

Evidence of High Genetic Diversity and Differences in the Population Diversity of the Eucalyptus Leaf Blight Pathogen Calonectria pseudoreteaudii from Diseased Leaves and Soil in a Plantation in Guangxi, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Calonectria pseudoreteaudii Isolates and DNA Extraction
2.2. Microsatellite Locus Sequencing and Allele Scoring
2.3. Genotype Accumulation Curve
2.4. Population Genetic Diversity Analyses
2.5. Population Structure, Minimum Spanning Network, and Molecular Variance Analyses
2.6. Population Reproduction Mode
3. Results
3.1. Microsatellite Locus Sequencing and Allele Scoring
3.2. Genotype Accumulation Curve
3.3. Population Genetic Diversity Analyses
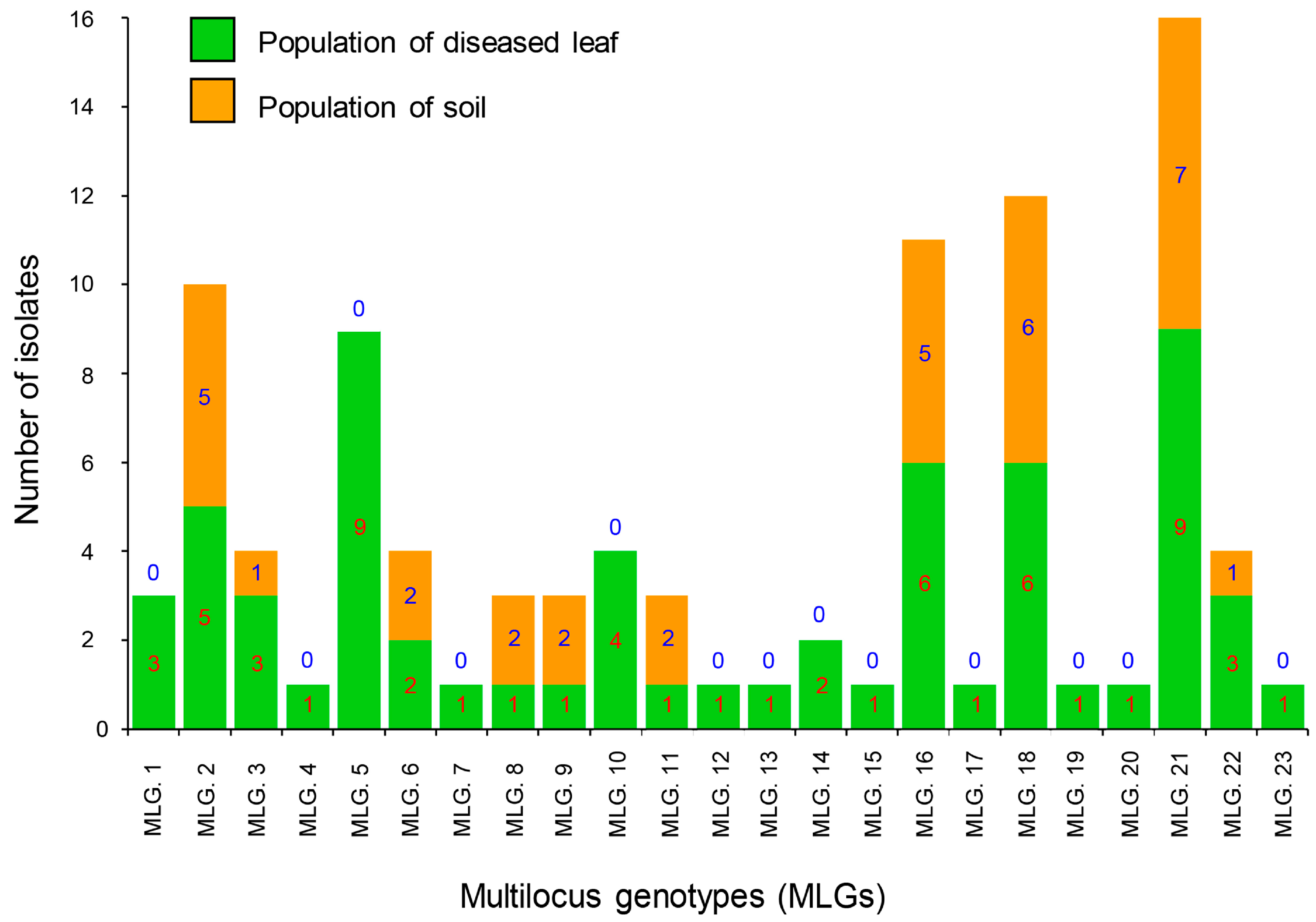
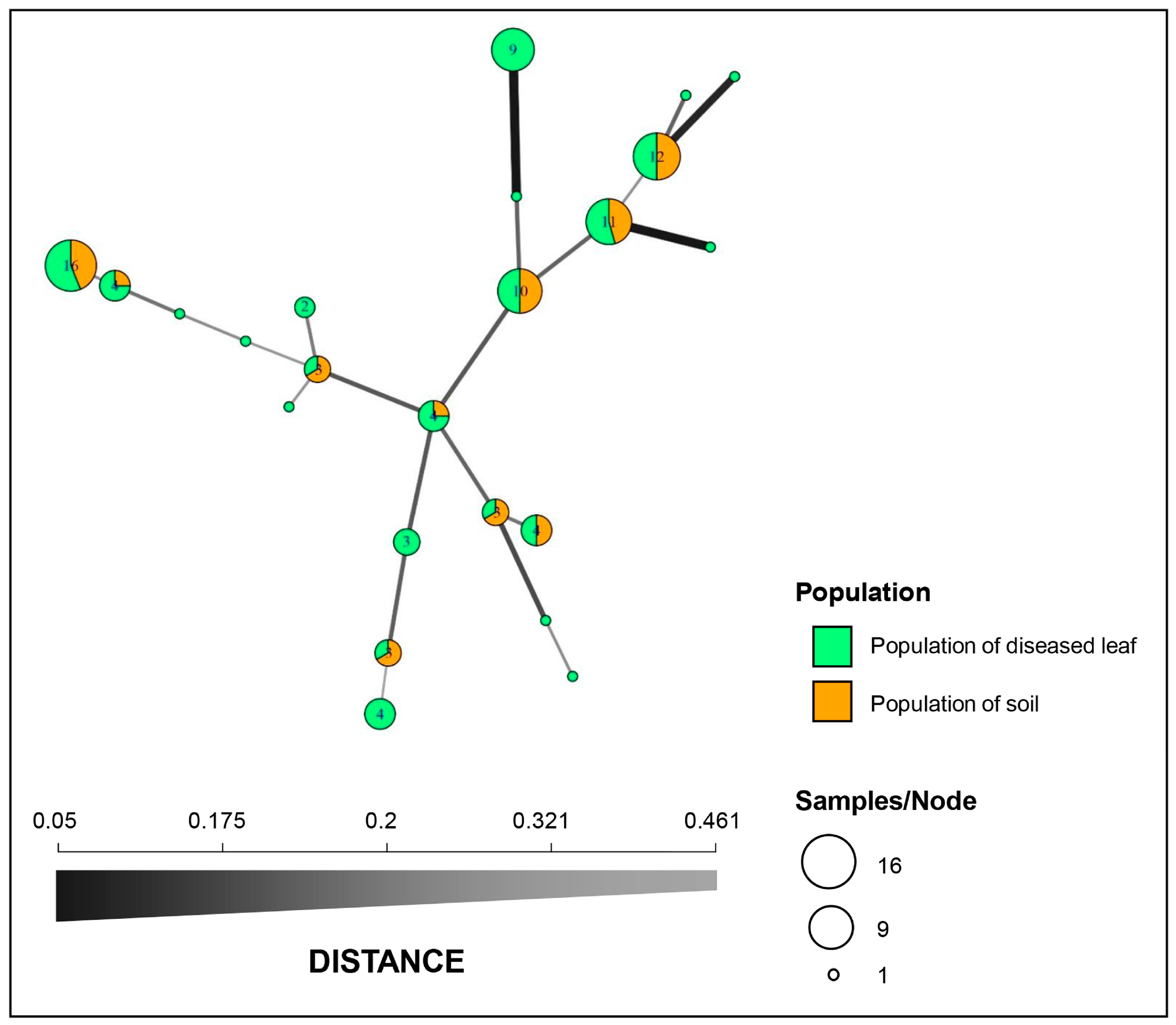
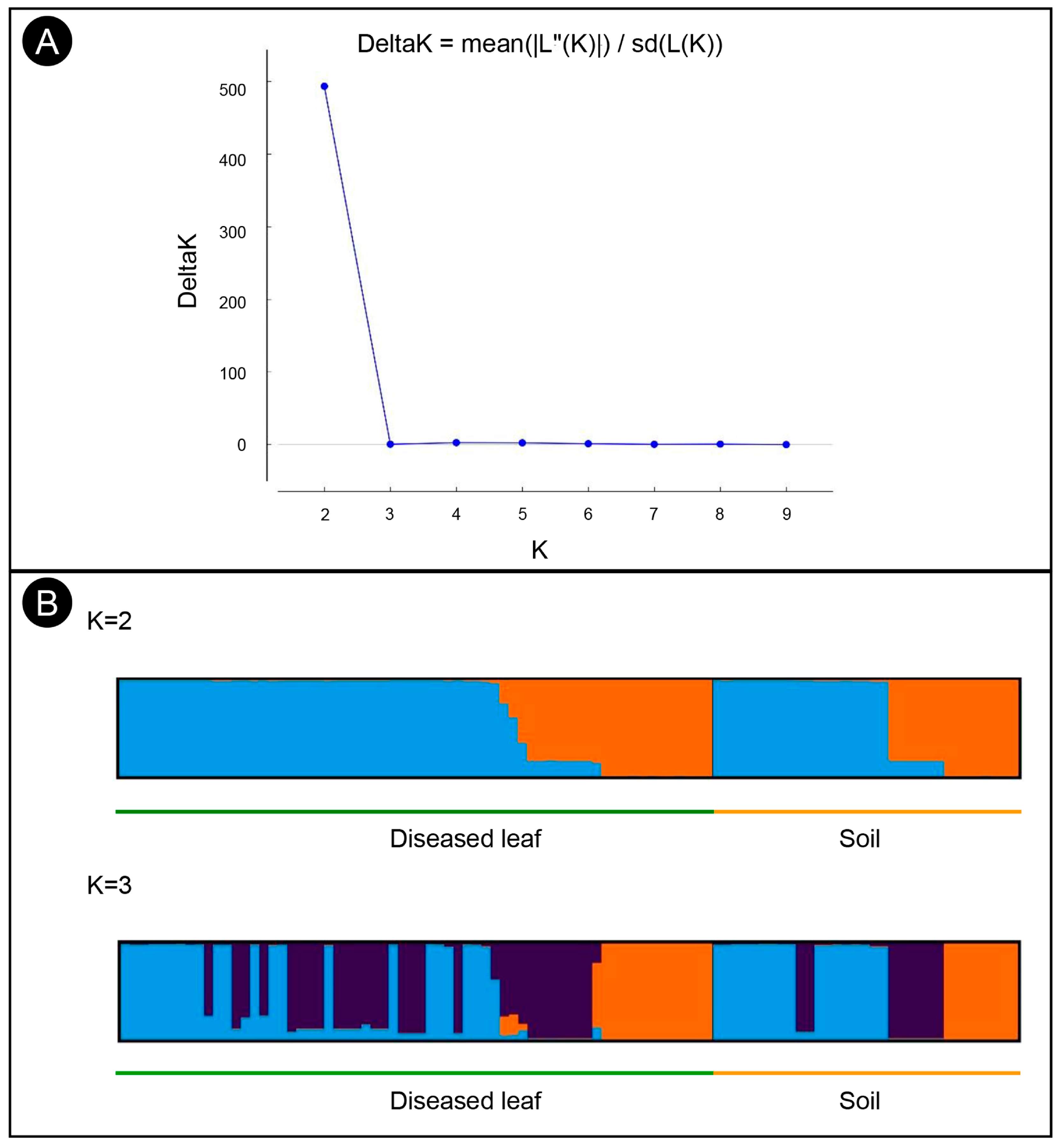
3.4. Population Structure, Minimum Spanning Network, and Molecular Variance Analyses
3.5. Population Reproduction Mode
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alfenas, R.F.; Lombard, L.; Pereira, O.L.; Alfenas, A.C.; Crous, P.W. Diversity and potential impact of Calonectria species in Eucalyptus plantations in Brazil. Stud. Mycol. 2015, 80, 89–130. [Google Scholar] [CrossRef]
- Bose, R.; Banerjee, S.; Pandey, A.; Bhandari, M.S.; Barthwal, S.; Pandey, S. Calonectria leaf blight of Eucalyptus: A global review. Ann. Appl. Biol. 2023, 182, 6–28. [Google Scholar] [CrossRef]
- Li, W.W.; Chen, S.F.; Wingfield, M.J.; Duong, T.A. Calonectria queenslandica: Causal agent of Eucalyptus leaf blight in Southern China. Plant Dis. 2023, 107, 730–742. [Google Scholar] [CrossRef]
- Li, W.W.; Chen, S.F.; Wingfield, M.J.; Duong, T.A. Calonectria species associated with diseased leaves and soils in southern China Eucalyptus plantations. Phytopathol. Res. 2023, 5, 29. [Google Scholar] [CrossRef]
- Liang, X.Y.; Wang, Q.C.; Chen, S.F. Phylogeny, morphology, distribution, and pathogenicity of seven Calonectria species from leaf blighted Eucalyptus in HaiNan Island, China. Plant Dis. 2023, 107, 2579–2605. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.L.; Li, J.Q.; Wingfield, M.J.; Duong, T.A.; Wingfield, B.D.; Crous, P.W.; Chen, S.F. Reconsideration of species boundaries and proposed DNA barcodes for Calonectria. Stud. Mycol. 2020, 97, 100106. [Google Scholar] [CrossRef] [PubMed]
- Pham, N.; Barnes, I.; Chen, S.F.; Liu, F.F.; Dang, Q.; Pham, T.; Lombard, L.; Crous, P.; Wingfield, M.J. Ten new species of Calonectria from Indonesia and Vietnam. Mycologia 2019, 111, 78–102. [Google Scholar] [CrossRef]
- Wang, Q.C.; Chen, S.F. Calonectria pentaseptata causes severe leaf disease on cultivated Eucalyptus in Leizhou Peninsula of southern China. Plant Dis. 2020, 104, 493–509. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.Z.; Guo, W.S.; Ye, X.Z.; Huang, X.P.; Wu, Y.Z. Identification of Calonectria associated with Eucalyptus leaf blight in Fujian Province. J. Fujian Coll. For. 2013, 33, 176–182. [Google Scholar]
- Wu, W.X.; Chen, S.F. Species diversity, mating strategy and pathogenicity of Calonectria species from diseased leaves and soils in the Eucalyptus plantation in Southern China. J. Fungi 2021, 7, 73. [Google Scholar] [CrossRef]
- Li, J.Q.; Wingfield, B.D.; Wingfield, M.J.; Barnes, I.; Fourie, A.; Crous, P.W.; Chen, S.F. Mating genes in Calonectria and evidence for a heterothallic ancestral state. Persoonia 2020, 45, 163–176. [Google Scholar] [CrossRef]
- Lombard, L.; Zhou, X.D.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Calonectria species associated with cutting rot of Eucalyptus. Persoonia 2010, 24, 1–11. [Google Scholar] [CrossRef]
- Tarigan, M.; Pham, N.Q.; Jami, F.; Oliveira, L.S.; Saha, M.A.; Durán, A.; Wingfield, M.J. Calonectria species diversity on eucalypts in Indonesia. South. For. J. For. Sci. 2023, 85, 56–64. [Google Scholar] [CrossRef]
- Li, J.Q.; Wingfield, M.J.; Liu, Q.L.; Barnes, I.; Roux, J.; Lombard, L.; Crous, P.W.; Chen, S.F. Calonectria species isolated from Eucalyptus plantations and nurseries in South China. IMA Fungus 2017, 8, 259–286. [Google Scholar] [CrossRef] [PubMed]
- Bose, R.; Banerjee, S.; Negi, N.; Pandey, A.; Bhandari, M.S.; Pandey, S. Identification and pathogenicity of Calonectria pseudoreteaudii causing leaf blight of Eucalyptus––a new record for India. Physiol. Mol. Plant Pathol. 2022, 122, 101917. [Google Scholar] [CrossRef]
- Chen, C.; Liang, X.; Lin, Y.; Hsiang, T.; Xiang, M.M.; Zhang, Y. First report of leaf spot and stem blight on blueberry (Vaccinium corymbosum ‘Bluerain’) caused by Calonectria pseudoreteaudii in China. Plant Dis. 2023, 7, 1951. [Google Scholar] [CrossRef]
- Crous, P.W.; Shivas, R.G.; Wingfield, M.J.; Summerell, B.A.; Rossman, A.Y.; Alves, J.L.; Adams, G.C.; Barreto, R.W.; Bell, A.; Coutinho, M.L.; et al. Fungal Planet description sheets: 128–153. Persoonia 2012, 29, 146–201. [Google Scholar] [CrossRef]
- Jiang, G.Z.; Gao, F.; Yue, H.; He, X.Y. First report of fruit spot of Macadamia sp. caused by Calonectria pentaseptata in China. Plant Dis. 2020, 104, 575. [Google Scholar] [CrossRef]
- Jiang, Z.E.; Xie, J.; Wei, J.G.; Luo, J.; Wu, Y.J.; Luo, J.T.; Yang, X.H.; Yang, X.B. First report of husk black spot on Macadamia ternifolia caused by Calonectria pentaseptata in China. Plant Dis. 2020, 104, 1551. [Google Scholar] [CrossRef]
- Phanthavong, S.; Daly, A.; Weir, B.; Lee, D.; Park, D.; Balmas, V.; Burgess, L. First report of Calonectria pseudoreteaudii in Lao PDR associated with a leaf spot disease of Macadamia integrifolia. Australas. Plant Pathol. 2023, 52, 23–26. [Google Scholar] [CrossRef]
- Lombard, L.; Chen, S.F.; Mou, X.; Zhou, X.D.; Crous, P.W.; Wingfield, M.J. New species, hyper-diversity and potential importance of Calonectria spp. from Eucalyptus in South China. Stud. Mycol. 2015, 80, 151–188. [Google Scholar] [CrossRef] [PubMed]
- van Burik, J.A.H.; Schreckhise, R.W.; White, T.C.; Bowden, R.A.; Myerson, D. Comparison of six extraction techniques for isolation of DNA from filamentous fungi. Med. Mycol. 1998, 36, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Li, W.W.; Liu, F.F.; Chen, S.F.; Wingfield, M.J.; Duong, T.A. High Genetic Diversity and Limited Regional Population Differentiation of Calonectria pseudoreteaudii Isolated from Diseased Eucalyptus Trees in Southern China; Research Institute of Fast-Growing Trees (RIFT): Zhanjiang, China; Chinese Academy of Forestry (CAF): Zhanjiang, China, 2023; manuscript in preparation. [Google Scholar]
- Kamvar, Z.N.; Tabima, J.F.; Grunwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [PubMed]
- Dowling, M.E.; Bryson, P.K.; Boatwright, H.G.; Wilson, J.R.; Fan, Z.; Everhart, S.E.; Brannen, P.M.; Schnabel, G. Effect of fungicide applications on Monilinia fructicola population diversity and transposon movement. Phytopathology 2016, 106, 1504–1512. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6, genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Nielsen, R.; Tarpy, D.R.; Reeve, H.K. Estimating effective paternity number in social insects and the effective number of alleles in a population. Mol. Ecol. 2003, 12, 3157–3164. [Google Scholar] [CrossRef]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. ACM SIGMOBILE Mob. Comput. Commun. Rev. 2001, 5, 3–55. [Google Scholar] [CrossRef]
- Stoddart, J.A.; Taylor, J.F. Genotypic diversity: Estimation and prediction in samples. Genetics 1988, 118, 705–711. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.; von Holdt, B. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. CLUMPAK: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 5, 1179–1191. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5, genetic analysis in Excel. Population genetic software for teaching and research–an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Agapow, P.M.; Burt, A. Indices of multilocus linkage disequilibrium. Mol. Ecol. Notes 2001, 1, 101102. [Google Scholar] [CrossRef]
- Grünwald, N.J.; Goodwin, S.B.; Milgroom, M.G.; Fry, W.E. Analysis of genotypic diversity data for populations of microorganisms. Phytopathology 2003, 93, 738–746. [Google Scholar] [CrossRef]
- Castroagudin, V.L.; Weiland, J.E.; Baysal-Gurel, F.; Cubeta, M.A.; Crouch, J.A. One clonal lineage of Calonectria pseudonaviculata is primarily responsible for the boxwood blight epidemic in the United States. Phytopathology 2020, 110, 1845–1853. [Google Scholar] [CrossRef]
- Freitas, R.G.; Alfenas, R.F.; Guimarães, L.M.S.; Badel, J.L.; Alfenas, A.C. Genetic diversity and aggressiveness of Calonectria pteridis in Eucalyptus spp. Plant Pathol. 2019, 68, 869–877. [Google Scholar] [CrossRef]
- LeBlanc, N.; Gehesquière, B.; Salgado-Salazar, C.; Heungens, K.; Crouch, J.A. Limited genetic diversity across pathogen populations responsible for the global emergence of boxwood blight identified using SSRs. Plant Pathol. 2019, 68, 861–868. [Google Scholar] [CrossRef]
- Li, J.Q.; Barnes, I.; Liu, F.F.; Wingfield, M.J.; Chen, S.F. Global genetic diversity and mating type distribution of Calonectria pauciramosa: An important wide-host-range plant pathogen. Plant Dis. 2021, 105, 1648–1656. [Google Scholar] [CrossRef]
- Wright, L.P.; Davis, A.J.; Wingfield, B.D.; Crous, P.W.; Brenneman, T.; Wingfield, M.J. Population structure of Cylindrocladium parasiticum infecting peanuts (Arachis hypogaea) in Georgia, USA. Eur. J. Plant Pathol. 2010, 127, 199–206. [Google Scholar] [CrossRef]
- Crous, P.W. Taxonomy and Pathology of Cylindrocladium (Calonectria) and Allied Genera; APS Press: St. Paul, MN, USA, 2002. [Google Scholar]
- McDonald, B.A.; McDermott, J.M. Population genetics of plant pathogenic fungi. Bioscience 1993, 43, 311–319. [Google Scholar] [CrossRef]
- McDonald, B.A.; Linde, C. Pathogen population genetics, evolutionary potential, and durable resistance. Annu. Rev. Phytopathol. 2002, 40, 349–379. [Google Scholar] [CrossRef] [PubMed]
- Alfenas, R.F.; Freitas, R.G.; Pereira, O.L.; Coutinho, M.M.; Zarpelon, T.G.; Cândido, T.S.; Alfenas, A.C. Screening of Corymbia and Eucalyptus species for resistance to Calonectria pteridis leaf blight. For. Pathol. 2016, 46, 76–81. [Google Scholar] [CrossRef]
- Rodas, C.A.; Lombard, L.; Gryzenhout, M.; Slipper, B.; Wingfield, M.J. Cylindrocladium blight of Eucalyptus grandis in Colombia. Australas. Plant Pathol. 2005, 34, 143–149. [Google Scholar] [CrossRef]
- Bai, Z.; Qin, Y.; Cao, K.; Du, J.; Han, Y.; Tan, Z.; Wu, G.; Tian, B.; Yang, Y.; Yu, Y.; et al. Genetic Diversity and Pathogenic Variation of the Rice False Smut Pathogen Ustilaginoidea virens from Different Rice Cultivars. Phytopathology 2023, 113, 549–558. [Google Scholar] [CrossRef]
- Wang, Q.C.; Chen, S.F. The resistances of eight Eucalyptus genotypes in southern China to Calonectria pentaseptata. Eucalypt Sci. Technol. 2020, 37, 1–9. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| SSR Locus | Allele Sizes (Isolate and GenBank Accession Numbers) | No. of Alleles | Hexp a | E.5 b |
|---|---|---|---|---|
| CPS103 | 283 c (CSF16053, OQ858289); 286 (CSF16063, OQ858290) | 2 | 0.48 | 0.96 |
| CPS108 | 258 (CSF16008, OQ858291); 273 (CSF16063, OQ858292); 279 (CSF16209, OQ858293); 282 (CSF16066, OQ858294); 285 (CSF16053, OQ858295) | 5 | 0.64 | 0.71 |
| CPS113 | 295 (CSF16066, OQ858296); 298 (CSF16083, OQ858297); 301 (CSF16053, OQ858298); 307 (CSF15996, OQ858299) | 4 | 0.65 | 0.94 |
| CPS115 | 381 (CSF15996, OQ865454); 384 (CSF16094, OQ865455); 387 (CSF16066, OQ865456); 390 (CSF16053, OQ865457); 393 (CSF16023, OQ865458) | 5 | 0.57 | 0.78 |
| CPS118 | 356 (CSF15984, OQ865459); 368 (CSF16101, OQ865460); 377 (CSF16063, OQ865461); 380 (CSF16053, OQ865462); 383 (CSF16068, OQ865463); 386 (CSF16066, OQ865464) | 6 | 0.79 | 0.92 |
| CPS140 | 349 (CSF15959, OQ865465); 355 (CSF16053, OQ865466); 358 (CSF15996, OQ865467); 364 (CSF16072, OQ865468); 385 (CSF15886, OQ865469) | 5 | 0.5 | 0.69 |
| CPS144 | 353 (CSF16009, OQ865470); 359 (CSF16080, OQ865471); 362 (CSF16063, OQ865472); 365 (CSF15865, OQ865473) | 4 | 0.55 | 0.77 |
| CPS156 | 298 (CSF16066, OQ865474); 302 (CSF15996, OQ865475); 306 (CSF15984, OQ865476); 314 (CSF15933, OQ865477); 318 (CSF16085, OQ865478); 326 (CSF16053, OQ865479); 334 (CSF15964, OQ865480); 342 (CSF16211, OQ865481); 346 (CSF16094, OQ865482); 406 (CSF15916, OQ865483); 418 (CSF15861, OQ865484) | 11 | 0.83 | 0.8 |
| CPS159 | 368 (CSF16072, OQ865485); 374 (CSF15886, OQ865486); 377 (CSF16053, OQ865487); 380 (CSF16209, OQ865488) | 4 | 0.51 | 0.66 |
| CPS161 | 435 (CSF15888, OQ865489); 443 (CSF16053, OQ865490); 447 (CSF16068, OQ865491); 451 (CSF15984, OQ865492) | 4 | 0.57 | 0.81 |
| Population | N a | Na b | Nef c | Hexp d | E e | MLG f | eMLG g | H h | G i |
|---|---|---|---|---|---|---|---|---|---|
| Diseased leaf | 64 | 50 | 3.090 | 0.636 | 0.800 | 23 | 16.310 | 2.795 | 12.721 |
| Soil | 33 | 34 | 2.535 | 0.581 | 0.840 | 10 | 10.000 | 2.102 | 7.118 |
| Source of Variation | Degrees of Freedom | Sum of Squares | Mean Squares | Estimate of Variance | Percentage of Total Variation (%) | PhiPT Value | p-Value a |
|---|---|---|---|---|---|---|---|
| Between populations | 1 | 4.537 | 4.537 | 0.033 | 1% | 0.011 | 0.177 |
| Within populations | 95 | 293.422 | 3.089 | 3.089 | 99% | – b | – |
| Total | 96 | 297.959 | N/A | 3.122 | 100% | – | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, W.; Li, W.; Liu, F.; Chen, S. Evidence of High Genetic Diversity and Differences in the Population Diversity of the Eucalyptus Leaf Blight Pathogen Calonectria pseudoreteaudii from Diseased Leaves and Soil in a Plantation in Guangxi, China. Microorganisms 2023, 11, 2785. https://doi.org/10.3390/microorganisms11112785
Wu W, Li W, Liu F, Chen S. Evidence of High Genetic Diversity and Differences in the Population Diversity of the Eucalyptus Leaf Blight Pathogen Calonectria pseudoreteaudii from Diseased Leaves and Soil in a Plantation in Guangxi, China. Microorganisms. 2023; 11(11):2785. https://doi.org/10.3390/microorganisms11112785
Chicago/Turabian StyleWu, Wenxia, Wenwen Li, Feifei Liu, and Shuaifei Chen. 2023. "Evidence of High Genetic Diversity and Differences in the Population Diversity of the Eucalyptus Leaf Blight Pathogen Calonectria pseudoreteaudii from Diseased Leaves and Soil in a Plantation in Guangxi, China" Microorganisms 11, no. 11: 2785. https://doi.org/10.3390/microorganisms11112785
APA StyleWu, W., Li, W., Liu, F., & Chen, S. (2023). Evidence of High Genetic Diversity and Differences in the Population Diversity of the Eucalyptus Leaf Blight Pathogen Calonectria pseudoreteaudii from Diseased Leaves and Soil in a Plantation in Guangxi, China. Microorganisms, 11(11), 2785. https://doi.org/10.3390/microorganisms11112785