host by inducing flaccid paralysis and disrupting the synthesis of nucleic acids and proteins [130,131]. Research by Kim et al. (2020) shows that DTXs and proteases from M. anisopliae strain JEF-279 cause flaccid paralysis in infected hosts [126]. In response, infected insects increase production of actin and tropomyosin, potentially as a defense against DTXs, with exposure to these metabolites leading to tetanic paralysis followed by flaccid paralysis [126]. Fungal infection prompts immune responses in M. alternatus, driven by the Toll and IMD pathways, which trigger antimicrobial peptide (AMP) production [132,133,134,135]. The reactive oxygen species (ROS) pathway also bolsters defense [132]. Additionally, bacteria such as Pseudomonas and Serratia associated with the insect inhibit B. bassiana by suppressing conidial germination and growth [136].
To counter these defenses, fungi employ strategies like immune evasion and enzyme secretion. For example, MCL1 in M. anisopliae conceals β-glucans on the fungal cell wall, reducing recognition by hemocytes [137]. The MaAC gene in the cAMP signaling pathway enhances fungal tolerance to oxidative and osmotic stress from the host’s immune system [138,139], while Ras GTPase (Ras3) and adenylate cyclase further support stress resistance [140,141]. When the host’s nutrients are exhausted, and death occurs, blastospores revert to hyphal growth and exit the cuticle, a process triggered by favorable environmental cues detected through G protein-mediated signaling [119]. The fungus then emerges, generates new conidia, and begins a new infection cycle [126].
5. Molecular Mechanisms of Microbial Enhancement of Pine Resistance Against PWN
Induced resistance (IR) is a phenomenon whereby plants, after appropriate stimulation, exhibit enhanced resistance to subsequent pathogen challenges [142]. This concept, first raised by Ross in the 1960s [143,144], primarily includes two distinct but interrelated defense mechanisms: systemic acquired resistance (SAR) and induced systemic resistance (ISR) [145]. SAR constitutes a defense mechanism activated through localized induction by virulent, avirulent, or non-pathogenic microbes, thereby enhancing the plant’s broad-spectrum defensive capacity against pathogens [145,146]. This process is characterized by elevated salicylic acid (SA) levels, a hormone central to immune responses against biotrophic pathogens [147], and concurrent activation of pathogenesis-related (PR) genes. These genes encode PR proteins with demonstrated antibacterial activity, which are functionally implicated in SAR-mediated defense [148]. In contrast, ISR is generally SA-independent and does not induce PR protein accumulation, distinguishing it from SAR [149]. ISR is primarily mediated by the phytohormones jasmonic acid (JA) and ethylene (ET) [145,150], functioning through enhancement of the host’s structural defenses rather than through direct pathogen killing or inhibition [146].
5.1. Mechanisms of Microorganisms Improve Pine SAR Against PWN
Pine trees exhibit enhanced resistance against PWN through several mechanisms regulated by microorganisms, phytohormones, secondary metabolites, oxidative stress responses, post-transcriptional regulation mediated by small RNAs, and the expression of defense-related genes [142,151]. The interaction between these factors contributes to the overall resilience of pine against pathogenic infections.
5.1.1. Improvement of SAR by Exogenous Microorganisms
Exogenous microorganisms play a pivotal role in bolstering pine resistance against PWN. Upon PWN invasion, certain molecular mechanisms were applied by microorganisms to enhance pine resistance, especially by regulating the expression of defense genes in host pine, including encoding cell wall hydrolases and chitinases, or by inducing the synthesis of nematicidal compounds and enzymes, such as ethylene, peroxidase, catalase, and polyphenol oxidase [152,153,154]. Following PWN invasion, ethylene levels in pine trees tend to increase, indicating their potential role in the development of PWD [155]. However, excessive accumulation of the ethylene precursor, 1-aminocyclopropane-1-carboxylate (ACC), can lead to plant damage and even mortality [152,156]. Nascimento et al. (2013) demonstrated that ACC deaminase produced by Pseudomonas putida strain UW4 can mitigate the harmful ethylene levels in PWN-infected pine seedlings, thereby slowing down disease progression [152]. Furthermore, Nunes da Silva et al. (2019) indicated that a biofertilizer comprising diazotrophic bacteria and the chitosan-producing fungus Cunninghamella elegans prevented declines in photosynthetic pigments and water content in infected Pinus pinaster, while enhancing phenolic synthesis in PWN-inoculated Pinus pinea [157].
5.1.2. Improvement of SAR by Endophytes
Endophytes associated with pine have demonstrated significant nematicidal activity and contribute to enhanced defensive capabilities against PWN [158]. For example, Peng et al. reported that the pine endophytic strain Pseudomonas abietaniphila BHJ04 significantly enhanced the growth of branches and roots of Pinus massoniana while inhibiting the spread of PWD [153]. Specifically, inoculation with strain BHJ04 resulted in a 64.97% increase in shoot dry weight and a 38.31% increase in root dry weight compared to control plants. Additionally, inoculation with strain BHJ04 led to a notable increase in the expression of cell wall hydrolase genes, chitinase genes, and genes belonging to the cytochrome P450 family in P. massoniana. Sun et al. (2024) identified the nematicidal properties of the endophyte Bacillus velezensis Pt-RP9, noting increased activities of peroxidase, catalase, and polyphenol oxidase in pine seedlings treated with this endophyte [154]. Given that pine trees typically engage in symbiotic relationships with various fungi, the impact of these microorganisms on PWN infection is notably significant [159,160,161,162]. For instance, while fungi such as Aspergillus can exacerbate PWN infections [163,164,165], beneficial biocontrol strains like Curtobacterium pusillum and Pseudomonas putida improve the resistance of pine trees against PWN [152,166].
5.1.3. Improvement of SAR by Ectomycorrhizal Fungi
Ectomycorrhizal fungi (EMF) have also been implicated in enhancing resistance in pine trees against PWD. Nakashima et al. (2015) demonstrated that pine seedlings that formed highly abundant ectomycorrhizae displayed the highest survival and growth rates under PWN invasion, suggesting a beneficial role of EMF in conferring defense against PWN [167]. Notably, colonization rates of EMF in roots of PWN-infested trees were significantly reduced, indicating a potentially negative interaction between EMF colonization and PWN invasion [168]. Chu et al. (2019) identified various EMF species, including Suillus lactifluus, Handkea utriformis, Amanita vaginata, and Suillus laricinus, which enhanced host pine resistance against PWD [169]. Furthermore, since the pine rhizosphere microbiome is highly dependent on the growth of pine trees, PWN infection can severely damage the microbiome, leading to a decline in community diversity [149]. Chu et al. found that 9 months after inoculation with the ectomycorrhizal fungus Suillus bovinus, which significantly enhances pine resistance against PWN, the diversity and richness of the pine rhizosphere microbiome were significantly higher than in the control group [170]. Therefore, the long-term application of beneficial microorganisms may promote the growth of the pine rhizosphere microbiome by enhancing their resistance.
5.2. Mechanisms of Microorganisms Induce Pine ISR Against PWN
ISR is a plant-mediated resistance response primarily activated through ET and JA signaling pathways and is generally not associated with SA [171,172]. Certain rhizobacteria, like Pseudomonas fluorescens WCS417r, can trigger ISR without SA involvement [173], while others, such as Pseudomonas aeruginosa 7NSK2, may depend on SA induction, challenging the view that ISR is independent of SA [174]. Small RNAs are typically 20–30 nucleotide molecules, including microRNAs (miRNAs), small interfering RNAs (siRNAs), and piwi-interacting RNAs (piRNAs), which play important roles in regulating biological processes [175]. In plants, small RNAs can have both positive and negative effects on their resistance responses to pathogens [176]. Consequently, microorganisms can trigger ISR through two strategies. The first strategy involves promoting the expression of small RNAs that exert positive effects in plants. For instance, the expression of miR396d and miR408 is crucial for plant resistance responses [177,178]. When B. subtilis 26D triggers ISR in bread spring wheat against the aphid Rhopalosiphum padi, miR396d and miR408 are induced, and their expression levels increase sharply [179]. The second strategy is to inhibit the expression of small RNAs, which has negative effects on plants. For example, miR472 is a key regulator in Arabidopsis that negatively impacts plant disease-resistance genes. Bacillus cereus AR156 suppresses miR472 in Arabidopsis, thereby increasing the expression of the CC-NBS-LRR gene that miR472 targets, which subsequently triggers ISR in plants [180,181]. Similarly, B. velezensis FZB42 may trigger ISR by inhibiting the expression of zma-miR169a, zma-miR169c, and zma-miR169i. Understanding these multifaceted mechanisms enhances our ability to leverage beneficial microorganisms to improve plant resistance to PWN. Microbial induction of pine resistance to the PWN was first studied in Japan [182], applying an avirulent fungus, Botrytis cinerea, to PWN-infected black pine seedlings, demonstrating a decrease in average mortality from 97% to 89.9%, confirming that microorganisms can induce resistance in pine trees against PWN. One crucial defense response during pathogenic infections is the accumulation of pathogenesis-related proteins (PRs) [183]. Kim et al. (2019) discovered that genes associated with the PR-3 family (class I chitinase, class IV chitinase) and metallothionein-like proteins exhibited high transcript levels in calli treated with Pseudomonas putida 16YSM-E48, Curtobacterium pusillum 16YSM-P180, and Stenotrophomonas rhizophila 16YSM-M39 [184]. Additionally, Park et al. (2020) reported that Bacillus thuringiensis JCK-1233 could enhance PR expression in susceptible pine seedlings against PWN [166]. Han et al. (2021) illustrated the impact of microorganisms that induce resistance on the microbial diversity of pine trees. Treatment with Pseudomonas koreensis IRP7 and Lysobacter enzymogenes IRP8 led to increased relative abundance of beneficial microorganisms against PWN, including Nitrospirillum, Bacillus, Luteibacter, and obligate bacterial predators of the Bdellovibrio genus [185]. These findings underscore the potential of leveraging beneficial microorganisms to enhance resistance in pine trees against PWN.
6. The Application of Microbial BCAs
The field application of microbial control agents (BCAs) for managing the pine wood nematode (PWN) encounters several challenges, including reduced efficacy, elevated ecological safety risks, and high costs [186]. These obstacles are elaborated as follows: (1) Significant disparities exist between laboratory and field conditions for microbial BCAs. The broad latitudinal distribution of PWN demands that BCAs exhibit robust environmental adaptability [11]. Moreover, resistance in PWN and its vector insects, potentially mediated by multidrug resistance-associated protein (MRP) genes, can diminish BCA efficacy [17]. Application methods and conditions further influence performance. For example, a field test using B. bassiana ERL836 revealed varying mortality rates in M. alternatus depending on whether a spray suspension or fungal powder was used, as well as the concentration applied [106]. Consequently, comprehensive efficacy assessments are essential prior to BCA deployment to ensure optimal outcomes. (2) Environmental safety is a critical factor in microbial control strategies targeting PWN. Prolonged use of BCAs may affect local ecosystems, while some agents, such as B. bassiana ERL836, demonstrate minimal non-target effects [106]. Ecological risks persist due to diverse geographical conditions and forest types. Additionally, pine tree growth is closely tied to rhizosphere microorganisms. BCAs may help restore pine rhizosphere microbial communities [170]; however, a broader evaluation of their environmental impact is still required. Furthermore, certain BCAs are pathogenic, introducing biosafety concerns during production, storage, and transportation. (3) High costs represent a significant barrier to the widespread adoption of BCAs. Reducing production expenses, which often exceed those of traditional control agents, is imperative. For instance, solid-state fermentation (SSF) with rice as a carrier to produce Esteya vermicola conidia offers a scalable, cost-effective, and practical alternative to liquid fermentation [187]. Therefore, we propose that microbial consortia for field deployment meet the following standards: (1) high nematicidal activity; (2) low resistance risk; (3) strong environmental adaptability; (4) low ecological risk or pathogenicity after assessment; (5) low production cost in the field practice.
7. Conclusions
This review synthesizes the molecular mechanisms underpinning microbial control strategies against PWD caused by Bursaphelenchus xylophilus. This review makes three significant contributions to pine wilt disease management: (1) we systematically evaluate three prevention pathways through comparative mechanistic analysis, proposing synergistic integration strategies to enhance nematode control; (2) we identify key knowledge gaps in microbial control of PWN, emphasizing the need to investigate microorganism-induced systemic resistance, evaluate ecological impacts of microbial applications, and model climate-mediated pathogen-vector-microbe interactions; (3) we highlight bioinformatics and multi-omics integration as critical tools for elucidating biocontrol mechanisms while outlining field application challenges for microbial BCAs and establishing selection criteria for their practical deployment.
The mechanisms of the microbiological control of PWD are supported by molecular insights into hydrolase activity, toxin biosynthesis, and plant defense gene regulation, offering a foundation for targeted biocontrol applications (Figure 2). The integration of bioinformatic tools and multi-omic techniques has revolutionized the study of molecular mechanisms in PWD biocontrol. Genomic sequencing of nematophagous fungi (e.g., E. vermicola) and bacteria (e.g., B. thuringiensis) has identified key nematicidal genes, such as those encoding proteases and Cry toxins [187,188,189], while transcriptomic analyses have unveiled host-pathogen interaction dynamics, including pine defense gene activation during induced systemic resistance [190,191,192]. Metabolomics has enabled the profiling of nematicidal compounds, such as VOCs from P. violaceinigra, linking chemical diversity to biocontrol efficacy [72]. Proteomic studies further elucidate enzymatic pathways critical for fungal penetration of nematode cuticles or insect exoskeletons [193]. Additionally, metagenomics provides insights into microbial community shifts in PWN-infected pines, guiding the design of ecologically compatible consortia [160,194]. Despite these advances, challenges remain in data harmonization and functional validation of omics-derived hypotheses, necessitating interdisciplinary collaboration to translate molecular discoveries into field applications.
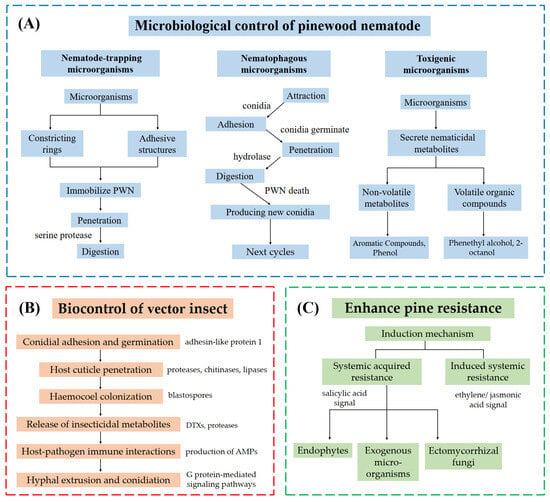
Figure 2.
Three strategies for biological control of pine wilt disease. (A) Direct microbiological control employs three mechanisms: nematode-trapping microorganisms immobilize pine wood nematode via constricting rings or adhesive structures, followed by tissue digestion through hydrolases (e.g., serine proteases); nematophagous fungi adhere to the nematode cuticle via conidia, which germinate, penetrate using hydrolases (proteases, chitinases), and digest internal tissues, with new conidia produced post-mortem to propagate infection; toxigenic microorganisms secrete nematicidal metabolites, including volatile organic compounds (VOCs) and non-volatile toxins. (B) Biological control of insect vectors involves six stages: adhesin-like protein 1 mediates conidial adhesion and germination on the host cuticle; hydrolases (proteases, chitinases, lipases) degrade the cuticle for fungal penetration; blastospores colonize the hemocoel; insecticidal metabolites (e.g., destruxins [DTXs], proteases) suppress host immunity; antimicrobial peptides (AMPs) modulate host-pathogen immune interactions; G protein-mediated signaling induces hyphal extrusion and conidiation to initiate new infection cycles. (C) Pine resistance is enhanced via systemic defense priming: systemic acquired resistance (SAR, salicylic acid-mediated) and induced systemic resistance (ISR, ethylene/jasmonic acid-mediated).
An integrated approach combining microbial agents with traditional management practices holds significant potential for sustainable PWD control. For instance, pairing nematophagous fungi with pheromone-based vector trapping or leveraging VOCs for dual nematode suppression and pine priming could enhance efficacy. The synergistic interactions among different biocontrol microorganisms require further study. The toxigenic bacterium Stenotrophomonas maltophilia, which produces nematocidal VOCs, can induce trapping structure formation in the nematode-trapping fungus A. oligospora [195]. A recent study showed S. maltophilia also directly triggers trapping structures by endogenizing within and participating in the nitrogen cycle of Arthrobotrys musiformis [196]. Genetic engineering of microbial strains to overexpress nematicidal proteins (e.g., Cry6Aa in B. toyonensis) or optimize VOC production may further improve field performance. However, ecological compatibility must be prioritized to avoid disrupting native microbial communities. Regional PWN distribution patterns, as highlighted in Figure 1, necessitate tailored strategies, such as prioritizing vector control in newly invaded areas and resistance enhancement in endemic zones.
Critical research gaps persist in understanding molecular mechanisms. First, the precise pathways by which microbes induce SAR/ISR in pines—particularly the roles of phytohormones, small RNAs, and secondary metabolites—remain poorly characterized. Second, the ecological interplay between introduced biocontrol agents, native microbiota, and PWN in forest ecosystems is underexplored, raising concerns about long-term sustainability. Third, while lab studies demonstrate the high efficacy of microbial agents (e.g., Pseudoduganella violaceinigra VOCs achieving >90% nematode mortality), field applications often face challenges due to environmental variability and host-pathogen co-evolution. Fourth, it is crucial to study the molecular mechanisms by which different types of biocontrol microorganisms (e.g., nematode-trapping fungi and VOC-producing bacteria) exert synergistic effects. Additionally, the molecular basis of PWN resistance to microbial toxins and the impact of climate change on microbial-nematode-vector tripartite interactions require deeper investigation. Addressing these gaps will be pivotal for advancing biocontrol strategies that are both effective and ecologically resilient.
In conclusion, the integration of molecular insights, ecological considerations, and innovative technologies will drive the development of next-generation biocontrol solutions. Future research should prioritize elucidating unresolved molecular interactions, optimizing microbial consortia for field stability, and validating integrated strategies under diverse environmental conditions to safeguard global pine forests.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms13061215/s1, Table S1: Diversity of biocontrol microorganisms against PWN. References [197,198,199,200,201,202,203,204,205,206,207,208,209,210,211,212,213] are cited in the supplementary materials.
Author Contributions
Coordinated the project, H.W. and Y.L. (Yongxia Li); wrote the manuscript, X.S., Y.L. (Yimou Luo) and Y.X.; revised the manuscript, J.H. and M.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Natural Science Foundation of China, 32400097 and Natural Science Foundation of Shandong Province, ZR2021MC149.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kusunoki, M. Symptom development of pine wilt disease-histopathological observations with electron microscopes. Jpn. J. Phytopathol. 1987, 53, 622–629. [Google Scholar] [CrossRef][Green Version]
- Mamiya, Y. Pathology of the pine wilt disease caused by Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 1983, 21, 201–220. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Guo, Q.; Xie, N.; Yuan, G.; Liao, M.; Gui, Q.; Ding, G. Predicting the global potential distribution of Bursaphelenchus xylophilus using an ecological niche model: Expansion trend and the main driving factors. BMC Ecol. Evol. 2024, 24, 48. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.C. Pine Wilt Disease in Korea. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 26–32. [Google Scholar]
- Hao, Z.; Huang, J.; Li, X.; Sun, H.; Fang, G. A multi-point aggregation trend of the outbreak of pine wilt disease in China over the past 20 years. For. Ecol. Manag. 2022, 505, 119890. [Google Scholar] [CrossRef]
- Vicente, C.; Espada, M.; Vieira, P.; Mota, M. Pine wilt disease: A threat to European forestry. Eur. J. Plant Pathol. 2012, 133, 89–99. [Google Scholar] [CrossRef]
- Back, M.A.; Bonifácio, L.; Inácio, M.L.; Mota, M.; Boa, E. Pine wilt disease: A global threat to forestry. Plant Pathol. 2024, 73, 1026–1041. [Google Scholar] [CrossRef]
- Mamiya, Y. History of pine wilt disease in Japan. J. Nematol. 1988, 20, 219. [Google Scholar]
- Yang, B.J. The history, dispersal and potential threat of pine wood nematode in China. In The Pinewood Nematode, Bursaphelenchus xylophilus; Brill: Leiden, The Netherlands, 2004; pp. 21–24. [Google Scholar]
- Zhang, X.; Zhao, J.; Yan, J.; Fang, G.; Huang, J. Economic loss assessment of pine wilt disease in mainland China in 2017. J. Beijing For. Univ. 2020, 42, 96–106. [Google Scholar] [CrossRef]
- Xu, Q.; Zhang, X.; Li, J.; Ren, J.; Ren, L.; Luo, Y. Pine wilt disease in Northeast and Northwest China: A comprehensive risk review. Forests 2023, 14, 174. [Google Scholar] [CrossRef]
- Zhao, H.; Xian, X.; Yang, N.; Guo, J.; Zhao, L.; Shi, J.; Liu, W. Risk assessment framework for pine wilt disease: Estimating the introduction pathways and multispecies interactions among the pine wood nematode, its insect vectors, and hosts in China. Sci. Total Environ. 2023, 905, 167075. [Google Scholar] [CrossRef]
- Ryss, A.Y.; Kulinich, O.A.; Sutherland, J.R. Pine wilt disease: A short review of worldwide research. For. Stud. China. 2011, 13, 132–138. [Google Scholar] [CrossRef]
- Bi, Z.; Gong, Y.; Huang, X.; Yu, H.; Bai, L.; Hu, J. Efficacy of four nematicides against the reproduction and development of pinewood nematode, Bursaphelenchus xylophilus. J. Nematol. 2015, 47, 126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Si, G.; Chen, L.; Hu, L.; Cui, G.; Wang, M.; Zhao, D. Current Status and Prospects of Pine Wilt Disease Management with Phytochemicals—A Review. Plants 2024, 13, 2129. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Hu, J.; Pang, H.; Yin, S.; Liu, H.; Tian, Y.; Gao, S.; Zhou, C.; Wu, P.; Miao, S.; et al. Sustainable DMSNs nano-biopesticide platform built by a “one-pot” method focusing on injury-free drug demonstration of pine wood nematodes. Environ. Sci. Nano 2024, 11, 363–372. [Google Scholar] [CrossRef]
- Hao, X.; Wang, B.; Chen, J.; Wang, B.; Xu, J.; Pan, J.; Ma, L. Molecular characterization and functional analysis of multidrug resistance-associated genes of Pinewood nematode (Bursaphelenchus xylophilus) for nematicides. Pestic. Biochem. Physiol. 2021, 177, 104902. [Google Scholar] [CrossRef]
- Pires, D.; Vicente, C.S.L.; Menendez, E.; Faria, J.M.S.; Rusinque, L.; Camacho, M.J.; Inacio, M.L. The Fight against Plant-Parasitic Nematodes: Current Status of Bacterial and Fungal Biocontrol Agents. Pathogens 2022, 11, 1178. [Google Scholar] [CrossRef]
- Ning, T.; Fang, Y.; Tang, J.; Sun, J. Advances in research on Bursaphelenchus xyophilus and its key vector Monochamus spp. Chin. J. Appl. Entomol. 2004, 41, 97–104. [Google Scholar] [CrossRef]
- Ye, J. Epidemic status of pine wilt disease in China and its prevention and control techniques and counter measures. Sci. Silvae Sin. 2019, 55, 1–10. [Google Scholar] [CrossRef]
- Jeon, H.W.; Park, A.R.; Sung, M.; Kim, N.; Mannaa, M.; Han, G.; Kim, J.; Koo, Y.; Seo, Y.S.; Kim, J.C. Systemic Acquired Resistance-Mediated Control of Pine Wilt Disease by Foliar Application with Methyl Salicylate. Front. Plant Sci. 2021, 12, 812414. [Google Scholar] [CrossRef]
- Xu, M.; Xu, F.Y.; Liu, Y.P.; Pan, Y.S.; Wu, X.Q. Assessment of Metarhizium anisopliae (Clavicipitaceae) and its vector, Scleroderma guani (Hymenoptera: Bethylidae), for the control of Monochamus alternatus (Coleoptera: Cerambycidae). Can. Entomol. 2015, 147, 628–634. [Google Scholar] [CrossRef]
- Xu, F.; Xu, K.; Xie, C.; Zhang, P.; Shin, S.; Cheong, Y. Studies on Scleroderma guani to Control the Pine Sawyer Beetle, Monochamus alternatus. In Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Mota, M.M., Vieira, P., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 379–388. [Google Scholar]
- Dou, G.; Yan, D. Research Progress on Biocontrol of Pine Wilt Disease by Microorganisms. Forests 2022, 13, 1047. [Google Scholar] [CrossRef]
- Hayashi, A.; Fujioka, S.; Nukina, M.; Kawano, T.; Shimada, A.; Kimura, Y. Fumiquinones A and B, nematicidal quinones produced by Aspergillus fumigatus. Biosci. Biotechnol. Biochem. 2007, 71, 1697–1702. [Google Scholar] [CrossRef] [PubMed]
- Akbulut, S.; Stamps, W. Insect vectors of the pinewood nematode: A review of the biology and ecology of Monochamus species. For. Pathol. 2012, 42, 89–99. [Google Scholar] [CrossRef]
- Zhao, L.; Mota, M.; Vieira, P.; Butcher, R.A.; Sun, J. Interspecific communication between pinewood nematode, its insect vector, and associated microbes. Trends Parasitol. 2014, 30, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Jung, J.K.; Hong, K.J.; Kim, C.J.; Lee, B.W.; Kim, I.K. Discovery and biology of Spathius verustus Chao (Hymenoptera: Braconidae), a potential biological agent on two Monochamus vectors of the pinewood nematode. Forests 2022, 13, 955. [Google Scholar] [CrossRef]
- Liu, X.; Song, J.; Li, X.; Xu, H.; Wang, Z. Current Status of Research on Control Methods of Monochamus Alternatus Hope (Co-leoptera: Cerambycidae). Inst. Contin. Judic. Educ. 2022, 8, 21–29. [Google Scholar] [CrossRef]
- Yang, J.; Li, J.; Liang, L.; Tian, B.; Zhang, Y.; Cheng, C.; Zhang, K.Q. Cloning and characterization of an extracellular serine protease from the nematode-trapping fungus Arthrobotrys conoides. Arch. Microbiol. 2007, 188, 167–174. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, C.Y.; Gu, L.J.; Sun, B.S.; Zhang, D.L.; Liu, L.; Lee, M.R.; Wang, C.L.; Li, Z.; Mo, E.K.; et al. Variabilities of two Drechslerella dactyloides isolates in Korea and high predacity against Bursaphelenchus xylophilus. Curr. Microbiol. 2011, 62, 472–478. [Google Scholar] [CrossRef]
- Jinyan, D.; Ru, L.; Keqin, Z. Advances in biological control of Bursaphelenchus xylophilus by natural enemies and nematocidal compounds from plants. Plant Prot. 2005, 31, 9–15. [Google Scholar]
- Zhang, X.; Zhang, H.; Jiang, Z.; Bai, Q.; Wu, S.; Wang, Y.; Li, C.; Zeng, X.; Gan, X.; Xie, X. A new strain of Volutella citrinella with nematode predation and nematicidal activity, isolated from the cysts of potato cyst nematodes in China. BMC Microbiol. 2021, 21, 323. [Google Scholar] [CrossRef]
- Zhu, M.; Li, X.; Zhao, N.; Yang, L.; Zhang, K.; Yang, J. Regulatory mechanism of trap formation in the nematode-trapping fungi. J. Fungi 2022, 8, 406. [Google Scholar] [CrossRef] [PubMed]
- Tunlid, A.; Jansson, H.; Nordbring-Hertz, B. Fungal attachment to nematodes. Mycol. Res. 1992, 96, 401–412. [Google Scholar] [CrossRef]
- Wang, D.; Ma, N.; Rao, W.; Zhang, Y. Recent advances in life history transition with nematode-trapping fungus Arthrobotrys oligospora and its application in sustainable agriculture. Pathogens 2023, 12, 367. [Google Scholar] [CrossRef] [PubMed]
- Bai, N.; Zhang, G.; Wang, W.; Feng, H.; Yang, X.; Zheng, Y.; Yang, L.; Xie, M.; Zhang, K.-Q.; Yang, J. Ric8 acts as a regulator of G-protein signalling required for nematode-trapping lifecycle of Arthrobotrys oligospora. Environ. Microbiol. 2022, 24, 1714–1730. [Google Scholar] [CrossRef]
- Tokumitsu, H.; Sakagami, H. Molecular mechanisms underlying Ca2+/calmodulin-dependent protein kinase kinase signal transduction. Int. J. Mol. Sci. 2022, 23, 11025. [Google Scholar] [CrossRef]
- Zhen, Z.; Zhang, G.; Yang, L.; Ma, N.; Li, Q.; Ma, Y.; Niu, X.; Zhang, K.; Yang, J. Characterization and functional analysis of calcium/calmodulin-dependent protein kinases (CaMKs) in the nematode-trapping fungus Arthrobotrys oligospora. Appl. Microbiol. Biotechnol. 2019, 103, 819–832. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, N.; Zhang, K. Extracellular enzymes serving as virulence factors in nematophagous fungi involved in infection of the host. Res. Microbiol. 2004, 155, 811–816. [Google Scholar] [CrossRef]
- Yang, J.; Tian, B.; Liang, L.; Zhang, K. Extracellular enzymes and the pathogenesis of nematophagous fungi. Appl. Microbiol. Biotechnol. 2007, 75, 21–31. [Google Scholar] [CrossRef]
- Vidal-Diez de Ulzurrun, G.; Hsueh, Y. Predator-prey interactions of nematode-trapping fungi and nematodes: Both sides of the coin. Appl. Microbiol. Biotechnol. 2018, 102, 3939–3949. [Google Scholar] [CrossRef]
- Veenhuis, M.; Van Wijk, C.; Wyss, U.; Nordbring-Hertz, B.; Harder, W. Significance of electron dense microbodies in trap cells of the nematophagous fungus Arthrobotrys oligospora. Antonie Van Leeuwenhoek 1989, 56, 251–261. [Google Scholar] [CrossRef]
- Paiva, G.; Proenca, D.N.; Francisco, R.; Verissimo, P.; Santos, S.S.; Fonseca, L.; Abrantes, I.M.; Morais, P.V. Nematicidal bacteria associated to pinewood nematode produce extracellular proteases. PLoS ONE 2013, 8, e79705. [Google Scholar] [CrossRef] [PubMed]
- Vicente, C.S.L.; Soares, M.; Faria, J.M.S.; Ramos, A.P.; Inacio, M.L. Insights into the Role of Fungi in Pine Wilt Disease. J. Fungi 2021, 7, 780. [Google Scholar] [CrossRef] [PubMed]
- Liou, J.; Shih, J.; Tzean, S. Esteya, a new nematophagous genus from Taiwan, attacking the pinewood nematode (Bursaphelenchus xylophilus). Mycol. Res. 1999, 103, 242–248. [Google Scholar] [CrossRef]
- Lin, F.; Ye, J.; Wang, H.; Zhang, A.; Zhao, B. Host deception: Predaceous fungus, Esteya vermicola, entices pine wood nematode by mimicking the scent of pine tree for nutrient. PLoS ONE 2013, 8, e71676. [Google Scholar] [CrossRef]
- Wang, C.Y.; Wang, Z.; Fang, Z.M.; Zhang, D.L.; Gu, L.J.; Liu, L.; Sung, C.K. Attraction of pinewood nematode to endoparasitic nematophagous fungus Esteya vermicola. Curr. Microbiol. 2010, 60, 387–392. [Google Scholar] [CrossRef]
- Wang, H.H.; Wang, Y.B.; Yin, C.; Gao, J.; Tao, R.; Sun, Y.L.; Wang, C.Y.; Wang, Z.; Li, Y.X.; Sung, C.K. In vivo infection of Bursaphelenchus xylophilus by the fungus Esteya vermicola. Pest Manag. Sci. 2020, 76, 2854–2864. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, C.Y.; Yang, Z.H.; Fang, Z.M.; Moon, Y.J.; Sun, B.S.; Lee, M.R.; Sung, C.K. Viability and pathogenicity of Esteya vermicola in pine trees. Biocontrol Sci. Technol. 2011, 21, 387–393. [Google Scholar] [CrossRef]
- Yin, C.; Wang, Y.; Zhang, Y.; Wang, H.; Duan, B.; Tao, R.; Gao, J.; Sung, C. Hypothesized mechanism of biocontrol against pine wilt disease by the nematophagous fungus Esteya vermicola. Eur. J. Plant Pathol. 2020, 156, 811–818. [Google Scholar] [CrossRef]
- Li, Y.; Yu, H.; Araújo, J.P.M.; Zhang, X.; Ji, Y.; Hulcr, J. Esteya floridanum sp. nov.: An Ophiostomatalean nematophagous fungus and its potential to control the pine wood nematode. Phytopathology 2021, 111, 304–311. [Google Scholar] [CrossRef]
- Guo, Y.; Weng, M.; Sun, Y.; Carballar-Lejarazú, R.; Wu, S.; Lian, C. Bacillus thuringiensis toxins with nematocidal activity against the pinewood nematode Bursaphelenchus xylophilus. J. Invertebr. Pathol. 2022, 189, 107726. [Google Scholar] [CrossRef]
- Li, L.; Tan, J.; Chen, F. Bacillus pumilus strain LYMC-3 shows nematicidal activity against Bursaphelenchus xylophilus via the production of a guanidine compound. Biocontrol Sci. Technol. 2018, 28, 1128–1139. [Google Scholar] [CrossRef]
- Wang, Y.; Mei, L.; Wu, J.; Zhang, L. Detection and characterisation of a Bacillus thuringiensis crystal protein with nematicidal activity against the pinewood nematode Bursaphelenchus xylophilus. Biocontrol Sci. Technol. 2012, 22, 1143–1153. [Google Scholar] [CrossRef]
- Li, L.; Sun, Y.; Chen, F.; Hao, D.; Tan, J. An alkaline protease from Bacillus cereus NJSZ-13 can act as a pathogenicity factor in infection of pinewood nematode. BMC Microbiol. 2023, 23, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, Y.; Wang, X.; Zhang, W.; Wen, X.; Liu, Z.; Feng, Y.; Zhang, X. Engineered pine endophytic Bacillus toyonensis with nematocidal and colonization abilities for pine wilt disease control. Front. Microbiol. 2023, 14, 1240984. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Chen, S.; Lu, F.; Guo, K.; Huang, L.; Su, X.; Chen, Y. Nematotoxicity of a Cyt-like protein toxin from Conidiobolus obscurus (Entomophthoromycotina) on the pine wood nematode Bursaphelenchus xylophilus. Pest Manag. Sci. 2021, 77, 686–692. [Google Scholar] [CrossRef]
- Kang, M.K.; Kim, M.H.; Liu, M.J.; Jin, C.Z.; Park, S.H.; Lee, J.M.; Kim, J.; Park, D.; Park, H.R.; Kim, Y.H.; et al. Nematicidal activity of teleocidin B4 isolated from Streptomyces sp. against pine wood nematode, Bursaphelenchus xylophilus. Pest Manag. Sci. 2021, 77, 1607–1615. [Google Scholar] [CrossRef]
- Kang, M.K.; Kim, J.H.; Liu, M.J.; Jin, C.Z.; Park, D.J.; Kim, J.; Son, K.H.; Kim, C.J. New discovery on the nematode activity of aureothin and alloaureothin isolated from endophytic bacteria Streptomyces sp. AE170020. Sci Rep. 2022, 12, 3947. [Google Scholar] [CrossRef]
- Zheng, L.; Li, G.; Wang, X.; Pan, W.; Li, L.; Hua, L.; Liu, F.; Dang, L.; Mo, M.; Zhang, K. Nematicidal endophytic bacteria obtained from plants. Ann. Microbiol. 2008, 58, 569–572. [Google Scholar] [CrossRef]
- Zeng, L.; Jin, H.; Lu, D.; Yang, X.; Pan, L.; Cui, H.; He, X.; Qiu, H.; Qin, B. Isolation and identification of chemical constituents from the bacterium Bacillus sp. and their nematicidal activities. J. Basic Microbiol. 2015, 55, 1239–1244. [Google Scholar] [CrossRef]
- Liu, M.J.; Hwang, B.S.; Jin, C.Z.; Li, W.J.; Park, D.J.; Seo, S.T.; Kim, C.J. Screening, isolation and evaluation of a nematicidal compound from actinomycetes against the pine wood nematode, Bursaphelenchus xylophilus. Pest Manag. Sci. 2019, 75, 1585–1593. [Google Scholar] [CrossRef]
- Sun, S.; Li, W.; Ju, K.; Xiong, X.; Li, J.; Yu, C.; Tian, Y.; Liu, H. Tetramycin B3: An Effective and Biological Nematicide for Bursaphelenchus xylophilus. Forests 2024, 15, 1699. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, C.; Du, G.; Deng, W.; Yang, H.; Li, R.; Xu, Q.; Guo, Q. Two nematicidal compounds from Lysinimonas M4 against the pine wood nematode, Bursaphelenchus xylophilus. Forests 2022, 13, 1191. [Google Scholar] [CrossRef]
- Huang, X.; Liu, J.; Ding, J.; He, Q.; Xiong, R.; Zhang, K. The investigation of nematocidal activity in Stenotrophomonas maltophilia G2 and characterization of a novel virulence serine protease. Can. J. Microbiol. 2009, 55, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.D.; Qi, H.; Zhang, J.; Li, J.S.; Zhang, S.Y.; Hao, Z.K.; Zhang, L.Q.; Xiang, W.S. Two new 13-hydroxylated milbemycin metabolites from the genetically engineered strain Streptomyces avermitilis AVE-H39. J. Asian Nat. Prod. Res. 2021, 23, 837–843. [Google Scholar] [CrossRef]
- Zhu, L.; Wu, X.; Hu, F.; Xu, H. Extraction and structure analysis of the nematicidal active substances produced by Bacillus amyloliquefaciens JK-JS3. Sci. Silvae Sin. 2011, 47, 133–137. [Google Scholar]
- Huang, B.; Chen, J.; Li, W.; Song, X.; Wang, B.; Zhang, C.; Zhao, S.; Niu, Q. Screening and identification of actinomycetes for biological control of Bursaphelenchus xylophilus and preliminary study on their toxicity factors. J. Beijing For. Univ. 2019, 41, 99–106. [Google Scholar] [CrossRef]
- Proença, D.N.; Heine, T.; Senges, C.H.; Bandow, J.E.; Morais, P.V.; Tischler, D. Bacterial metabolites produced under iron limitation kill pinewood nematode and attract Caenorhabditis elegans. Front. Microbiol. 2019, 10, 2166. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, C.; Zhang, S.; Chen, F.; Pei, F.; Zhou, S.; Lin, H. Screening, isolation and mechanism of a nematicidal extract from actinomycetes against the pine wood nematode Bursaphelenchus xylophilus. Heliyon 2022, 8, e11713. [Google Scholar] [CrossRef]
- Wang, F.; Guo, Q.; Wang, L.; Ma, Y.; Zhang, T.; Li, R. Nematicidal activities of bacterial volatiles from Pseudoduganella violaceinigra G5-3 and Novosphingobium pokkalii G8-2 against the pine wood nematode Bursaphelenchus xylophilus. Chiang Mai J. Sci. 2019, 46, 236–246. [Google Scholar]
- Gu, Y.Q.; Mo, M.H.; Zhou, J.P.; Zou, C.S.; Zhang, K.Q. Evaluation and identification of potential organic nematicidal volatiles from soil bacteria. Soil Biol. Biochem. 2007, 39, 2567–2575. [Google Scholar] [CrossRef]
- Yu, J.; Du, G.; Li, R.; Li, L.; Li, Z.; Zhou, C.; Chen, C.; Guo, D. Nematicidal activities of bacterial volatiles and components from two marine bacteria, Pseudoalteromonas marina strain H-42 and Vibrio atlanticus strain S-16, against the pine wood nematode, Bursaphelenchus xylophilus. Nematology 2015, 17, 1011–1025. [Google Scholar] [CrossRef]
- Li, G.H.; Yu, Z.F.; Li, X.; Wang, X.B.; Zheng, L.J.; Zhang, K.Q. Nematicidal metabolites produced by the endophytic fungus Geotrichum sp. AL4. Chem. Biodivers. 2007, 4, 1520–1524. [Google Scholar] [CrossRef] [PubMed]
- Lou, J.; Yu, R.; Wang, X.; Mao, Z.; Fu, L.; Liu, Y.; Zhou, L. Alternariol 9-methyl ether from the endophytic fungus Alternaria sp. Samif01 and its bioactivities. Braz. J. Microbiol. 2016, 47, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Dong, J.; Wang, L.; Zhou, W.; Li, L.; He, H.; Liu, H.; Zhang, K. Screening and isolation of antinematodal metabolites against Bursaphelenchus xylophilus produced by fungi. Ann. Microbiol. 2008, 58, 375–380. [Google Scholar] [CrossRef]
- Ohtani, K.; Fujioka, S.; Shimada, A.; Kimura, Y. Nematicidal activities of 4-hydroxyphenylacetic acid and oidiolactone D produced by the fungus Oidiodendron sp. Z. Naturforsch. C 2011, 66, 31–34. [Google Scholar] [CrossRef]
- Song, H.; Shen, W.; Dong, J. Nematicidal metabolites from Gliocladium roseum YMF1. 00133. Appl. Biochem. Microbiol. 2016, 52, 324–330. [Google Scholar] [CrossRef]
- Dong, J.; Li, G.; Zhang, K. Screening and isolation of anti-nematodal metabolites against Bursaphelenchus xylophilus produced by fungi and plant. In Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Mota, M.M., Vieira, P., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 347–358. [Google Scholar]
- Dong, J.Y.; Li, R.; He, H.P.; Zhang, K.Q. Nematicidal sphingolipids from the freshwater fungus Paraniesslia sp. YMF1. 01400. Eur. J. Lipid Sci. Technol. 2005, 107, 779–785. [Google Scholar] [CrossRef]
- Dong, J.Y.; Song, H.C.; Li, J.H.; Tang, Y.S.; Sun, R.; Wang, L.; Zhou, Y.P.; Wang, L.M.; Shen, K.Z.; Wang, C.R.; et al. Ymf 1029A−E, Preussomerin Analogues from the Fresh-Water-Derived Fungus YMF 1.01029. J. Nat. Prod. 2008, 71, 952–956. [Google Scholar] [CrossRef]
- Zhou, Y.; Shen, K.; Dong, J.; Wang, L.; Sun, R.; Wang, C.; Wang, L.; Zhang, K. Nematicidal metabolites of the aquatic fungus Coelomycetes sp. YMF1. 01029. Chin. J. Antibiot. 2009, 34, 74–78. [Google Scholar]
- Kamaruzzaman, M.; Zheng, L.; Zhou, S.; Ye, W.; Yuan, Y.; Qi, Q.; Gao, Y.; Tan, J.; Wang, Y.; Chen, B. Evaluation of the novel endophytic fungus Chaetomium ascotrichoides 1-24-2 from Pinus massoniana as a biocontrol agent against pine wilt disease caused by Bursaphelenchus xylophilus. Pest Manag. Sci. 2024, 80, 4924–4940. [Google Scholar] [CrossRef]
- Kwon, H.R.; Son, S.W.; Han, H.R.; Choi, G.J.; Jang, K.S.; Choi, Y.H.; Lee, S.; Sung, N.D.; Kim, J.C. Nematicidal activity of bikaverin and fusaric acid isolated from Fusarium oxysporum against pine wood nematode, Bursaphelenchus xylophilus. Plant Pathol. J. 2007, 23, 318–321. [Google Scholar] [CrossRef]
- Kimura, Y.; Tani, S.; Hayashi, A.; Ohtani, K.; Fujioka, S.; Kawano, T.; Shimada, A. Nematicidal activity of 5-hydroxymethyl-2-furoic acid against plant-parasitic nematodes. Z. Naturforsch. C 2007, 62, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Gómez, T.; Harte, S.J.; Zamora, P.; Bareyre, M.; Díez, J.J.; Herrero, B.; Niño-Sánchez, J.; Martín-García, J. Nematicidal effect of Beauveria species and the mycotoxin beauvericin against pinewood nematode Bursaphelenchus xylophilus. Front. For. Glob. Chang. 2023, 6, 1229456. [Google Scholar] [CrossRef]
- Dong, J.Y.; Wang, L.; Song, H.C.; Wang, L.M.; Shen, K.Z.; Sun, R.; Li, G.H.; Li, L.; Zhang, K.Q. Ophiocerol, a novel macrocylic neolignan from the aquatic fungus Ophioceras dolichostomum YMF1.00988. Nat. Prod. Res. 2010, 24, 1004–1012. [Google Scholar] [CrossRef]
- Dong, J.Y.; He, H.P.; Shen, Y.M.; Zhang, K.Q. Nematicidal Epipolysulfanyldioxopiperazines from Gliocladium roseum. J. Nat. Prod. 2005, 68, 1510–1513. [Google Scholar] [CrossRef]
- Yang, Z.; Yu, Z.; Lei, L.; Xia, Z.; Shao, L.; Zhang, K.; Li, G. Nematicidal effect of volatiles produced by Trichoderma sp. J. Asia-Pac. Entomol. 2012, 15, 647–650. [Google Scholar] [CrossRef]
- Li, H.; Dou, G.; Gao, M.; Ren, F.; Li, R.; Zhang, X.; Yan, D.H. Annulohypoxylon sp. FPYF3050 produces volatile organic compounds against the pine wood nematode, Bursaphelenchus xylophilus. Nematology 2020, 22, 245–255. [Google Scholar] [CrossRef]
- Dong, J.; Zhao, Z.; Cai, L.; Liu, S.; Zhang, H.; Duan, M.; Zhang, K. Nematicidal effect of freshwater fungal cultures against the pine-wood nematode, Bursaphelenchus xylophilus. Fungal Divers. 2004, 15, 125–135. [Google Scholar]
- Shimada, A.; Fujioka, S.; Koshino, H.; Kimura, Y. Nematicidal activity of beauvericin produced by the fungus Fusarium bulbicola. Z. Naturforsch. C 2010, 65, 207–210. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, X.; Huang, C.; Zhou, J.; Shi, J.; Zhao, L. Temperature-regulated metabolites of Serratia marcescens inhibited reproduction of pinewood nematode Bursaphelenchus xylophilus. iScience 2023, 26, 107082. [Google Scholar] [CrossRef]
- Wang, S.; Han, M.; Li, H.; Xie, J.; Wei, K.; Wang, X. Testing the host range of Cyanopterus ninghais (Hymenoptera: Braconidae), a candidate for the biological control of Monochamus alternatus, the vector of pine wilt disease in Asia. Biol. Control. 2024, 195, 105547. [Google Scholar] [CrossRef]
- Kim, J.C.; Lee, S.J.; Kim, S.; Lee, M.R.; Baek, S.; Park, S.E.; Kim, J.; Shin, T.Y.; Kim, J.S. Management of pine wilt disease vectoring Monochamus alternatus adults using spray and soil application of Metarhizium anisopliae JEF isolates. J. Asia-Pac. Entomol. 2020, 23, 224–233. [Google Scholar] [CrossRef]
- Álvarez-Baz, G.; Fernández-Bravo, M.; Pajares, J.; Quesada-Moraga, E. Potential of native Beauveria pseudobassiana strain for biological control of Pine Wood Nematode vector Monochamus galloprovincialis. J. Invertebr. Pathol. 2015, 132, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.Y.; Shi, H.L.; Wang, D. A New Strain of Metarhizium robertsii Isolated from Loess Plateau and Its Virulence and Pathological Characteristics against Monochamus alternatus. Microorganisms 2024, 12, 514. [Google Scholar] [CrossRef]
- Tian, H.; Koski, T.M.; Zhao, L.; Liu, Z.; Sun, J. Invasion history of the pinewood nematode Bursaphelenchus xylophilus influences the abundance of Serratia sp. in pupal chambers and tracheae of insect-vector Monochamus alternatus. Front. Plant Sci. 2022, 13, 856841. [Google Scholar] [CrossRef]
- Wu, S.; Wu, J.; Wang, Y.; Qu, Y.; He, Y.; Wang, J.; Cheng, J.; Zhang, L.; Cheng, C. Discovery of entomopathogenic fungi across geographical regions in southern China on pine sawyer beetle Monochamus alternatus and implication for multi-pathogen vectoring potential of this beetle. Front. Plant Sci. 2022, 13, 1061520. [Google Scholar] [CrossRef]
- Hussein, K.A.; Abdel-Rahman, M.A.; Abdel-Mallek, A.Y.; El-Maraghy, S.S.; Joo, J.H. Climatic factors interference with the occurrence of Beauveria bassiana and Metarhizium anisopliae in cultivated soil. Afr. J. Biotechnol. 2010, 9, 7674–7682. [Google Scholar]
- Fernandes, E.K.; Rangel, D.E.; Moraes, A.M.; Bittencourt, V.R.; Roberts, D.W. Cold activity of Beauveria and Metarhizium, and thermotolerance of Beauveria. J. Invertebr. Pathol. 2008, 98, 69–78. [Google Scholar] [CrossRef]
- Omuse, E.R.; Niassy, S.; Wagacha, J.M.; Ong’amo, G.O.; Azrag, A.G.; Dubois, T. Suitable models to describe the effect of temperature on conidial germination and mycelial growth of Metarhizium anisopliae and Beauveria bassiana. Biocontrol Sci. Technol. 2022, 32, 281–298. [Google Scholar] [CrossRef]
- McGuire, A.V.; Northfield, T.D. Tropical occurrence and agricultural importance of Beauveria bassiana and Metarhizium anisopliae. Front. Sustain. Food Syst. 2020, 4, 6. [Google Scholar] [CrossRef]
- Kim, J.C.; Lee, M.R.; Kim, S.; Lee, S.J.; Park, S.E.; Baek, S.; Gasmi, L.; Shin, T.Y.; Kim, J.S. Long-term storage stability of Beauveria bassiana ERL836 granules as fungal biopesticide. J. Asia-Pac. Entomol. 2019, 22, 537–542. [Google Scholar] [CrossRef]
- Kim, J.C.; Lee, M.R.; Yu, J.S.; Park, S.E.; Ha, P.; Kim, J.S. Management of overwintering pine sawyer beetle, Monochamus alternatus with colonized Beauveria bassiana ERL836. PLoS ONE 2022, 17, e0274086. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhang, L.; Lin, H.; Mao, S. Investigation of pathogens of Monochamus alternatus in East China and virulence. Chin. J. Biol. Control. 2009, 25, 220–225. [Google Scholar] [CrossRef]
- Shimazu, M. Potential of the cerambycid-parasitic type of Beauveria brongniartii (Deuteromycotina: Hyphomycetes) for microbial control of Monochamus alternatus Hope (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1994, 29, 127–130. [Google Scholar] [CrossRef][Green Version]
- Maehara, N.; He, X.; Shimazu, M. Maturation feeding and transmission of Bursaphelenchus xylophilus (Nematoda: Parasitaphelenchidae) by Monochamus alternatus (Coleoptera: Cerambycidae) inoculated with Beauveria bassiana (Deuteromycotina: Hyphomycetes). J. Econ. Entomol. 2007, 100, 49–53. [Google Scholar] [CrossRef]
- Takatsuka, J. Specific PCR assays for the detection of DNA from Beauveria bassiana F-263, a highly virulent strain affecting the Japanese pine sawyer, Monochamus alternatus (Coleoptera: Cerambycidae), by a sequence-characterized amplified region (SCAR) marker. Appl. Entomol. Zool. 2007, 42, 619–628. [Google Scholar] [CrossRef][Green Version]
- He, R.; Cui, X.; Ying, Y.; Qu, L.; Wang, R.; Zhang, Y. Screening and identification of Beauveria bassiana strains for biocontrol of Monochamus alternatus adults (Coleoptera: Cerambycidae). Sci. Silvae Sin. 2020, 56, 129–134. [Google Scholar] [CrossRef]
- Kim, H.M.; Choi, I.S.; Lee, S.; Hwang, I.M.; Chun, H.H.; Wi, S.G.; Kim, J.C.; Shin, T.Y.; Kim, J.C.; Kim, J.S. Advanced strategy to produce insecticidal destruxins from lignocellulosic biomass Miscanthus. Biotechnol. Biofuels 2019, 12, 188. [Google Scholar] [CrossRef]
- He, X.; Chen, S.; Huang, J. Preliminary screening of virulent strains of Metarhizium anisopliae against Monochamus alternatus. Acta Entomol. Sin. 2005, 6, 975–981. [Google Scholar] [CrossRef]
- Kim, J.C.; Baek, S.; Park, S.E.; Kim, S.; Lee, M.R.; Jo, M.; Im, J.S.; Ha, P.; Kim, J.S.; Shin, T.Y. Colonization of Metarhizium anisopliae on the surface of pine tree logs: A promising biocontrol strategy for the Japanese pine sawyer, Monochamus alternatus. Fungal Biol. 2020, 124, 125–134. [Google Scholar] [CrossRef]
- Pan, Y.S.; Xu, F.Y.; Han, Z.M. Controlling on Monochamus alternatus larva with Scleroderma guani and carried fungus. J. Jiangsu For. Sci. Technol. 2009, 36, 11–14. [Google Scholar]
- He, X.; Chen, S.; Yang, X.; Huang, J.; Huang, B.; Cai, S. The investigation of Metarhizium anisopliae in forest soil in Fujian and Jiangxi provinces and pathogenicity against Monochamus alternatus. Mycosystema 2007, 2, 289–294. [Google Scholar] [CrossRef]
- Yin, Y.; Song, Z.; Xie, N.; Wang, Z. Nutrition requirement and culture characteristic of Metarhizium anisopliae strain CQMa117. Chin. J. Biol. Control. 2010, 26, 206. [Google Scholar] [CrossRef]
- Guo, Y.; Carballar-Lejarazú, R.; Sheng, L.; Fang, Y.; Wang, S.; Liang, G.; Hu, X.; Wang, R.; Zhang, F.; Wu, S. Identification and characterization of aminopeptidase-N as a binding protein for Cry3Aa in the midgut of Monochamus alternatus (Coleoptera: Cerambycidae). J. Econ. Entomol. 2020, 113, 2259–2268. [Google Scholar] [CrossRef] [PubMed]
- Valero-Jimenez, C.A.; Wiegers, H.; Zwaan, B.J.; Koenraadt, C.J.; van Kan, J.A. Genes involved in virulence of the entomopathogenic fungus Beauveria bassiana. J. Invertebr. Pathol. 2016, 133, 41–49. [Google Scholar] [CrossRef]
- Peng, Z.; Huang, S.; Chen, J.; Li, N.; Wei, Y.; Nawaz, A.; Deng, S. An update of a green pesticide: Metarhizium anisopliae. All Life 2022, 15, 1141–1159. [Google Scholar] [CrossRef]
- Cai, N.; Liu, R.; Yan, D.; Zhang, N.; Zhu, K.; Zhang, D.; Nong, X.; Tu, X.; Zhang, Z.; Wang, G. Bioinformatics Analysis and Functional Characterization of the CFEM Proteins of Metarhizium anisopliae. J. Fungi 2022, 8, 661. [Google Scholar] [CrossRef]
- Wang, C.; St Leger, R.J. The MAD1 adhesin of Metarhizium anisopliae links adhesion with blastospore production and virulence to insects, and the MAD2 adhesin enables attachment to plants. Eukaryot. Cell 2007, 6, 808–816. [Google Scholar] [CrossRef]
- Zacharuk, R.Y. Fine structure of the fungus Metarrhizium anisopliae infecting three species of larval Elateridae (Coleoptera) II. Conidial Germ tubes and appressoria. J. Invertebr. Pathol. 1970, 15, 81–91. [Google Scholar] [CrossRef]
- Wen, Z.; Tian, H.; Xia, Y.; Jin, K. MaPmt1, a protein O-mannosyltransferase, contributes to virulence through governing the appressorium turgor pressure in Metarhizium acridum. Fungal Genet. Biol. 2020, 145, 103480. [Google Scholar] [CrossRef]
- Krieger de Moraes, C.; Schrank, A.; Vainstein, M.H. Regulation of extracellular chitinases and proteases in the entomopathogen and acaricide Metarhizium anisopliae. Curr. Microbiol. 2003, 46, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Jeong, S.G.; Choi, I.S.; Yang, J.E.; Lee, K.H.; Kim, J.; Kim, J.C.; Kim, J.S.; Park, H.W. Mechanisms of Insecticidal Action of Metarhizium anisopliae on Adult Japanese Pine Sawyer Beetles (Monochamus alternatus). ACS Omega 2020, 5, 25312–25318. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Wu, H.; Liu, Z.; Wang, Z.; Huang, B. G-Protein Subunit Galpha(i) in Mitochondria, MrGPA1, Affects Conidiation, Stress Resistance, and Virulence of Entomopathogenic Fungus Metarhizium robertsii. Front. Microbiol. 2020, 11, 1251. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Widemann, E.; Bernard, G.; Lesot, A.; Pinot, F.; Pedrini, N.; Keyhani, N.O. CYP52X1, representing new cytochrome P450 subfamily, displays fatty acid hydroxylase activity and contributes to virulence and growth on insect cuticular substrates in entomopathogenic fungus Beauveria bassiana. J. Biol. Chem. 2012, 287, 13477–13486. [Google Scholar] [CrossRef]
- Wanchoo, A.; Lewis, M.W.; Keyhani, N.O. Lectin mapping reveals stage-specific display of surface carbohydrates in in vitro and haemolymph-derived cells of the entomopathogenic fungus Beauveria bassiana. Microbiology 2009, 155, 3121–3133. [Google Scholar] [CrossRef]
- James, P.; Kershaw, M.; Reynolds, S.; Charnley, A. Inhibition of desert locust (Schistocerca gregaria) Malpighian tubule fluid secretion by destruxins, cyclic peptide toxins from the insect pathogenic fungus Metarhizium anisopliae. J. Insect Physiol. 1993, 39, 797–804. [Google Scholar] [CrossRef]
- Pucheta Díaz, M.; Flores Macías, A.; Rodríguez Navarro, S.; De la Torre, M. Mechanism of action of entomopathogenic fungi. Interciencia 2006, 31, 856–860. [Google Scholar]
- Kim, J.C.; Lee, M.R.; Kim, S.; Park, S.E.; Lee, S.J.; Shin, T.Y.; Kim, W.J.; Kim, J. Transcriptome Analysis of the Japanese Pine Sawyer Beetle, Monochamus alternatus, Infected with the Entomopathogenic Fungus Metarhizium anisopliae JEF-197. J. Fungi 2021, 7, 373. [Google Scholar] [CrossRef]
- Zhang, W.; Meng, J.; Ning, J.; Qin, P.; Zhou, J.; Zou, Z.; Wang, Y.; Jiang, H.; Ahmad, F.; Zhao, L.; et al. Differential immune responses of Monochamus alternatus against symbiotic and entomopathogenic fungi. Sci. China Life Sci. 2017, 60, 902–910. [Google Scholar] [CrossRef][Green Version]
- Yokoi, K.; Koyama, H.; Minakuchi, C.; Tanaka, T.; Miura, K. Antimicrobial peptide gene induction, involvement of Toll and IMD pathways and defense against bacteria in the red flour beetle, Tribolium castaneum. Results Immunol. 2012, 2, 72–82. [Google Scholar] [CrossRef]
- Chu, X.; Yang, M.; Yu, L.; Xie, H.; Liu, J.; Wu, S.; Zhang, F.; Hu, X. Double-strand RNAs targeting MaltRelish and MaltSpz reveals potential targets for pest management of Monochamus alternatus. Pestic. Biochem. Physiol. 2023, 194, 105495. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Xu, W.; Lv, G.; Yuan, H.; Zhang, Q.H.; Wickham, J.D.; Xu, L.; Zhang, L. Associated bacteria of a pine sawyer beetle confer resistance to entomopathogenic fungi via fungal growth inhibition. Environ. Microbiome 2022, 17, 47. [Google Scholar] [CrossRef]
- Wang, C.; St Leger, R.J. A collagenous protective coat enables Metarhizium anisopliae to evade insect immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 6647–6652. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Peng, G.; Xia, Y. The adenylate cyclase gene MaAC is required for virulence and multi-stress tolerance of Metarhizium acridum. BMC Microbiol. 2012, 12, 163. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.B.; Yu, S.F.; Wang, C.L.; Wang, L. cAMP Signalling Pathway in Biocontrol Fungi. Curr. Issues Mol. Biol. 2022, 44, 2622–2634. [Google Scholar] [CrossRef]
- Guan, Y.; Wang, D.Y.; Ying, S.H.; Feng, M.G. A novel Ras GTPase (Ras3) regulates conidiation, multi-stress tolerance and virulence by acting upstream of Hog1 signaling pathway in Beauveria bassiana. Fungal Genet. Biol. 2015, 82, 85–94. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, G.; Ying, S.H.; Feng, M.G. Adenylate cyclase orthologues in two filamentous entomopathogens contribute differentially to growth, conidiation, pathogenicity, and multistress responses. Fungal Biol. 2014, 118, 422–431. [Google Scholar] [CrossRef]
- Modesto, I.; Mendes, A.; Carrasquinho, I.; Miguel, C.M. Molecular defense response of pine trees (Pinus spp.) to the parasitic nematode Bursaphelenchus xylophilus. Cells 2022, 11, 3208. [Google Scholar] [CrossRef]
- Van Loon, L. Systemic Induced Resistance. In Mechanisms of Resistance to Plant Diseases; Slusarenko, A.J., Fraser, R.S.S., van Loon, L.C., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 521–574. [Google Scholar]
- Ross, A.F. Localized acquired resistance to plant virus infection in hypersensitive hosts. Virology 1961, 14, 329–339. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Kamle, M.; Borah, R.; Bora, H.; Jaiswal, A.K.; Singh, R.K.; Kumar, P. Systemic acquired resistance (SAR) and induced systemic resistance (ISR): Role and mechanism of action against phytopathogens. In Fungal Biotechnology and Bioengineering; Springer: Cham, Switzerland, 2020; pp. 457–470. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Wang, C.; Wang, H.; Chen, H.; Tang, M. Pine wilt disease alters soil properties and root-associated fungal communities in Pinus tabulaeformis forest. Plant Soil 2016, 404, 237–249. [Google Scholar] [CrossRef]
- Romera, F.J.; Garcia, M.J.; Lucena, C.; Martinez-Medina, A.; Aparicio, M.A.; Ramos, J.; Alcantara, E.; Angulo, M.; Perez-Vicente, R. Induced Systemic Resistance (ISR) and Fe Deficiency Responses in Dicot Plants. Front. Plant Sci. 2019, 10, 287. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, J.; Xia, R.; Li, D.; Wang, F. Antioxidant processes involving epicatechin decreased symptoms of pine wilt disease. Front. Plant Sci. 2022, 13, 1015970. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Vicente, C.S.L.; Barbosa, P.; Espada, M.; Glick, B.R.; Mota, M.; Oliveira, S. Evidence for the involvement of ACC deaminase from Pseudomonas putida UW4 in the biocontrol of pine wilt disease caused by Bursaphelenchus xylophilus. BioControl 2012, 58, 427–433. [Google Scholar] [CrossRef]
- Peng, Y.; Tang, Y.; Li, D.; Ye, J. The Growth-Promoting and Colonization of the Pine Endophytic Pseudomonas abietaniphila for Pine Wilt Disease Control. Microorganisms 2024, 12, 1089. [Google Scholar] [CrossRef]
- Sun, M.; Liang, C.; Fu, X.; Liu, G.; Zhong, Y.; Wang, T.; Tang, G.; Li, P. Nematocidal activity and biocontrol efficacy of endophytic Bacillus velezensis Pt-RP9 from Pinus tabuliformis against pine wilt disease caused by Bursaphelenchus xylophilus. Biol. Control. 2024, 196, 105579. [Google Scholar] [CrossRef]
- Boller, T. The plant hormone ethylene. In Ethylene in Pathogenesis and Disease Resistance; Mattoo, A.K., Suttle, J.C., Eds.; CRC: Boca Raton, FL, USA, 1991; pp. 297–301. [Google Scholar]
- Trobacher, C.P. Ethylene and programmed cell death in plants. Botany 2009, 87, 757–769. [Google Scholar] [CrossRef]
- Nunes da Silva, M.; Pintado, M.E.; Sarmento, B.; Stamford, N.P.; Vasconcelos, M.W. A biofertilizer with diazotrophic bacteria and a filamentous fungus increases Pinus pinaster tolerance to the pinewood nematode (Bursaphelenchus xylophilus). Biol. Control 2019, 132, 72–80. [Google Scholar] [CrossRef]
- Ponpandian, L.N.; Rim, S.O.; Shanmugam, G.; Jeon, J.; Park, Y.H.; Lee, S.K.; Bae, H. Phylogenetic characterization of bacterial endophytes from four Pinus species and their nematicidal activity against the pine wood nematode. Sci. Rep. 2019, 9, 12457. [Google Scholar] [CrossRef] [PubMed]
- Robinet, C.; Raffin, A.; Jactel, H.; Kersaudy, E.; Deuffic, P.; Clopeau, A.; Hotte, H.; Kleinhentz, M.; Robin, C.; Roux, G. Improving monitoring and management methods is of the utmost importance in countries at risk of invasion by the pinewood nematode. Ann. For. Sci. 2024, 81, 16. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, X.; Li, Y.; Wei, P.; Sun, N.; Wen, X.; Liu, Z.; Li, D.; Feng, Y.; Zhang, X. Differences between microbial communities of Pinus species having differing level of resistance to the pine wood nematode. Microb. Ecol. 2022, 84, 1245–1255. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Liu, X.; Chen, J.; Wang, B.; Li, Y.; Ye, Y.; Ma, W.; Ma, L. Effects on community composition and function Pinus massoniana infected by Bursaphelenchus xylophilus. BMC Microbiol. 2022, 22, 157. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, J.; Liang, S.; Zhu, W.; Li, H. Changes in Phyllosphere Microbial Communities of Pinus tabuliformis after Infestation by Bursaphelenchus xylophilus. Forests 2023, 14, 179. [Google Scholar] [CrossRef]
- Jia, J.; Chen, L.; Yu, W.; Cai, S.; Su, S.; Xiao, X.; Tang, X.; Jiang, X.; Chen, D.; Fang, Y. The novel nematicide chiricanine A suppresses Bursaphelenchus xylophilus pathogenicity in Pinus massoniana by inhibiting Aspergillus and its secondary metabolite, sterigmatocystin. Front. Plant Sci. 2023, 14, 1257744. [Google Scholar] [CrossRef]
- Vicente, C.S.; Nascimento, F.; Espada, M.; Barbosa, P.; Mota, M.; Glick, B.R.; Oliveira, S. Characterization of bacteria associated with pinewood nematode Bursaphelenchus xylophilus. PLoS ONE 2012, 7, e46661. [Google Scholar] [CrossRef]
- Xue, Q.; Xiang, Y.; Wu, X.; Li, M. Bacterial communities and virulence associated with pine wood nematode Bursaphelenchus xylophilus from different Pinus spp. Int. J. Mol. Sci. 2019, 20, 3342. [Google Scholar] [CrossRef]
- Park, A.R.; Jeong, S.I.; Jeon, H.W.; Kim, J.; Kim, N.; Ha, M.T.; Mannaa, M.; Kim, J.; Lee, C.W.; Min, B.S.; et al. A Diketopiperazine, Cyclo-(L-Pro-L-Ile), Derived From Bacillus thuringiensis JCK-1233 Controls Pine Wilt Disease by Elicitation of Moderate Hypersensitive Reaction. Front. Plant Sci. 2020, 11, 1023. [Google Scholar] [CrossRef]
- Nakashima, H.; Eguchi, N.; Uesugi, T.; Yamashita, N.; Matsuda, Y. Effect of ectomycorrhizal composition on survival and growth of Pinus thunbergii seedlings varying in resistance to the pine wilt nematode. Trees 2015, 30, 475–481. [Google Scholar] [CrossRef]
- Chu, H.; Tang, M.; Wang, H.; Wang, C. Pinewood nematode infection alters root mycoflora of Pinus tabulaeformis Carr. J. Appl. Microbiol. 2018, 125, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Wang, C.; Li, Z.; Wang, H.; Xiao, Y.; Chen, J.; Tang, M. The dark septate endophytes and ectomycorrhizal fungi effect on Pinus tabulaeformis Carr. seedling growth and their potential effects to pine wilt disease resistance. Forests 2019, 10, 140. [Google Scholar] [CrossRef]
- Chu, H.; Wang, H.; Zhang, Y.; Li, Z.; Wang, C.; Dai, D.; Tang, M. Inoculation With Ectomycorrhizal Fungi and Dark Septate Endophytes Contributes to the Resistance of Pinus spp. to Pine Wilt Disease. Front. Microbiol. 2021, 12, 687304. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.K.; Prakash, A.; Johri, B.N. Induced systemic resistance (ISR) in plants: Mechanism of action. Indian J. Microbiol. 2007, 47, 289–297. [Google Scholar] [CrossRef]
- Van Wees, S.C.; Luijendijk, M.; Smoorenburg, I.; Van Loon, L.C.; Pieterse, C.M. Rhizobacteria-mediated induced systemic resistance (ISR) in Arabidopsis is not associated with a direct effect on expression of known defense-related genes but stimulates the expression of the jasmonate-inducible gene Atvsp upon challenge. Plant Mol. Biol. 1999, 41, 537–549. [Google Scholar] [CrossRef]
- Pieterse, C.M.; van Wees, S.C.; Hoffland, E.; van Pelt, J.A.; van Loon, L.C. Systemic resistance in Arabidopsis induced by biocontrol bacteria is independent of salicylic acid accumulation and pathogenesis-related gene expression. Plant Cell 1996, 8, 1225–1237. [Google Scholar] [CrossRef]
- De Meyer, G.; Capieau, K.; Audenaert, K.; Buchala, A.; Métraux, J.; Höfte, M. Nanogram amounts of salicylic acid produced by the rhizobacterium Pseudomonas aeruginosa 7NSK2 activate the systemic acquired resistance pathway in bean. Mol. Plant-Microbe Interact. 1999, 12, 450–458. [Google Scholar] [CrossRef]
- Yu, Y.; Gui, Y.; Li, Z.; Jiang, C.; Guo, J.; Niu, D. Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants 2022, 11, 386. [Google Scholar] [CrossRef]
- Tang, J.; Gu, X.; Liu, J.; He, Z. Roles of small RNAs in crop disease resistance. Stress Biol. 2021, 1, 6. [Google Scholar] [CrossRef]
- Yan, J.; Qiu, R.; Wang, K.; Liu, Y.; Zhang, W. Enhancing alfalfa resistance to Spodoptera herbivory by sequestering microRNA396 expression. Plant Cell Rep. 2023, 42, 805–819. [Google Scholar] [CrossRef]
- Gao, Y.; Feng, B.; Gao, C.; Zhang, H.; Wen, F.; Tao, L.; Fu, G.; Xiong, J. The evolution and functional roles of miR408 and its targets in plants. Int. J. Mol. Sci. 2022, 23, 530. [Google Scholar] [CrossRef] [PubMed]
- Rumyantsev, S.D.; Veselova, S.V.; Burkhanova, G.F.; Alekseev, V.Y.; Maksimov, I.V. Bacillus subtilis 26D triggers induced systemic resistance against Rhopalosiphum padi L. by regulating the expression of genes AGO, DCL and microRNA in bread spring wheat. Microorganisms 2023, 11, 2983. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Fan, Z.; Li, Z.; Niu, D.; Li, Y.; Zheng, M.; Wang, Q.; Jin, H.; Guo, J. Bacillus cereus AR156 triggers induced systemic resistance against Pseudomonas syringae pv. tomato DC3000 by suppressing miR472 and activating CNLs-mediated basal immunity in Arabidopsis. Mol. Plant Pathol. 2020, 21, 854–870. [Google Scholar] [CrossRef] [PubMed]
- Rosli, R.; Amiruddin, N.; Ab Halim, M.A.; Chan, P.; Chan, K.; Azizi, N.; Morris, P.E.; Leslie Low, E.; Ong-Abdullah, M.; Sambanthamurthi, R. Comparative genomic and transcriptomic analysis of selected fatty acid biosynthesis genes and CNL disease resistance genes in oil palm. PLoS ONE 2018, 13, e0194792. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Kanzaki, N.; Futai, K. How different is induced host resistance against the pine wood nematode, Bursaphelenchus xylophilus, by two avirulent microbes? Nematology 2006, 8, 435–442. [Google Scholar] [CrossRef]
- Linthorst, H.J.; Van Loon, L. Pathogenesis-related proteins of plants. Crit. Rev. Plant Sci. 1991, 10, 123–150. [Google Scholar] [CrossRef]
- Kim, N.; Jeon, H.W.; Mannaa, M.; Jeong, S.I.; Kim, J.; Kim, J.; Lee, C.; Park, A.R.; Kim, J.C.; Seo, Y.S. Induction of resistance against pine wilt disease caused by Bursaphelenchus xylophilus using selected pine endophytic bacteria. Plant Pathol. 2018, 68, 434–444. [Google Scholar] [CrossRef]
- Han, G.; Mannaa, M.; Kim, N.; Jeon, H.W.; Jung, H.; Lee, H.H.; Kim, J.; Park, J.; Park, A.R.; Kim, J.C.; et al. Response of Pine Rhizosphere Microbiota to Foliar Treatment with Resistance-Inducing Bacteria against Pine Wilt Disease. Microorganisms 2021, 9, 688. [Google Scholar] [CrossRef]
- Ayaz, M.; Li, C.; Ali, Q.; Zhao, W.; Chi, Y.; Shafiq, M.; Ali, F.; Yu, X.; Yu, Q.; Zhao, J. Bacterial and fungal biocontrol agents for plant disease protection: Journey from lab to field, current status, challenges, and global perspectives. Molecules 2023, 28, 6735. [Google Scholar] [CrossRef]
- Wang, R.; Dong, L.; Chen, Y.; Wang, S.; Qu, L. Third generation genome sequencing reveals that endobacteria in Nematophagous fungi Esteya vermicola contain multiple genes encoding for Nematicidal proteins. Front. Microbiol. 2022, 13, 842684. [Google Scholar] [CrossRef]
- Cheng, C.; Qin, J.; Wu, C.; Lei, M.; Wang, Y.; Zhang, L. Suppressing a plant-parasitic nematode with fungivorous behavior by fungal transformation of a Bt cry gene. Microb. Cell Factories 2018, 17, 116. [Google Scholar] [CrossRef] [PubMed]
- Sajid, M.; Geng, C.; Li, M.; Wang, Y.; Liu, H.; Zheng, J.; Peng, D.; Sun, M. Whole-genome analysis of Bacillus thuringiensis revealing partial genes as a source of novel Cry toxins. Appl. Environ. Microbiol. 2018, 84, e00277-18. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.H.; Choi, B.Y.; Kim, D.S.; Han, H.; Kim, Y.-H.; Shim, D. Temporal transcriptome profiling of Pinus densiflora infected with pine wood nematode reveals genetically programmed changes upon pine wilt disease. Phytopathology 2024, 114, 982–989. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Li, Y.; Ma, L.; Li, D.; Zhang, W.; Feng, Y.; Liu, Z.; Wang, X.; Wen, X.; Zhang, X. Transcriptomic response of Pinus massoniana to infection stress from the pine wood nematode Bursaphelenchus xylophilus. Stress Biol. 2023, 3, 50. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, J.; Ye, L.; Liu, N.; Wang, F. Methyl jasmonate induced tolerance effect of Pinus koraiensis to Bursaphelenchus xylophilus. Pest Manag. Sci. 2025, 81, 80–92. [Google Scholar] [CrossRef]
- Cardoso, J.M.; Manadas, B.; Abrantes, I.; Robertson, L.; Arcos, S.C.; Troya, M.T.; Navas, A.; Fonseca, L. Pine wilt disease: What do we know from proteomics? BMC Plant Biol. 2024, 24, 98. [Google Scholar] [CrossRef]
- Li, J.; Zou, C.; Xu, J.; Ji, X.; Niu, X.; Yang, J.; Huang, X.; Zhang, K.Q. Molecular mechanisms of nematode-nematophagous microbe interactions: Basis for biological control of plant-parasitic nematodes. Annu. Rev. Phytopathol. 2015, 53, 67–95. [Google Scholar] [CrossRef]
- Wang, X.; Li, G.; Zou, C.; Ji, X.; Liu, T.; Zhao, P.; Liang, L.; Xu, J.; An, Z.; Zheng, X. Bacteria can mobilize nematode-trapping fungi to kill nematodes. Nat. Commun. 2014, 5, 5776. [Google Scholar] [CrossRef]
- Zheng, H.; Chen, T.; Li, W.; Hong, J.; Xu, J.; Yu, Z. Endosymbiotic bacteria within the nematode-trapping fungus Arthrobotrys musiformis and their potential roles in nitrogen cycling. Front. Microbiol. 2024, 15, 1349447. [Google Scholar] [CrossRef]
- Zhang, H.; Wei, Z.; Zhang, J.; Liu, X. Classification of dendrocola nematode-trapping fungi. J. For. Res. 2021, 32, 1295–1304. [Google Scholar] [CrossRef]
- Jia, H.; Xia, R.; Zhang, R.; Liang, G.; Zhuang, Y.; Zhou, Y.; Li, D.; Wang, F. Transcriptome analysis highlights the influence of temperature on hydrolase and traps in nematode-trapping fungi. Front. Microbiol. 2024, 15, 1384459. [Google Scholar] [CrossRef] [PubMed]
- Saiki, H.; Saito, T.; Yoneda, K.; Umi, M.; Uchida, K.; Yamanaka, K. Biological control of the pine-wood nematode by spraying a nematode-trapping fungus. J. Jpn. For. Soc. 1984, 66, 30–32. [Google Scholar] [CrossRef]
- Wingfield, M.J. Fungi associated with the pine wood nematode, Bursaphelenchus xylophilus, and cerambycid beetles in Wisconsin. Mycologia 1987, 79, 325–328. [Google Scholar] [CrossRef]
- Kano, S.; Aimi, T.; Masumoto, S.; Kitamoto, Y.; Morinaga, T. Physiology and molecular characteristics of a pine wilt nematode-trapping fungus, Monacrosporium megalosporum. Curr. Microbiol. 2004, 49, 158–164. [Google Scholar] [CrossRef]
- Kubátová, A.; Novotný, D.; Práil, K.; Mrácek, Z. The nematophagous hyphomycete Esteya vermicola found in the Czech Republic. Czech Mycol. 2000, 52, 227–235. [Google Scholar] [CrossRef]
- Wang, C.Y.; Fang, Z.M.; Sun, B.S.; Gu, L.J.; Zhang, K.Q.; Sung, C.K. High infectivity of an endoparasitic fungus strain, Esteya vermicola, against nematodes. J. Microbiol. 2008, 46, 380–389. [Google Scholar] [CrossRef]
- Wang, X.; Wang, T.; Wang, J.; Guan, T.; Li, H. Morphological, molecular and biological characterization of Esteya vermicola, a nematophagous fungus isolated from intercepted wood packing materials exported from Brazil. Mycoscience 2014, 55, 367–377. [Google Scholar] [CrossRef]
- Meng, Q.; Shi, X.; Meng, F.; Feng, X.; Sun, J. Isolation of an Acremonium sp. from a screening of 52 seawater fungal isolates and preliminary characterization of its growth conditions and nematicidal activity. Biotechnol. Lett. 2012, 34, 1847–1850. [Google Scholar] [CrossRef]
- Dong, J.; Zhou, Y.; Li, R.; Zhou, W.; Li, L.; Zhu, Y.; Huang, R.; Zhang, K. New nematicidal azaphilones from the aquatic fungus Pseudohalonectria adversaria YMF1. 01019. FEMS Microbiol. Lett. 2006, 264, 65–69. [Google Scholar] [CrossRef]
- Yuan, Y.; Xiang, M.; Xi, P.; Jiang, Z. Screening of nematicidal isolates from the endophytic fungi of Quisqualis indica and optimization of culture conditions for the isolate. Chin. J. Biol. Control 2010, 26, 474. [Google Scholar] [CrossRef]
- Proença, D.N.; Francisco, R.; Santos, C.V.; Lopes, A.; Fonseca, L.; Abrantes, I.M.; Morais, P.V. Diversity of bacteria associated with Bursaphelenchus xylophilus and other nematodes isolated from Pinus pinaster trees with pine wilt disease. PLoS ONE 2010, 5, e15191. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ponpandian, L.N.; Kim, H.; Jeon, J.; Hwang, B.S.; Lee, S.K.; Park, S.C.; Bae, H. Distribution and diversity of bacterial endophytes from four Pinus species and their efficacy as biocontrol agents for devastating pine wood nematodes. Sci. Rep. 2019, 9, 12461. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, C.; Guo, Q.; Li, L.; Guo, D. Identification and culture condition study of marine actinomycete HT-8 with nematicidal activity against pine wood nematode. J. Microbiol. 2016, 6, 18. [Google Scholar] [CrossRef]
- Zhang, W.; Wu, X.; Wang, Y. Nematicidal activity of bacteria against Bursaphelenchus xylophilus and its fermentation and culture characteristics. Biotechnol. Bull. 2019, 35, 76. [Google Scholar] [CrossRef]
- Xu, H.; Xu, J.; Zhang, L.; Lin, H. Nematicidal activity of Bacillus thuringiensis to Bursaphelenchus xylophilus. Chin. J. Biol. Control 2010, 26, 85. [Google Scholar]
- Zhao, Y.; Yuan, Z.; Wang, S.; Wang, H.; Chao, Y.; Sederoff, R.R.; Sederoff, H.; Yan, H.; Pan, J.; Peng, M.; et al. Gene sdaB is involved in the Nematocidal activity of Enterobacter ludwigii AA4 against the pine wood nematode Bursaphelenchus xylophilus. Front. Microbiol. 2022, 13, 870519. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).