


Repression of Staphylococcus aureus and Escherichia coli by Lactiplantibacillus plantarum Strain AG10 in Drosophila melanogaster In Vivo Model
 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Antibacterial Activity of LAB Strains (Agar Drop Diffusion Test)
2.3. Antibacterial Activity of LAB Strains in the Fermented Milk
2.4. D. melanogaster Husbandry
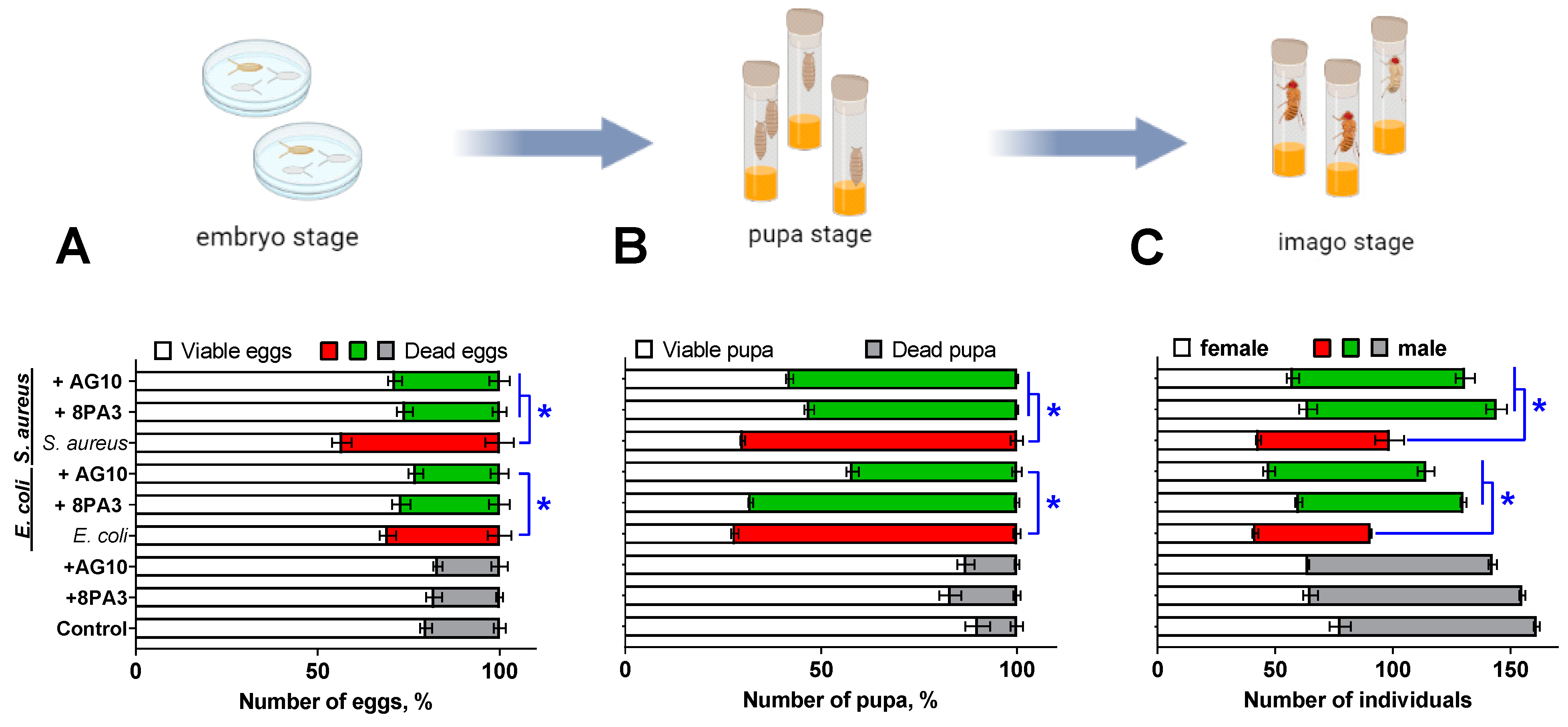
2.5. The Analysis of Flies Viability on Embryonic and Metamorphosis Stages
2.6. Fertility Analysis
2.7. DNA Comet Assay
2.8. SMART Test
2.9. DNA Extraction
2.10. Genome Sequencing and Assembling
2.11. Bioinformatic Analysis
2.12. Statistical Analysis
3. Results
3.1. In Vitro Antagonistic Activity of L. plantarum AG10
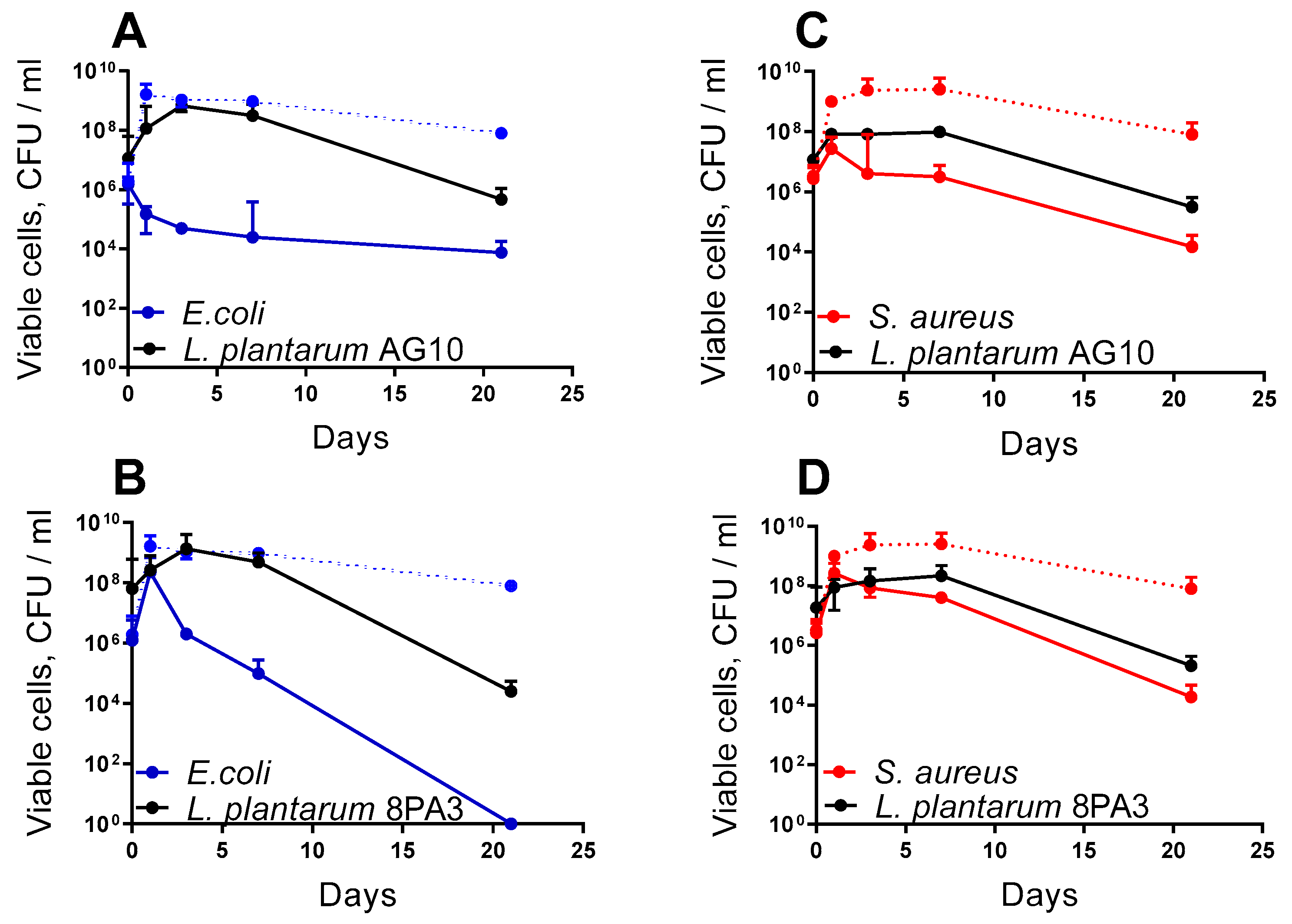
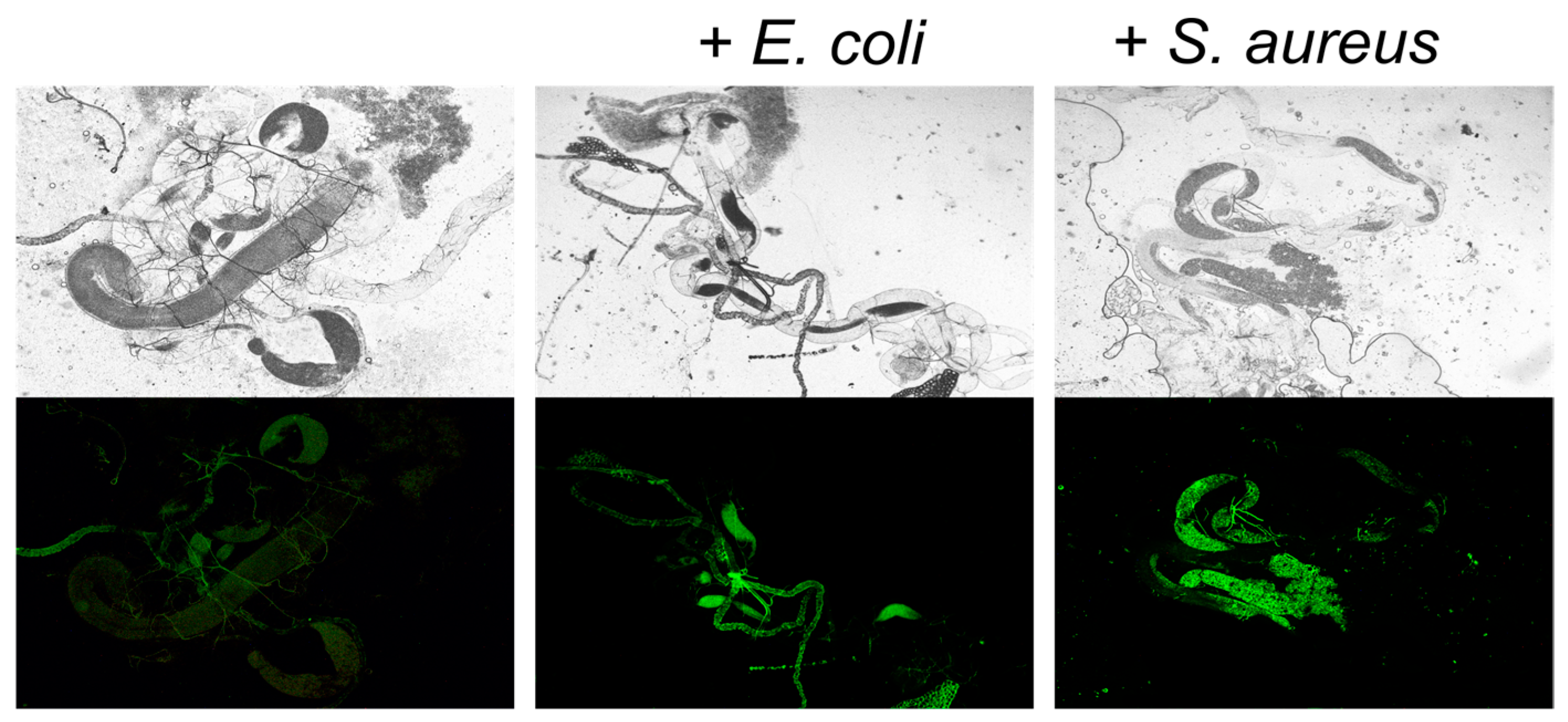
3.2. Antagonistic Activity of Lactobacillus in the Drosophila Model
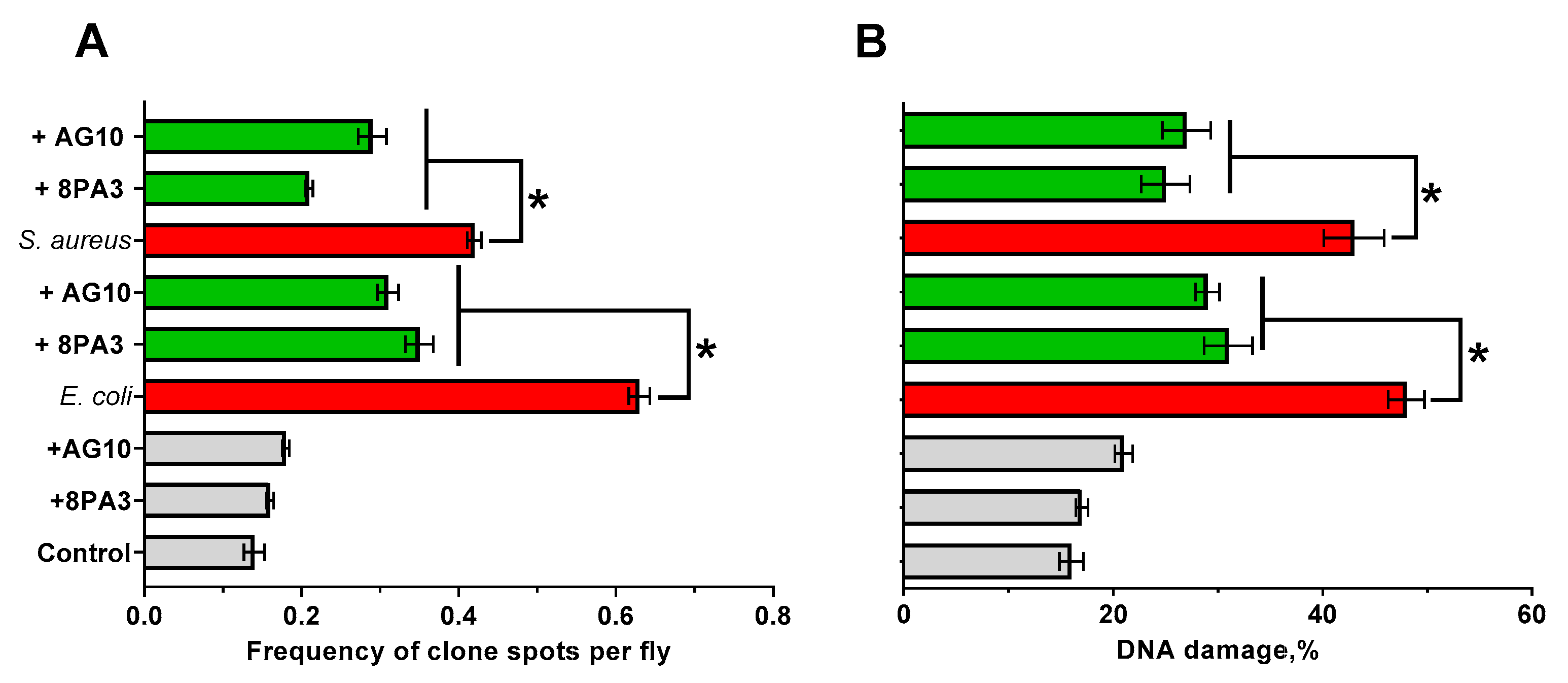
3.3. Genotoxicity of Lactobacillus Strains in the Drosophila Model
3.4. Whole Genome Sequence of L. plantarum AG10
3.5. In Silico Identification of Antimicrobial Peptides and Virulence Factors in LAB Genomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sneeringer, S.; MacDonald, J.M.; Key, N.; McBride, W.D.; Mathews, K. Economics of antibiotic use in US livestock production. USDA Econ. Res. Rep. 2015, 200. [Google Scholar]
- Manaia, C.M.; Vaz-Moreira, I.; Nunes, O.C. Antibiotic resistance in waste water and surface water and human health implications. Emerg. Org. Contam. Hum. Health 2012, 20, 173–212. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Insights into antibiotic resistance through metagenomic approaches. Future Microbiol. 2012, 7, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Smits, C.H.; Li, D.; Patience, J.F.; den Hartog, L. Animal nutrition strategies and options to reduce the use of antimicrobials in animal production. Food Agric. Organ. U. N. 2021, 2021, 184. [Google Scholar]
- Hawkey, P.M. The growing burden of antimicrobial resistance. J. Antimicrob. Chemother. 2008, 62 (Suppl. S1), i1–i9. [Google Scholar] [CrossRef]
- Qiao, M.; Ying, G.G.; Singer, A.C.; Zhu, Y.G. Review of antibiotic resistance in China and its environment. Environ. Int. 2018, 110, 160–172. [Google Scholar] [CrossRef]
- Hinton, M.; Linton, A.H. Control of salmonella infections in broiler chickens by the acid treatment of their feed. Vet. Rec. 1988, 123, 416–421. [Google Scholar] [CrossRef]
- Van Immerseel, F.; De Buck, J.; Boyen, F.; Bohez, L.; Pasmans, F.; Volf, J.; Sevcik, M.; Rychlik, I.; Haesebrouck, F.; Ducatelle, R. Medium-chain fatty acids decrease colonization and invasion through hilA suppression shortly after infection of chickens with Salmonella enterica serovar Enteritidis. Appl. Environ. Microbiol. 2004, 70, 3582–3587. [Google Scholar] [CrossRef]
- Thompson, J.L.; Hinton, M. Antibacterial activity of formic and propionic acids in the diet of hens on Salmonellas in the crop. Br. Poult. Sci. 1997, 38, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Mitsch, P.; Zitterl-Eglseer, K.; Köhler, B.; Gabler, C.; Losa, R.; Zimpernik, I. The effect of two different blends of essential oil components on the proliferation of Clostridium perfringens in the intestines of broiler chickens. Poult. Sci. 2004, 83, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K.A.; Carson, C.F.; Riley, T.V. Antimicrobial activity of essential oils and other plant extracts. J. Appl. Microbiol. 1999, 86, 985–990. [Google Scholar] [CrossRef]
- Hernández, F.; Madrid, J.; García, V.; Orengo, J.; Megías, M.D. Influence of two plant extracts on broilers performance, digestibility, and digestive organ size. Poult. Sci. 2004, 83, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.; Buick, R.; Elliott, C.T. Antibacterial activities of naturally occurring compounds against antibiotic-resistant Bacillus cereus vegetative cells and spores, Escherichia coli, and Staphylococcus aureus. J. Food Prot. 2004, 67, 1774–1778. [Google Scholar] [CrossRef]
- Lewis, K.; Ausubel, F.M. Prospects for plant-derived antibacterials. Nat. Biotechnol. 2006, 24, 1504–1507. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [PubMed]
- Vaou, N.; Stavropoulou, E.; Voidarou, C.; Tsigalou, C.; Bezirtzoglou, E. Towards Advances in Medicinal Plant Antimicrobial Activity: A Review Study on Challenges and Future Perspectives. Microorganisms 2021, 9, 9102041. [Google Scholar] [CrossRef]
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S. Antimicrobial Peptides as Potential Alternatives to Antibiotics in Food Animal Industry. Int. J. Mol. Sci. 2016, 17, 17050603. [Google Scholar] [CrossRef]
- Anee, I.J.; Alam, S.; Begum, R.A.; Shahjahan, R.M.; Khandaker, A.M. The role of probiotics on animal health and nutrition. J. Basic Appl. Zool. 2021, 82, 52. [Google Scholar] [CrossRef]
- Alagawany, M.; Abd El-Hack, M.E.; Farag, M.R.; Sachan, S.; Karthik, K.; Dhama, K. The use of probiotics as eco-friendly alternatives for antibiotics in poultry nutrition. Environ. Sci. Pollut. Res. Int. 2018, 25, 10611–10618. [Google Scholar] [CrossRef]
- Vasile, N.; Ghindea, R.; Vassu, T. Probiotics—An alternative treatment for various diseases. Roum. Arch. Microbiol. Immunol. 2011, 70, 54–59. [Google Scholar]
- Hotel, A.C.P.; Cordoba, A. Health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria. Prevention 2001, 5, 1–10. [Google Scholar]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Simon, O.; Jadamus, A.; Vahjen, W. Probiotic feed additives-effectiveness and expected modes of action. J. Anim. Feed Sci. 2001, 10, 51–68. [Google Scholar] [CrossRef]
- Tan, L.T.; Chan, K.G.; Lee, L.H.; Goh, B.H. Streptomyces Bacteria as Potential Probiotics in Aquaculture. Front. Microbiol. 2016, 7, 79. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 9091021. [Google Scholar] [CrossRef]
- Soccol, C.R.; de Souza Vandenberghe, L.P.; Spier, M.R.; Medeiros, A.P.; Yamaguishi, C.T.; De Dea Lindner, J.; Pandey, A. The potential of probiotics: A review. Food Technol. Biotechnol. 2010, 48, 413–434. [Google Scholar]
- Hammes, W.P.; Hertel, C. The genera lactobacillus and carnobacterium. Prokaryotes 2006, 4, 320–403. [Google Scholar]
- Wyszyńska, A.; Kobierecka, P.; Bardowski, J.; Jagusztyn-Krynicka, E.K. Lactic acid bacteria—20 years exploring their potential as live vectors for mucosal vaccination. Appl. Microbiol. Biotechnol. 2015, 99, 2967–2977. [Google Scholar] [CrossRef]
- Cui, C.; Shen, C.J.; Jia, G.; Wang, K.N. Effect of dietary Bacillus subtilis on proportion of Bacteroidetes and Firmicutes in swine intestine and lipid metabolism. Genet. Mol. Res. 2013, 12, 1766–1776. [Google Scholar] [CrossRef]
- Sanders, M.E.; Morelli, L.; Tompkins, T.A. Sporeformers as Human Probiotics: Bacillus, Sporolactobacillus, and Brevibacillus. Compr. Rev. Food Sci. Food Saf. 2003, 2, 101–110. [Google Scholar] [CrossRef]
- Moshood, Y.; Tengku, A. Lactic Acid Bacteria: Bacteriocin Producer: A Mini Review. IOSR J. Pharm. 2013, 3, 44–55. [Google Scholar]
- Piard, J.C.; Desmazeaud, M. Inhibiting Factors Produced by Lactic-acid Bacteria 1. Oxygen Metabolites and Catabolism end-Products. Lait 1991, 71, 525–541. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Ancarani, F.; Del Prete, M.S.; Fortuna, M.; Scalise, G. In Vitro activities of polycationic peptides alone and in combination with clinically used antimicrobial agents against Rhodococcus equi. Antimicrob. Agents Chemother. 1999, 43, 2093–2096. [Google Scholar] [CrossRef] [PubMed]
- Vaara, M.; Porro, M. Group of peptides that act synergistically with hydrophobic antibiotics against gram-negative enteric bacteria. Antimicrob. Agents Chemother. 1996, 40, 1801–1805. [Google Scholar] [CrossRef] [PubMed]
- Martín, V.; Cárdenas, N.; Ocaña, S.; Marín, M.; Arroyo, R.; Beltrán, D.; Badiola, C.; Fernández, L.; Rodríguez, J.M. Rectal and vaginal eradication of Streptococcus agalactiae (GBS) in pregnant women by using Lactobacillus salivarius CECT 9145, a target-specific probiotic strain. Nutrients 2019, 11, 810. [Google Scholar] [CrossRef]
- Abatemarco Júnior, M.; Sandes, S.H.C.; Ricci, M.F.; Arantes, R.M.E.; Nunes, Á.C.; Nicoli, J.R.; Neumann, E. Protective effect of Lactobacillus diolivorans 1Z, isolated from Brazilian kefir, against Salmonella enterica serovar typhimurium in experimental murine models. Front. Microbiol. 2018, 9, 2856. [Google Scholar] [CrossRef]
- Zheng, Y.-D.; Pan, Y.; He, K.; Li, N.; Yang, D.; Du, G.-F.; Ge, R.; He, Q.-Y.; Sun, X. SPD_1495 contributes to capsular polysaccharide synthesis and virulence in Streptococcus pneumoniae. Msystems 2020, 5, e00025-20. [Google Scholar] [CrossRef]
- Singh, A.K.; Tiwari, U.P.; Berrocoso, J.D.; Dersjant-Li, Y.; Awati, A.; Jha, R. Effects of a combination of xylanase, amylase and protease, and probiotics on major nutrients including amino acids and non-starch polysaccharides utilization in broilers fed different level of fibers. Poult. Sci. 2019, 98, 5571–5581. [Google Scholar] [CrossRef]
- Corcionivoschi, N.; Drinceanu, D.; Pop, I.M.; Stack, D.; Stef, L.; Julean, C.; Bourke, B. The effect of probiotics on animal health. Anim. Sci. Biotechnol. 2010, 43, 35–41. [Google Scholar]
- Yoo, J.Y.; Kim, S.S. Probiotics and Prebiotics: Present Status and Future Perspectives on Metabolic Disorders. Nutrients 2016, 8, 173. [Google Scholar] [CrossRef]
- Linares, D.M.; Ross, P.; Stanton, C. Beneficial Microbes: The pharmacy in the gut. Bioengineered 2016, 7, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Oelschlaeger, T.A. Mechanisms of probiotic actions—A review. Int. J. Med. Microbiol. 2010, 300, 57–62. [Google Scholar] [CrossRef]
- Lee, I.K.; Kye, Y.C.; Kim, G.; Kim, H.W.; Gu, M.J.; Umboh, J.; Maaruf, K.; Kim, S.W.; Yun, C.H. Stress, Nutrition, and Intestinal Immune Responses in Pigs—A Review. Asian-Australas. J. Anim. Sci. 2016, 29, 1075–1082. [Google Scholar] [CrossRef]
- Bajagai, Y.S.; Klieve, A.V.; Dart, P.J.; Bryden, W.L. Probiotics in Animal Nutrition: Production, Impact and Regulation; FAO: Rome, Italy, 2016. [Google Scholar]
- Thelestam, M.; Frisan, T. Cytolethal distending toxins. Rev. Physiol. Biochem. Pharm. 2004, 152, 111–133. [Google Scholar] [CrossRef]
- Chumduri, C.; Gurumurthy, R.K.; Zietlow, R.; Meyer, T.F. Subversion of host genome integrity by bacterial pathogens. Nat. Rev. Mol. Cell Biol. 2016, 17, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Deplanche, M.; Mouhali, N.; Nguyen, M.T.; Cauty, C.; Ezan, F.; Diot, A.; Raulin, L.; Dutertre, S.; Langouet, S.; Legembre, P.; et al. Staphylococcus aureus induces DNA damage in host cell. Sci. Rep. 2019, 9, 7694. [Google Scholar] [CrossRef] [PubMed]
- Prazdnova, E.V.; Mazanko, M.S.; Chistyakov, V.A.; Bogdanova, A.A.; Refeld, A.G.; Kharchenko, E.Y.; Chikindas, M.L. Antimutagenic Activity as a Criterion of Potential Probiotic Properties. Probiotics Antimicrob. Proteins 2022, 14, 1094–1109. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Y.; Ye, L.; Wang, C. The anti-cancer effects and mechanisms of lactic acid bacteria exopolysaccharides in vitro: A review. Carbohydr. Polym. 2021, 253, 117308. [Google Scholar] [CrossRef] [PubMed]
- Gavrilova, E.; Anisimova, E.; Gabdelkhadieva, A.; Nikitina, E.; Vafina, A.; Yarullina, D.; Bogachev, M.; Kayumov, A. Newly isolated lactic acid bacteria from silage targeting biofilms of foodborne pathogens during milk fermentation. BMC Microbiol. 2019, 19, 248. [Google Scholar] [CrossRef]
- Nikitina, E.; Petrova, T.; Vafina, A.; Ezhkova, A.; Nait Yahia, M.; Kayumov, A. Textural and Functional Properties of Skimmed and Whole Milk Fermented by Novel Lactiplantibacillus plantarum AG10 Strain Isolated from Silage. Fermentation 2022, 8, 290. [Google Scholar] [CrossRef]
- Nikitina, E.; Petrova, T.; Sungatullina, A.; Bondar, O.; Kharina, M.; Mikshina, P.; Gavrilova, E.; Kayumov, A. The Profile of Exopolysaccharides Produced by Various Lactobacillus Species from Silage during Not-Fat Milk Fermentation. Fermentation 2023, 9, 197. [Google Scholar] [CrossRef]
- Nikitina, E. Potential use of Lactiplantibacillus plantarum strains for fermentation of fat milk. IOP Conf. Ser. Earth Environ. Sci. 2022, 978, 012047. [Google Scholar] [CrossRef]
- Trinder, M.; Daisley, B.A.; Dube, J.S.; Reid, G. as a High-Throughput Model for Host-Microbiota Interactions. Front. Microbiol. 2017, 8, 751. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, B.; Miguel-Aliaga, I. The digestive tract of Drosophila melanogaster. Annu. Rev. Genet. 2013, 47, 377–404. [Google Scholar]
- Sahu, S.; Jaysingh, P.; Mishra, M. Drosophila melanogaster as an In Vivo Model for the Investigation of Host-Microbiota Interaction. In Prebiotics, Probiotics and Nutraceuticals; Springer: Berlin/Heidelberg, Germany, 2022; pp. 275–300. [Google Scholar]
- Broderick, N.A.; Buchon, N.; Lemaitre, B. Microbiota-induced changes in Drosophila melanogaster host gene expression and gut morphology. mBio 2014, 5, e01117-14. [Google Scholar] [CrossRef]
- Vallet-Gely, I.; Lemaitre, B.; Boccard, F. Bacterial strategies to overcome insect defences. Nat. Rev. Microbiol. 2008, 6, 302–313. [Google Scholar] [CrossRef]
- Hedges, L.M.; Johnson, K.N. Induction of host defence responses by Drosophila C virus. J. Gen. Virol. 2008, 89, 1497–1501. [Google Scholar] [CrossRef]
- Chamilos, G.; Lewis, R.E.; Hu, J.; Xiao, L.; Zal, T.; Gilliet, M.; Halder, G.; Kontoyiannis, D.P. Drosophila melanogaster as a model host to dissect the immunopathogenesis of zygomycosis. Proc. Natl. Acad. Sci. USA 2008, 105, 9367–9372. [Google Scholar] [CrossRef]
- Chen, K.; Luan, X.; Liu, Q.; Wang, J.; Chang, X.; Snijders, A.M.; Mao, J.-H.; Secombe, J.; Dan, Z.; Chen, J.-H. Drosophila histone demethylase KDM5 regulates social behavior through immune control and gut microbiota maintenance. Cell Host Microbe 2019, 25, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Slankster, E.; Lee, C.; Hess, K.M.; Odell, S.; Mathew, D. Effect of gut microbes on olfactory behavior of Drosophila melanogaster larva. Bios 2019, 90, 227. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Pirofski, L.-A. The damage-response framework of microbial pathogenesis. Nat. Rev. Microbiol. 2003, 1, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Cui, Y.; Qu, X. Exopolysaccharides of lactic acid bacteria: Structure, bioactivity and associations: A review. Carbohydr. Polym. 2019, 207, 317–332. [Google Scholar] [CrossRef] [PubMed]
- Storelli, G.; Defaye, A.; Erkosar, B.; Hols, P.; Royet, J.; Leulier, F. Lactobacillus plantarum promotes Drosophila systemic growth by modulating hormonal signals through TOR-dependent nutrient sensing. Cell Metab. 2011, 14, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Poinsot, P.; Penhoat, A.; Mitchell, M.; Sauvinet, V.; Meiller, L.; Louche-Pélissier, C.; Meugnier, E.; Ruiz, M.; Schwarzer, M.; Michalski, M.-C. Probiotic from human breast milk, Lactobacillus fermentum, promotes growth in animal model of chronic malnutrition. Pediatr. Res. 2020, 88, 374–381. [Google Scholar] [CrossRef]
- Yamada, R.; Deshpande, S.A.; Bruce, K.D.; Mak, E.M.; William, W.J. Microbes promote amino acid harvest to rescue undernutrition in Drosophila. Cell Rep. 2015, 10, 865–872. [Google Scholar] [CrossRef]
- Consuegra, J.; Grenier, T.; Baa-Puyoulet, P.; Rahioui, I.; Akherraz, H.; Gervais, H.; Parisot, N.; Da Silva, P.; Charles, H.; Calevro, F. Drosophila-associated bacteria differentially shape the nutritional requirements of their host during juvenile growth. PLoS Biol. 2020, 18, e3000681. [Google Scholar] [CrossRef]
- Newell, P.D.; Douglas, A.E. Interspecies interactions determine the impact of the gut microbiota on nutrient allocation in Drosophila melanogaster. Appl. Environ. Microbiol. 2014, 80, 788–796. [Google Scholar] [CrossRef]
- Deshpande, S.A.; Carvalho, G.B.; Amador, A.; Phillips, A.M.; Hoxha, S.; Lizotte, K.J.; Ja, W.W. Quantifying Drosophila food intake: Comparative analysis of current methodology. Nat. Methods 2014, 11, 535–540. [Google Scholar] [CrossRef]
- Leitão-Gonçalves, R.; Carvalho-Santos, Z.; Francisco, A.P.; Fioreze, G.T.; Anjos, M.; Baltazar, C.; Elias, A.P.; Itskov, P.M.; Piper, M.D.; Ribeiro, C. Commensal bacteria and essential amino acids control food choice behavior and reproduction. PLoS Biol. 2017, 15, e2000862. [Google Scholar] [CrossRef]
- Jones, R.M.; Luo, L.; Ardita, C.S.; Richardson, A.N.; Kwon, Y.M.; Mercante, J.W.; Alam, A.; Gates, C.L.; Wu, H.; Swanson, P.A. Symbiotic lactobacilli stimulate gut epithelial proliferation via Nox-mediated generation of reactive oxygen species. EMBO J. 2013, 32, 3017–3028. [Google Scholar] [CrossRef]
- Jones, R.M.; Desai, C.; Darby, T.M.; Luo, L.; Wolfarth, A.A.; Scharer, C.D.; Ardita, C.S.; Reedy, A.R.; Keebaugh, E.S.; Neish, A.S. Lactobacilli modulate epithelial cytoprotection through the Nrf2 pathway. Cell Rep. 2015, 12, 1217–1225. [Google Scholar] [CrossRef]
- Kasozi, K.I.; Bukenya, A.; Eze, E.D.; Kasolo, J.; Tayebwa, D.S.; Ssempijja, F.; Suubo, J.; Tamale, A.; Echoru, I.; Ntulume, I. Low concentrations of Lactobacillus rhamnosus GG (Yoba®) are safe in male Drosophila melanogaster. BMC Res. Notes 2019, 12, 269. [Google Scholar] [CrossRef]
- Hegan, P.; Mooseker, M.; Handelsman, J.; Miles, J. Effect of probiotic and pathogenic bacteria on Drosophila intestinal pathology. J. Biomol. Tech. 2014, 25, S26. [Google Scholar]
- Ramírez-Camejo, L.; García-Alicea, M.; Maldonado-Morales, G.; Bayman, P. Probiotics may protect Drosophila from infection by Aspergillus flavus. Int. J. Pharm. Sci. Res. 2017, 8, 1624. [Google Scholar]
- Tsapieva, A.; Duplik, N.; Suvorov, A. Structure of plantaricin locus of Lactobacillus plantarum 8P-A3. Benef. Microbes 2011, 2, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Stern, N.J.; Svetoch, E.A.; Eruslanov, B.V.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; Pokhilenko, V.D.; Levchuk, V.P.; Svetoch, O.E.; Seal, B.S. Isolation of a Lactobacillus salivarius strain and purification of its bacteriocin, which is inhibitory to Campylobacter jejuni in the chicken gastrointestinal system. Antimicrob. Agents Chemother. 2006, 50, 3111–3116. [Google Scholar] [CrossRef] [PubMed]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Baidamshina, D.R.; Trizna, E.Y.; Holyavka, M.G.; Bogachev, M.I.; Artyukhov, V.G.; Akhatova, F.S.; Rozhina, E.V.; Fakhrullin, R.F.; Kayumov, A.R. Targeting microbial biofilms using Ficin, a nonspecific plant protease. Sci. Rep. 2017, 7, 46068. [Google Scholar] [CrossRef]
- Sharafutdinov, I.S.; Trizna, E.Y.; Baidamshina, D.R.; Ryzhikova, M.N.; Sibgatullina, R.R.; Khabibrakhmanova, A.M.; Latypova, L.Z.; Kurbangalieva, A.R.; Rozhina, E.V. Klinger-Strobel, M. Antimicrobial Effects of Sulfonyl Derivative of 2(5H)-Furanone against Planktonic and Biofilm Associated Methicillin-Resistant and -Susceptible Staphylococcus aureus. Front. Microbiol. 2017, 8, 2246. [Google Scholar] [CrossRef]
- Tikhomirova, M. Geneticheskii analiz; Izdadelstvo LGU: Leningrad, Russia, 1990. [Google Scholar]
- Adonyeva, N.V.; Menshanov, P.N.; Gruntenko, N. A link between atmospheric pressure and fertility of drosophila laboratory strains. Insects 2021, 12, 947. [Google Scholar] [CrossRef]
- Mukhopadhyay, I.; Chowdhuri, D.K.; Bajpayee, M.; Dhawan, A. Evaluation of in vivo genotoxicity of cypermethrin in Drosophila melanogaster using the alkaline Comet assay. Mutagenesis 2004, 19, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Do Amaral, V.S.; da Silva, R.M.; Reguly, M.L.; de Andrade, H.H.R. Drosophila wing-spot test for genotoxic assessment of pollutants in water samples from urban and industrial origin. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2005, 583, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Kaya, B.; Creus, A.; Yanikoğlu, A.; Cabré, O.; Marcos, R. Use of the Drosophila wing spot test in the genotoxicity testing of different herbicides. Environ. Mol. Mutagen. 2000, 36, 40–46. [Google Scholar] [CrossRef]
- Graf, U.; Frei, H.; Kägi, A.; Katz, A.J.; Würgler, F.E. Thirty compounds tested in the Drosophila wing spot test. Mutat. Res. 1989, 222, 359–373. [Google Scholar] [CrossRef] [PubMed]
- FastQC: A Quality Control Tool for High Throughput Sequence Data [Online]. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 15 March 2023).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Jhong, J.-H.; Yao, L.; Pang, Y.; Li, Z.; Chung, C.-R.; Wang, R.; Li, S.; Li, W.; Luo, M.; Ma, R. dbAMP 2.0: Updated resource for antimicrobial peptides with an enhanced scanning method for genomic and proteomic data. Nucleic Acids Res. 2022, 50, D460–D470. [Google Scholar] [CrossRef]
- Waghu, F.H.; Barai, R.S.; Gurung, P.; Idicula-Thomas, S. CAMP(R3): A database on sequences, structures and signatures of antimicrobial peptides. Nucleic Acids Res. 2016, 44, D1094–D1097. [Google Scholar] [CrossRef]
- Ramond, E.; Jamet, A.; Ding, X.; Euphrasie, D.; Bouvier, C.; Lallemant, L.; He, X.; Arbibe, L.; Coureuil, M.; Charbit, A. Reactive Oxygen Species-Dependent Innate Immune Mechanisms Control Methicillin-Resistant Staphylococcus aureus Virulence in the. mBio 2021, 12, e0027621. [Google Scholar] [CrossRef] [PubMed]
- Needham, A.J.; Kibart, M.; Crossley, H.; Ingham, P.W.; Foster, S.J. Drosophila melanogaster as a model host for Staphylococcus aureus infection. Microbiology 2004, 150 Pt 7, 2347–2355. [Google Scholar] [CrossRef]
- Kim, E.K.; Lee, K.A.; Hyeon, D.Y.; Kyung, M.; Jun, K.Y.; Seo, S.H.; Hwang, D.; Kwon, Y.; Lee, W.J. Bacterial Nucleoside Catabolism Controls Quorum Sensing and Commensal-to-Pathogen Transition in the Drosophila Gut. Cell Host Microbe 2020, 27, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Ott, L.C.; Engelken, M.; Scott, S.M.; McNeill, E.M.; Mellata, M. Model for Gut-Mediated Horizontal Transfer of Narrow- and Broad-Host-Range Plasmids. mSphere 2021, 6, e0069821. [Google Scholar] [CrossRef]
- Fast, D.; Duggal, A.; Foley, E. Monoassociation with Lactobacillus plantarum disrupts intestinal homeostasis in adult Drosophila melanogaster. mBio 2018, 9, e01114-18. [Google Scholar] [CrossRef] [PubMed]
- Kostenko, V.; Mouzykantov, A.; Baranova, N.; Boulygina, E.; Markelova, M.; Khusnutdinova, D.; Trushin, M.; Chernova, O.; Chernov, V. Development of Resistance to Clarithromycin and Amoxicillin-Clavulanic Acid in Lactiplantibacillus plantarum In Vitro Is Followed by Genomic Rearrangements and Evolution of Virulence. Microbiol. Spectr. 2022, 10, e02360-21. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LAB Strain | Growth Inhibition, mm | |||||
|---|---|---|---|---|---|---|
| B. cereus | E. coli | S. aureus | K. pneumoniae | S. marcescens | P. aeruginosa | |
| L. plantarum AG10 | 1.0 ± 0.09 | 2.3 ± 0.20 | 2.8 ± 0.24 | 1.0 ± 0.70 | 3.0 ± 0.24 | 5.0 ± 0.36 |
| L. plantarum 8PA3 | 1.0 ± 0.11 | 1.5 ± 0.13 | 1.5 ± 0.13 | 1.0 ± 0.80 | 1.5 ± 0.60 | 2.5 ± 0.14 |
| Number of Genes | Description |
|---|---|
| 262 | Carbohydrates |
| 188 | Amino acids and derivatives |
| 127 | Protein Metabolism |
| 112 | Cofactors, vitamins, prosthetic groups, pigments |
| 90 | Nucleosides and nucleotides |
| 77 | Cell wall and capsule |
| 65 | DNA metabolism |
| 41 | RNA metabolism |
| 39 | Virulence, disease and defense |
| 38 | Fatty acids, lipids, and isoprenoids |
| 37 | Membrane transport |
| 30 | Phages, prophages, transposable elements, plasmids |
| 21 | Stress response |
| 16 | Regulation and cell signaling |
| 15 | Respiration |
| 14 | Miscellaneous |
| 9 | Nitrogen metabolism |
| 8 | Metabolism of aromatic compounds |
| 6 | Dormancy and sporulation |
| 6 | Phosphorus metabolism |
| 5 | Potassium metabolism |
| 5 | Iron acquisition and metabolism |
| 4 | Cell division and cell cycle |
| 4 | Secondary metabolism |
| 3 | Sulfur metabolism |
| 0 | Photosynthesis |
| 0 | Motility and chemotaxis |
| Protein ID | L. plantarum AG10 Protein Sequence | AMP ID | Identity, % | Alignment Length |
|---|---|---|---|---|
| LLCBMPMO_01958 | MNKMIKNLDVVDASAPISNNKLNGVVGGDAWKNFWSSLRKGFYDGEAGRANPSLINGLKLRRAYSGNQINY | dbAMP_09551 | 92 | 50 |
| dbAMP_02440 | 100 | 21 | ||
| LLCBMPMO_01967 | MKKFLVLRDRELNAISGGVFHAYSARGVRNNYKSAVGPADWVISAVRGFIHG | dbAMP_11717 | 100 | 34 |
| LLCBMPMO_01968 | MLQFEKLQYSRLPQKKLAKISGGFNRGGYNFGKSVRHVVDAIGSVAGIRGILKSIR | dbAMP_08433 | 100 | 56 |
| dbAMP_02258 | 100 | 33 | ||
| dbAMP_02257 | 91 | 33 |
| Protein ID | L. plantarum 8PA3 Protein Sequence | AMP ID | Identity | Alignment Length |
|---|---|---|---|---|
| WP_003641985.1 | MLQFEKLQYSRLPQKKLAKISGGFNRGGYNFGKSVRHVVDAIGSVAGIRGILKSIR | dbAMP_08433 | 100 | 56 |
| dbAMP_02258 | 100 | 33 | ||
| dbAMP_02257 | 91 | 33 | ||
| WP_003643800.1 | MDKFEKISTSNLEKISGGDLTTKLWSSWGYYLGKKARWNLKHPYVQF | dbAMP_06066 | 100 | 28 |
| WP_003643811.1 | MKKFLVLRDRELNAISGGVFHAYSARGVRNNYKSAVGPADWVISAVRGFIHG | dbAMP_11717 | 100 | 34 |
| WP_046947768.1 | MKIQIKSMKQLSNKEMQKIVGGKSSAYSLQMGATAIKQVKKLFKKWGW | dbAMP_07932 | 98 | 48 |
| dbAMP_05363 | 100 | 26 | ||
| dbAMP_00773 | 100 | 23 | ||
| dbAMP_12287 | 100 | 22 | ||
| dbAMP_12286 | 95 | 21 | ||
| dbAMP_02438 | 100 | 17 |
| N | The Protein Precursor | CAMPR3 | DBAASP | |||
|---|---|---|---|---|---|---|
| SVM | Random Forest | DAC | E. coli ATCC 25922 | S. aureus ATCC 25923 | ||
| 1 | LLCBMPMO_01958 | 0.399 | 0.556 | 0.399 | Not Active (0.70) | Not Active (0.78) |
| 2 | LLCBMPMO_01967 | 0.230 | 0.347Koheцфoрмы | 0.230 | Active (0.51) | Not Active (0.61) |
| 3 | LLCBMPMO_01968 | 0.757 | 0.812 | 0.757 | Active (0.53) | Active (0.50) |
| 4 | WP_003641985.1 | 0.716 | 0.757 | 0.812 | Active (0.54) | Not Active (0.51) |
| 5 | WP_003643800.1 | 0.096 | 0.122 | 0.023 | Not Active (0.63) | Not Active (0.69) |
| 6 | WP_003643811.1 | 0.189 | 0.230 | 0.347 | Not Active (0.60) | Not Active (0.62) |
| 7 | WP_046947768.1 | 0.477 | 0.610 | 0.500 | Active (0.74) | Active (0.60) |
| L. plantarum AG10 Protein | Gene Locus | Bacterial Virulence Factor |
|---|---|---|
| ATP-dependent Clp endopeptidase proteolytic subunit ClpP | QBL19_01375 | ATP-dependent Clp protease proteolytic subunit (Listeria) |
| UTP-glucose-1-phosphate uridylyltransferase GalU | QBL19_01520 | UTP-glucose-1-phosphate uridylyltransferase HasC (product); |
| Hyaluronic acid capsule (relative VF) | ||
| (Streptococcus) | ||
| choloylglycine hydrolase | QBL19_03780 | Bile salt hydrolase (Listeria) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gavrilova, E.; Kostenko, V.; Zadorina, I.; Khusnutdinova, D.; Yarullina, D.; Ezhkova, A.; Bogachev, M.; Kayumov, A.; Nikitina, E. Repression of Staphylococcus aureus and Escherichia coli by Lactiplantibacillus plantarum Strain AG10 in Drosophila melanogaster In Vivo Model. Microorganisms 2023, 11, 1297. https://doi.org/10.3390/microorganisms11051297
Gavrilova E, Kostenko V, Zadorina I, Khusnutdinova D, Yarullina D, Ezhkova A, Bogachev M, Kayumov A, Nikitina E. Repression of Staphylococcus aureus and Escherichia coli by Lactiplantibacillus plantarum Strain AG10 in Drosophila melanogaster In Vivo Model. Microorganisms. 2023; 11(5):1297. https://doi.org/10.3390/microorganisms11051297
Chicago/Turabian StyleGavrilova, Elizaveta, Victoria Kostenko, Iva Zadorina, Dilyara Khusnutdinova, Dina Yarullina, Asya Ezhkova, Mikhail Bogachev, Airat Kayumov, and Elena Nikitina. 2023. "Repression of Staphylococcus aureus and Escherichia coli by Lactiplantibacillus plantarum Strain AG10 in Drosophila melanogaster In Vivo Model" Microorganisms 11, no. 5: 1297. https://doi.org/10.3390/microorganisms11051297
APA StyleGavrilova, E., Kostenko, V., Zadorina, I., Khusnutdinova, D., Yarullina, D., Ezhkova, A., Bogachev, M., Kayumov, A., & Nikitina, E. (2023). Repression of Staphylococcus aureus and Escherichia coli by Lactiplantibacillus plantarum Strain AG10 in Drosophila melanogaster In Vivo Model. Microorganisms, 11(5), 1297. https://doi.org/10.3390/microorganisms11051297