
Microbial Community Response and Assembly Process of Yellow Sand Matrix in a Desert Marginal Zone Under Morchella Cultivation
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Overview of the Experimental Area
2.2. Materials
2.3. Methods
2.3.1. Experimental Design
2.3.2. Sample Collection
2.3.3. Determination of Physical and Chemical Properties
2.3.4. Extraction and Sequencing of Soil Microbial DNA
2.3.5. Real-Time Quantitative PCR (qPCR) Analysis [26]
2.3.6. Bioinformatic Analysis
2.4. Data Analysis and Statistics
3. Results
3.1. Yield of Morchella
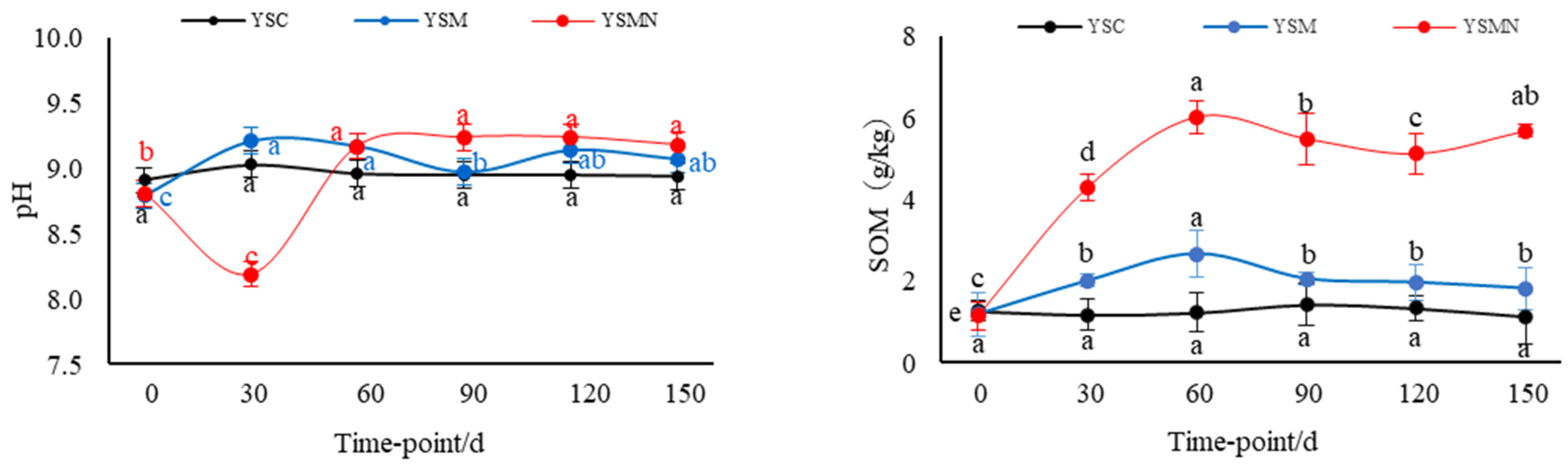
3.2. Physicochemical Properties of the Yellow Sand Substrate
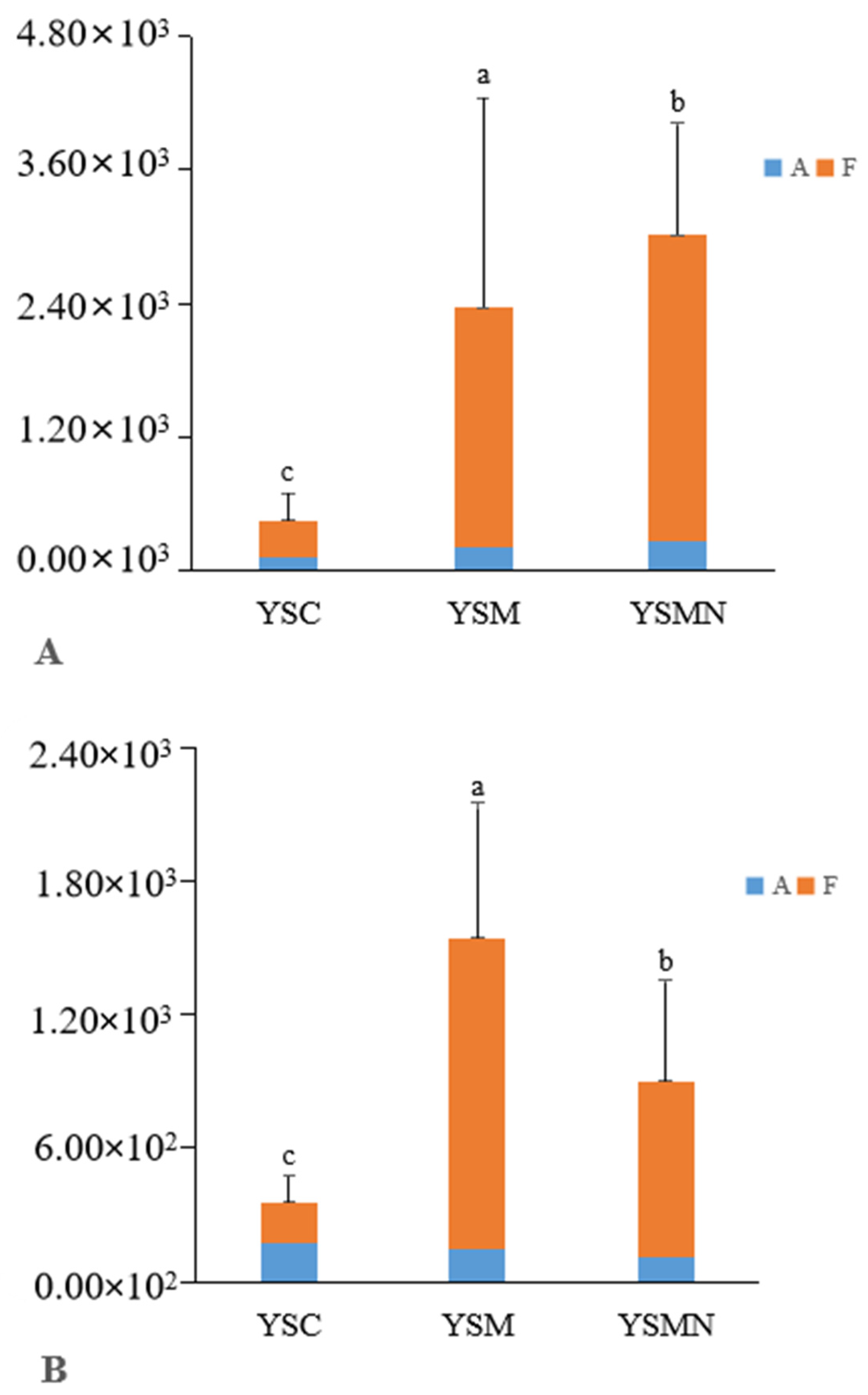
3.3. Bacterial 16S and Fungal ITS Content
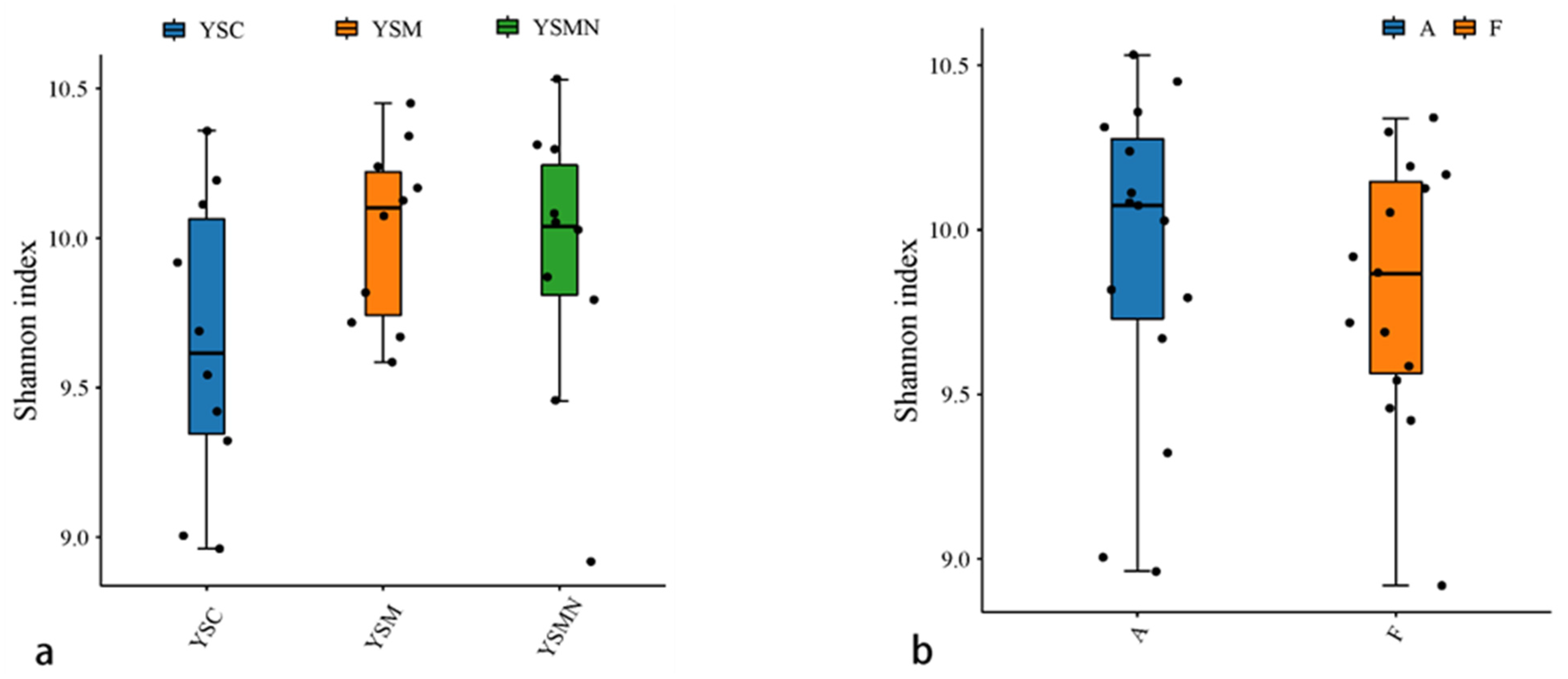
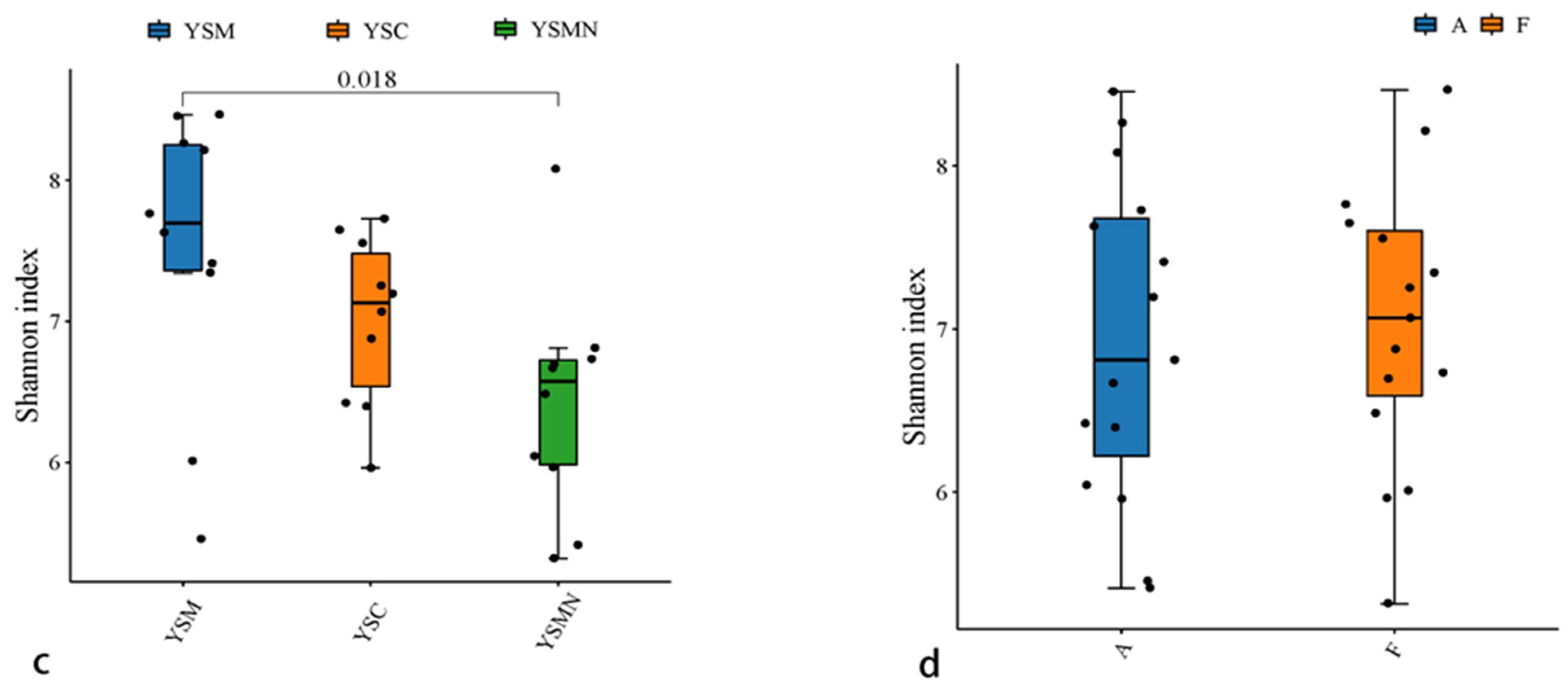
3.4. Microbial Community Diversity Analysis
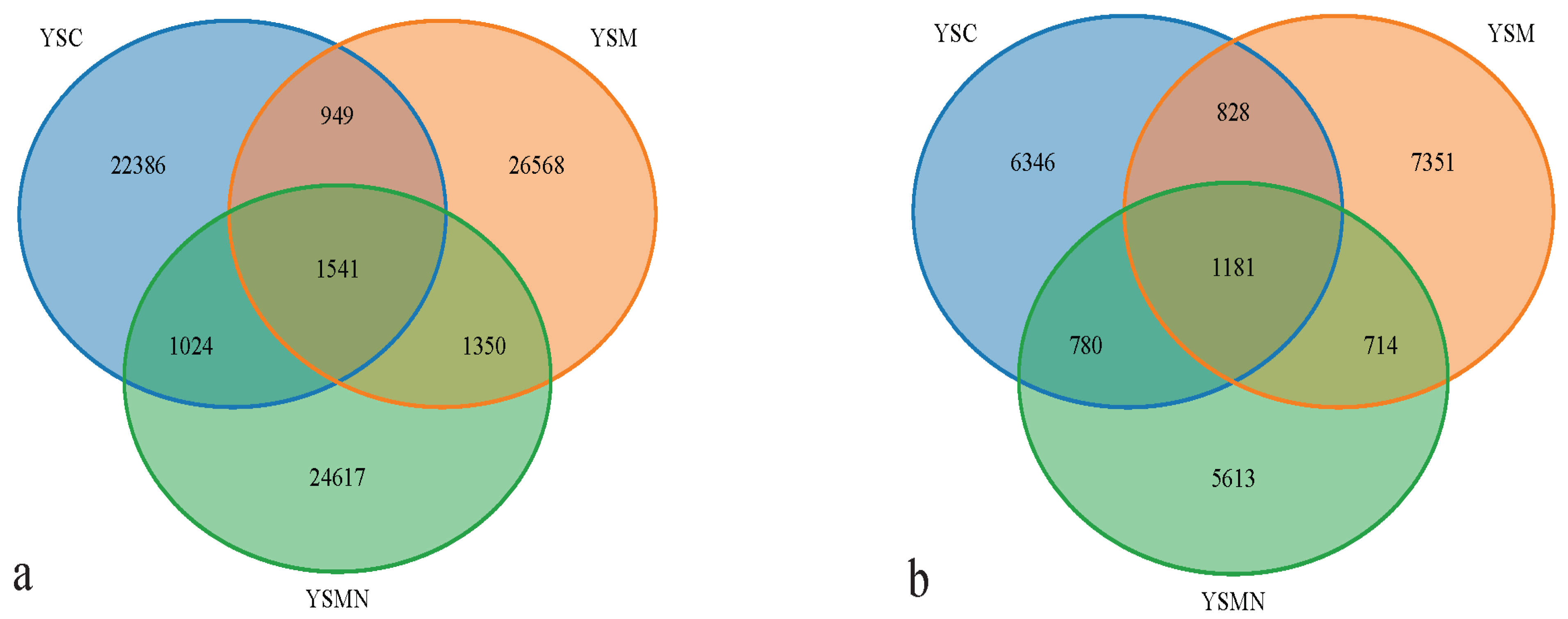
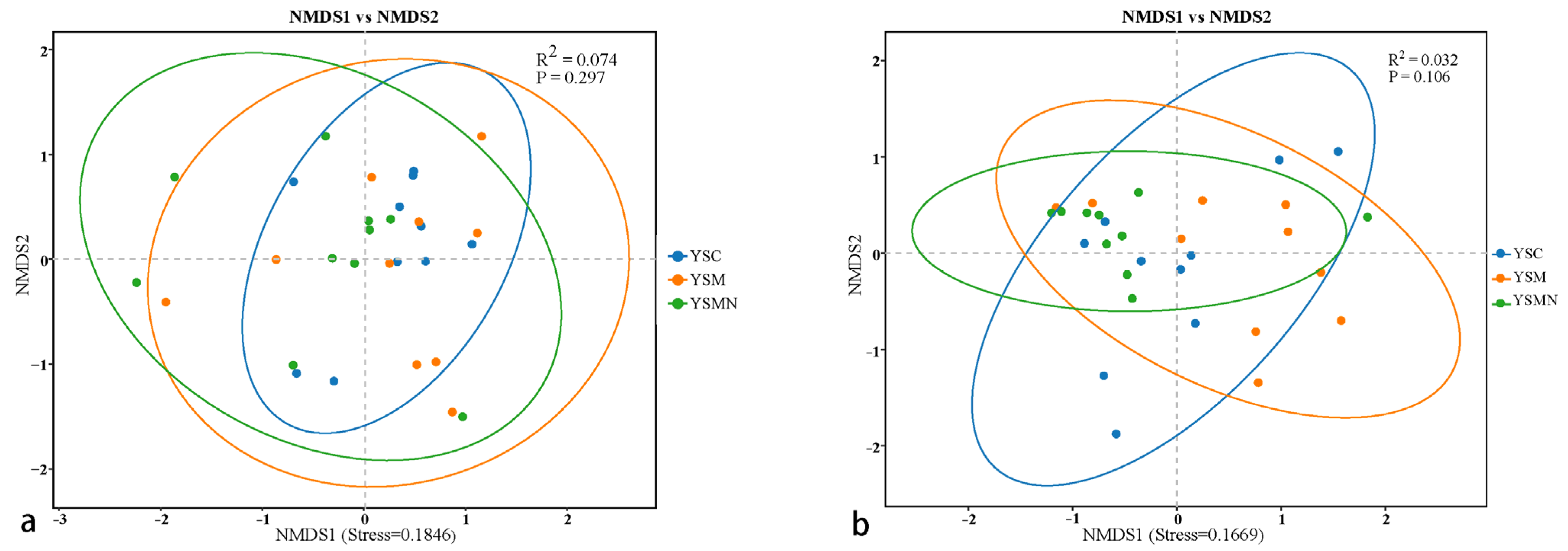
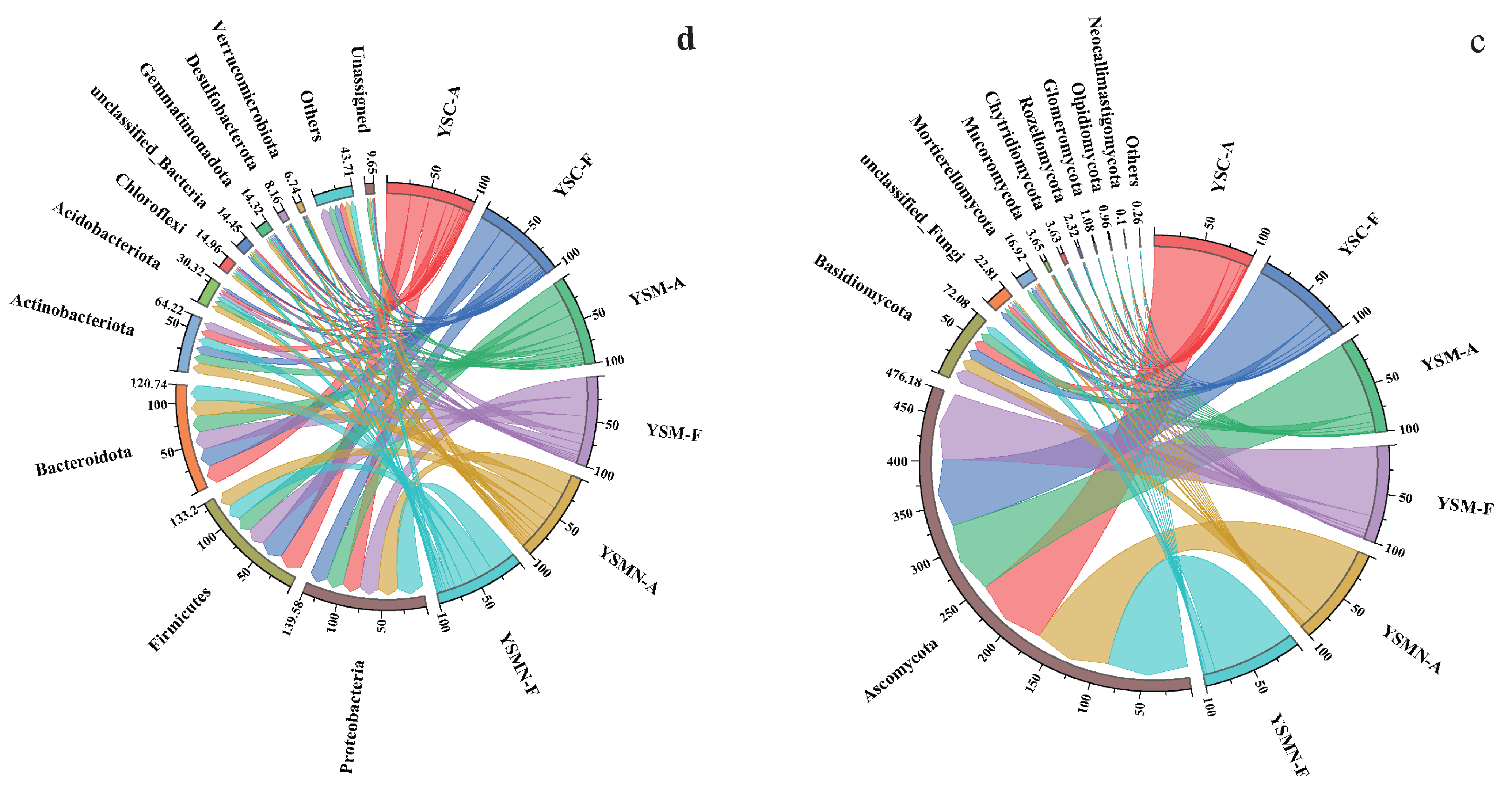
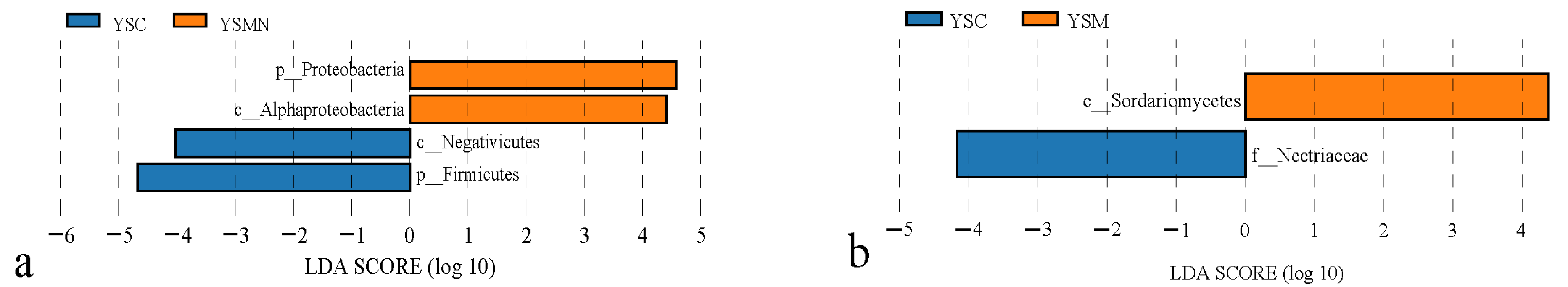
3.5. Microbial Community Compositional Variation Analysis
3.6. Correlation Analysis Between the Microbial Community and Environmental Factors of Yellow Sand Substrate
3.7. Microbial Community Correlation Network Analysis
3.8. Microbial Community Assembly Processes Based on iCAMP Modeling
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Haider, R.; Agnello, L.; Shah, S.M.; Sufyan, M.; Khan, N.; Nazir, A.; Ciaccio, M.; Rehman, S. Evaluating the antioxidant, anti-inflammatory, and neuroprotective potential of fruiting body and mycelium extracts from edible yellow morel (Morchella esculenta L. Pers.). J. Food Sci. 2025, 90, e17619. [Google Scholar] [CrossRef]
- Wang, D.D.; Yin, Z.Q.; Ma, L.K.; Han, L.; Chen, Y.; Pan, W.J.; Gong, K.; Gao, Y.Y.; Yang, X.X.; Chen, L.Y.; et al. Polysaccharide MCP extracted from Morchella esculenta reduces atherosclerosis in LDLR-deficient mice. Food Funct. 2021, 12, 4842–4854. [Google Scholar] [CrossRef] [PubMed]
- Ramya, H.; Ravikumar, K.S.; Fathimathu, Z.; Janardhanan, K.K.; Ajith, T.A.; Shah, M.A.; Farooq, R.; Reshi, Z.A. Morel mushroom, Morchella from Kashmir Himalaya: A potential source of therapeutically useful bioactives that possess free radical scavenging, anti-inflammatory, and arthritic edema-inhibiting activities. Drug Chem. Toxicol. 2021, 45, 10–11. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Fang, D.L.; Benard, M.K.; Chen, X.; Wu, X.; Du, J.X.; Yang, Q.; Chen, H.; Zheng, H.H.; An, X.x.; et al. Analysis of umami taste substances of morel mushroom (Morchella sextelata) hydrolysates derived from different enzymatic systems. Food Chem. 2021, 362, 130192. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Y.; Zou, G.; Bao, D.P.; Wu, Y.Y. Current Advances in the Functional Genes of Edible and Medicinal Fungi: Research Techniques, Functional Analysis, and Prospects. J. Fungi 2024, 10, 311. [Google Scholar] [CrossRef]
- Pildain, M.B.; Visnovsky, S.B.; Barroetaveña, C. Phylogenetic diversity of true morels (Morchella), the main edible non-timber product from native Patagonian forests of Argentina. Fungal Biol. 2014, 118, 755–763. [Google Scholar] [CrossRef]
- Yu, F.M.; Jayawardena, R.S.; Luangharn, T.; Zeng, X.Y.; Li, C.J.; Bao, S.X.; Ba, H.; Zhou, D.Q.; Tang, S.M.; Hyde, K.D.; et al. Species diversity of fungal pathogens on cultivated mushrooms: A case study on morels (Morchella, Pezizales). Fungal Divers. 2024, 125, 157–220. [Google Scholar] [CrossRef]
- Li, Y.T.; Chen, H.Y.; Zhang, X. Cultivation, nutritional value, bioactive compounds of morels, and their health benefits: A systematic review. Front. Nutr. 2023, 10, 1159029. [Google Scholar] [CrossRef]
- He, S.L.; Zhao, K.T.; Ma, L.F.; Yang, J.J.; Chang, Y.W. Effects of different cultivation material formulas on the growth and quality of Morchella spp. Saudi J. Biol. Sci. 2018, 25, 719–723. [Google Scholar] [CrossRef]
- Liu, Y.W.; Guo, B.H.; Bi, X.K.; Sibirina, L.A.; Qi, X.J.; Yu, X.D. Determining why continuous cropping reduces the production of the morel Morchella sextelata. Front. Microbiol. 2022, 13, 903983. [Google Scholar]
- Huang, K.; Li, L.; Wu, W.; Pu, K.; Qi, W.; Qi, J.; Li, M. Enhancing Morchella Mushroom Yield and Quality Through the Amendment of Soil Physicochemical Properties and Microbial Community with Wood Ash. Microorganisms 2024, 12, 2406. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Kohler, A.; Miao, R.; Liu, T.; Zhang, Q.; Zhang, B.; Jiang, L.; Wang, Y.; Xie, L.; Tang, J.; et al. Multi-omic analyses of exogenous nutrient bag decomposition by the black morel Morchella importuna reveal sustained carbon acquisition and transferring. Environ. Microbiol. 2019, 21, 3909–3926. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Du, H.X.; Bonito, G.; Masaphy, S. Editorial: Morels: Physiology, genetics, and interactions with the environment. Front. Microbiol. 2024, 14, 1352719. [Google Scholar] [CrossRef]
- Tan, H.; Yu, Y.; Tang, J.; Liu, T.; Miao, R.; Huang, Z.; Martin, F.M.; Peng, W. Build your own mushroom soil: Microbiota succession and nutritional accumulation in semisynthetic substratum drive the fructification of a soil-saprotrophicmorel. Front. Microbiol. 2021, 12, 656656. [Google Scholar] [CrossRef]
- Xie, Y.H.; Dang, X.H.; Meng, Z.J.; Jiang, H.T.; Li, X.J.; Zhou, R.P.; Zhou, D.D.; Liu, X.X.; Ding, J.; Wu, X.Q.; et al. Wind and sand control by an oasis protective system: A case from the southeastern edge of the Tengger Desert, China. J. Mt. Sci. 2019, 16, 2548–2561. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Liu, T.H.; He, M.J.; Zhang, S.C.; Liao, J.; Lei, T.Z.; Wu, X.; Yang, Y.; Wang, T.; Tan, H. A nationwide study of heavy metal(loid)s in agricultural soils and the soil-grown black morel Morchella sextelata in China. J. Environ. Manag. 2024, 369, 122243. [Google Scholar] [CrossRef]
- Guan, Q.Y.; Wang, L.J.; Wang, L.; Pan, B.T.; Zhao, S.L.; Zheng, Y. Analysis of trace elements (heavy metal based) in the surface soils of a desert–loess transitional zone in the south of the Tengger Desert. Environ. Earth Sci. 2014, 72, 3015–3023. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, S.; Luo, D.; Mao, P.; Rosazlina, R.; Martin, F.; Xu, L. Decline in Morel Production upon Continuous Cropping Is Related to Changes in Soil Mycobiome. J. Fungi 2023, 9, 492. [Google Scholar] [CrossRef]
- Song, H.; Chen, D.; Sun, S.; Li, J.; Tu, M.; Xu, Z.; Gong, R.; Jiang, G. Peach-Morchella intercropping mode affects soil properties and fungal composition. Peer J. 2021, 9, e11705. [Google Scholar] [CrossRef]
- Marco, F.; Daniel, S.; Marco, C.; Silvia, T.; Simone, R. Connect the dots: Sketching out microbiome interactions through networking approaches. Microbiome Res. Rep. 2023, 2, 25. [Google Scholar]
- Das, S.; Kim, J.P.; Nie, M.; Chabbi, A. Soil organic matter in the anthropocene: Role in climate change mitigation, carbon sequestration, and food security. Agric. Ecosyst. Environ. 2024, 375, 109180. [Google Scholar] [CrossRef]
- Wang, J.N.; Pan, Z.; Yu, J.S.; Zhang, Z.; Li, Y.Z. Global assembly of microbial communities. mSystems 2023, 29, e0128922. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L.; Chenu, C.; Kappler, A.; Rillig, M.C.; Fierer, N. The interplay between microbial communities and soil properties. Nat. Rev. Microbiol. 2023, 22, 226–239. [Google Scholar] [CrossRef]
- Bao, S.D. Agrochemical Analysis of Soil Mushrooms; China Agricultural Press: Beijing, China, 2005. [Google Scholar]
- NY/T 1121.11-2006; Soil Testing-Part 3: Mechanical Composition of Soil. Industry Standard of the People’s Republic of China. Standards Press of China: Beijing, China, 2006.
- Yu, Y.; Kang, X.; Liu, T.; Wang, Y.; Tang, J.; Peng, W.; Martin, F.M.; Tan, H. Inoculation of the Morchella importuna mycosphere with Pseudomonas chlororaphis alleviated a soil-borne disease caused by Paecilomyces penicillatus. Biol. Fertil. Soils 2024, 61, 141–161. [Google Scholar] [CrossRef]
- Yue, Y.H.; Hao, H.B.; Wang, Q.; Xiao, T.T.; Zhang, Y.C.; Chen, Q.; Chen, H.; Zhang, J.J. Dynamics of the soil microbial community associated with Morchella cultivation: Diversity, assembly mechanism and yield prediction. Front. Microbiol. 2024, 15, 1345231. [Google Scholar] [CrossRef]
- Li, Z.X.; Li, J.H.; Zhang, Q.; Lu, J.J.; Gao, C.H.; Dong, S.J.; Xu, M.G. Response and Assembly Process of Soil Microbial Communities Under Different Reclamation Measures. Environ. Sci. Huan Jing Ke Xue Huanjing Kexue 2024, 45, 7326–7336. [Google Scholar]
- Yin, W.; Wang, L.; Shang, Q.H.; Li, Y.X.; Sa, W.; Dong, Q.M.; Liang, J. Effects of drought stress and Morchella inoculation on the physicochemical properties, enzymatic activities, and bacterial community of Poa pratensis L. rhizosphere soil. PeerJ 2025, 13, 18793. [Google Scholar] [CrossRef]
- Tan, H.; Liu, T.; Yu, Y.; Tang, J.; Jiang, L.; Martin, F.M.; Peng, W. Morel Production Related to Soil Microbial Diversity and Evenness. Microbiol. Spectr. 2021, 9, e0022921. [Google Scholar] [CrossRef]
- Liu, X.; Liu, J.Z.; Liu, J.; Zhang, J.; Wang, C.L. Determination of the Effects of Pear-Morchella Intercropping Mode on M. sextelata Quality, Yield, and Soil Microbial Community. J. Fungi 2024, 10, 759. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, Q.; Uroz, S.; Gao, T.; Li, J.; He, F.; Rosazlina, R.; Martin, F.; Xu, L. The cultivation regimes of Morchella sextelata trigger shifts in the community assemblage and ecological traits of soil bacteria. Front. Microbiol. 2023, 14, 1257905. [Google Scholar] [CrossRef]
- Cailleau, G.; Hanson, B.T.; Cravero, M.; Zhioua, S.; Hilpish, P.; Ruiz, C.; Robinson, A.J.; Kelliher, J.M.; Morales, D.; Gallegos-Graves, L.V.; et al. Associated bacterial communities, confrontation studies, and comparative genomics reveal important interactions between Morchella with Pseudomonas spp. Front. Fungal Biol. 2023, 4, 1285531. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.M.; Jayawardena, R.S.; Thongklang, N.; Lv, M.L.; Zhu, X.T.; Zhao, Q. Morel production associated with soil nitrogen-fixing and nitrifying microorganisms. J. Fungi 2022, 8, 299. [Google Scholar] [CrossRef] [PubMed]
- Lohberger, A.; Spangenberg, J.E.; Ventura, Y.; Bindschedler, S.; Verrecchia, E.P.; Bshary, R.; Junier, P. Effect of organic carbon and nitrogen on the interactions of Morchella spp. and bacteria dispersing on their mycelium. Front. Microbiol. 2019, 10, 124. [Google Scholar] [CrossRef]
- Tang, S.J.; Fan, T.T.; Jin, L.; Pin, L.; Shao, C.X.; Wu, S.L.; Yang, Y.; He, Y.L.; Ren, R.; Xu, J. Soil microbial diversity and functional capacity associated with the production of edible mushroom Stropharia rugosoannulata in croplands. PeerJ 2022, 10, e14130. [Google Scholar] [CrossRef]
- Shen, C.; Xia, L.; Qin, J.F. Kiwifruit-Agaricus blazei intercropping effectively improved yield productivity, nutrient uptake, and rhizospheric bacterial community. Sci. Rep. 2024, 14, 16546. [Google Scholar]
- Moretti, G.L.; Crusciol, C.A.C.; Leite, A.F.M.; Leite, M.F.A.; Momesso, L.; Bossolani, J.W.; Costa, O.Y.A.; Hungria, M.H.; Kuramae, E.E. Diverse bacterial consortia: Key drivers of rhizosoil fertility modulating microbiome functions, plant physiology, nutrition, and Soybean grain yield. Environ. Microbiome 2024, 19, 50. [Google Scholar] [CrossRef]
- Xu, Y.Y.; Tang, J.; Wang, Y.; He, X.L.; Tan, H.; Yu, Y.; Chen, Y.; Peng, W.H. Large-scale commercial cultivation of morels: Current state and perspectives. Appl. Microbiol. Biotechnol. 2022, 106, 4401–4412. [Google Scholar] [CrossRef]
- Duan, Z.M.; Yang, C.C.; Bao, L.Y.; Han, D.; Wang, H.Z.; Zhang, Y.Z.; Liu, H.G.; Yang, S.Q. Morchella esculenta cultivation in fallow paddy fields and drylands affects the diversity of soil bacteria and soil chemical properties. Front. Genet. 2023, 14, 1251695. [Google Scholar] [CrossRef]
- Anil, K.; Prakash, V.S.; Satish, K. Studies on domestication of the true morel Morchella importuna in India. Indian Phytopathol 2024, 77, 301–310. [Google Scholar]
- Qi, Y.; Wang, P.; Liu, Z.S.; Yu, Y.; Tan, X.; Huang, X.; Wen, J.W.; Zhang, W.D. Analysis of Soil Microbial Community Structure and Function in Morchella esculenta Habitats in Jilin Province. Agronomy 2025, 15, 15. [Google Scholar]
- Lian, J.S.; Wang, H.Y.; Deng, Y.; Xu, M.G.; Liu, S.T.; Zhou, B.K.; Kamlesh, J.; Duan, Y.H. Impact of long-term application of manure and inorganic fertilizers on common soil bacteria in different soil types. Agric. Ecosyst. Environ. 2022, 337, 108044. [Google Scholar] [CrossRef]
- Krause, A.E.; Frank, K.A.; Mason, D.M.; Ulanowicz, R.E.; Taylor, W.W. Compartments revealed in food-web structure. Nature 2003, 426, 282–285. [Google Scholar] [CrossRef] [PubMed]
- Morriën, E.; Hannula, S.E.; Snoek, L.B.; Helmsing, N.R.; Zweers, H.; De Hollander, M.; Soto, R.L.; Bouffaud, M.L.; Buée, M.; Dimmers, W.; et al. Soil networks become more connected and take up more carbon as nature restoration progresses. Nat. Commun. 2017, 8, 14349. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Xiong, J.Z.Q.; Liu, L.H.; Wang, F.F.; Cao, Z.W.; Xu, W.F. Microbial interactions strengthen deterministic processes during community assembly in a subtropical estuary. Sci. Total Environ. 2024, 906, 167499. [Google Scholar] [CrossRef]
- He, R.J.; Zeng, J.; Zhao, D.Y.; Wang, S.R.; Wu, Q.L. Decreased spatial variation and deterministic processes of bacterial community assembly in the rhizosphere of Phragmites australis across the Middle-Lower Yangtze plain. Mol. Ecol. 2022, 31, 1180–1195. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Network Indexes | Bacteria | Fungal | ||||
|---|---|---|---|---|---|---|
| YSC | YSM | YSMN | YSC | YSM | YSMN | |
| Total nodes | 171 | 138 | 163 | 164 | 152 | 165 |
| Total links | 345 | 308 | 316 | 352 | 333 | 678 |
| Positive links | 44.9 | 45.4 | 76.5 | 66.7 | 54.9 | 87.4 |
| Average degree (avgK) | 4.04 | 4.46 | 3.88 | 4.29 | 4.38 | 8.22 |
| Average clustering coefficient (avgCC) | 0.15 | 0.13 | 0.12 | 0.17 | 0.16 | 0.23 |
| Average path distance (GD) | 4.62 | 4.45 | 5.45 | 5.07 | 5.01 | 5.89 |
| Transitivity (Trans) | 0.16 | 0.16 | 0.19 | 0.26 | 0.34 | 0.68 |
| Density (D) | 0.02 | 0.03 | 0.02 | 0.03 | 0.03 | 0.05 |
| Category | Assembly Process | YSC | YSM | YSMN |
|---|---|---|---|---|
| Bacteria | Deterministic | 65.44 | 68.89 | 95.56 |
| Stochastic | 35.56 | 31.11 | 2.22 | |
| Fungal | Deterministic | 20.00 | 20.00 | 51.11 |
| Stochastic | 80.00 | 80.00 | 46.67 |
| Category | Treatment | Homogeneous Selection | Heterogeneous Selection | Diffusion Limitation | Homogeneous Diffusion | Non-Dominant Process |
|---|---|---|---|---|---|---|
| Bacteria | YSC | 17.78 | 46.67 | 4.44 | 17.78 | 13.33 |
| YSM | 26.67 | 42.22 | 13.33 | 17.78 | ||
| YSMN | 95.56 | 2.22 | ||||
| Fungal | YSC | 11.11 | 8.89 | 55.56 | 24.44 | |
| YSM | 20.00 | 55.56 | 24.44 | |||
| YSMN | 4.44 | 46.67 | 28.89 | 17.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, C.; Zhang, S.; Zhou, Y.; Tan, H.; Zhang, S.; Wang, T.; Ding, Z.; Liao, J. Microbial Community Response and Assembly Process of Yellow Sand Matrix in a Desert Marginal Zone Under Morchella Cultivation. Microorganisms 2025, 13, 921. https://doi.org/10.3390/microorganisms13040921
Su C, Zhang S, Zhou Y, Tan H, Zhang S, Wang T, Ding Z, Liao J. Microbial Community Response and Assembly Process of Yellow Sand Matrix in a Desert Marginal Zone Under Morchella Cultivation. Microorganisms. 2025; 13(4):921. https://doi.org/10.3390/microorganisms13040921
Chicago/Turabian StyleSu, Cuicui, Shengyin Zhang, Yanfang Zhou, Hao Tan, Shuncun Zhang, Tao Wang, Zhaoyun Ding, and Jie Liao. 2025. "Microbial Community Response and Assembly Process of Yellow Sand Matrix in a Desert Marginal Zone Under Morchella Cultivation" Microorganisms 13, no. 4: 921. https://doi.org/10.3390/microorganisms13040921
APA StyleSu, C., Zhang, S., Zhou, Y., Tan, H., Zhang, S., Wang, T., Ding, Z., & Liao, J. (2025). Microbial Community Response and Assembly Process of Yellow Sand Matrix in a Desert Marginal Zone Under Morchella Cultivation. Microorganisms, 13(4), 921. https://doi.org/10.3390/microorganisms13040921