
The Metabolic Potential of Endophytic Actinobacteria Associated with Medicinal Plant Thymus roseus as a Plant-Growth Stimulator

 ,
,  , ,
, ,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Screening of Potent Bacterial Endophytes for Plant Beneficial Traits
2.1.1. Indole-3-Acetic Acid (IAA) Production Assay
2.1.2. Phosphorus Solubilization Assay
2.1.3. Biological Nitrogen Fixation
2.1.4. Production of Siderophores by Endophytic Bacteria
2.1.5. Screening the Extracellular Enzymatic Activities
2.1.6. Screening for Antagonistic Activity
2.2. Effect of the Most Potent Endophytic Bacteria on Plant Growth Parameters
2.2.1. Greenhouse Experimental Design
2.2.2. Bacterial Inoculations and Soil Condition
2.2.3. Estimation of Plant Growth Parameters
2.3. Extraction and Identification of Metabolites
2.3.1. Isolation and Purification of Bioactive Compounds
2.3.2. Identification of Bioactive Compounds
2.4. Statistical Analysis
3. Results
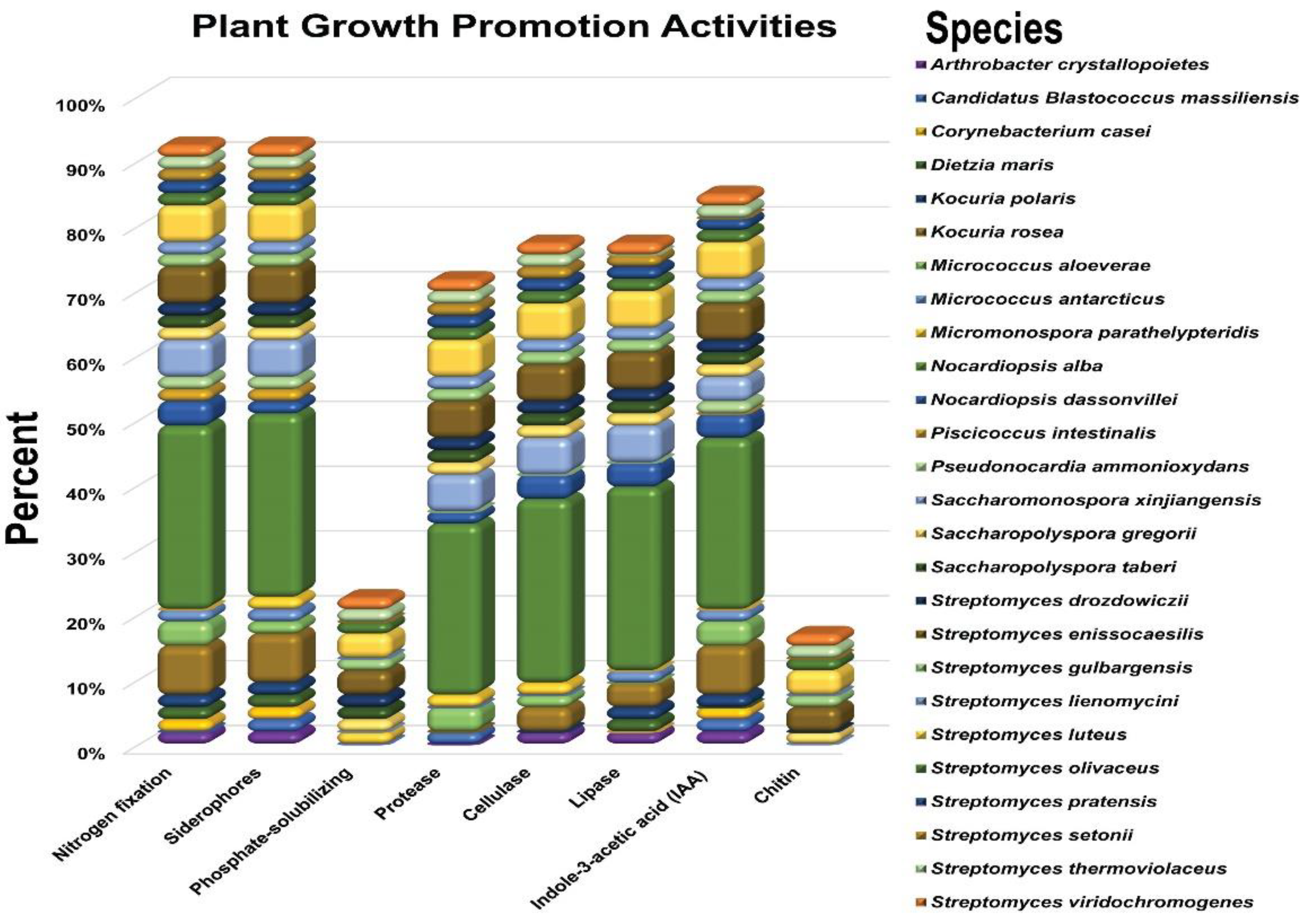
3.1. Plant Growth-Promoting (PGP) Parameters
3.1.1. Evaluation of PGP Traits
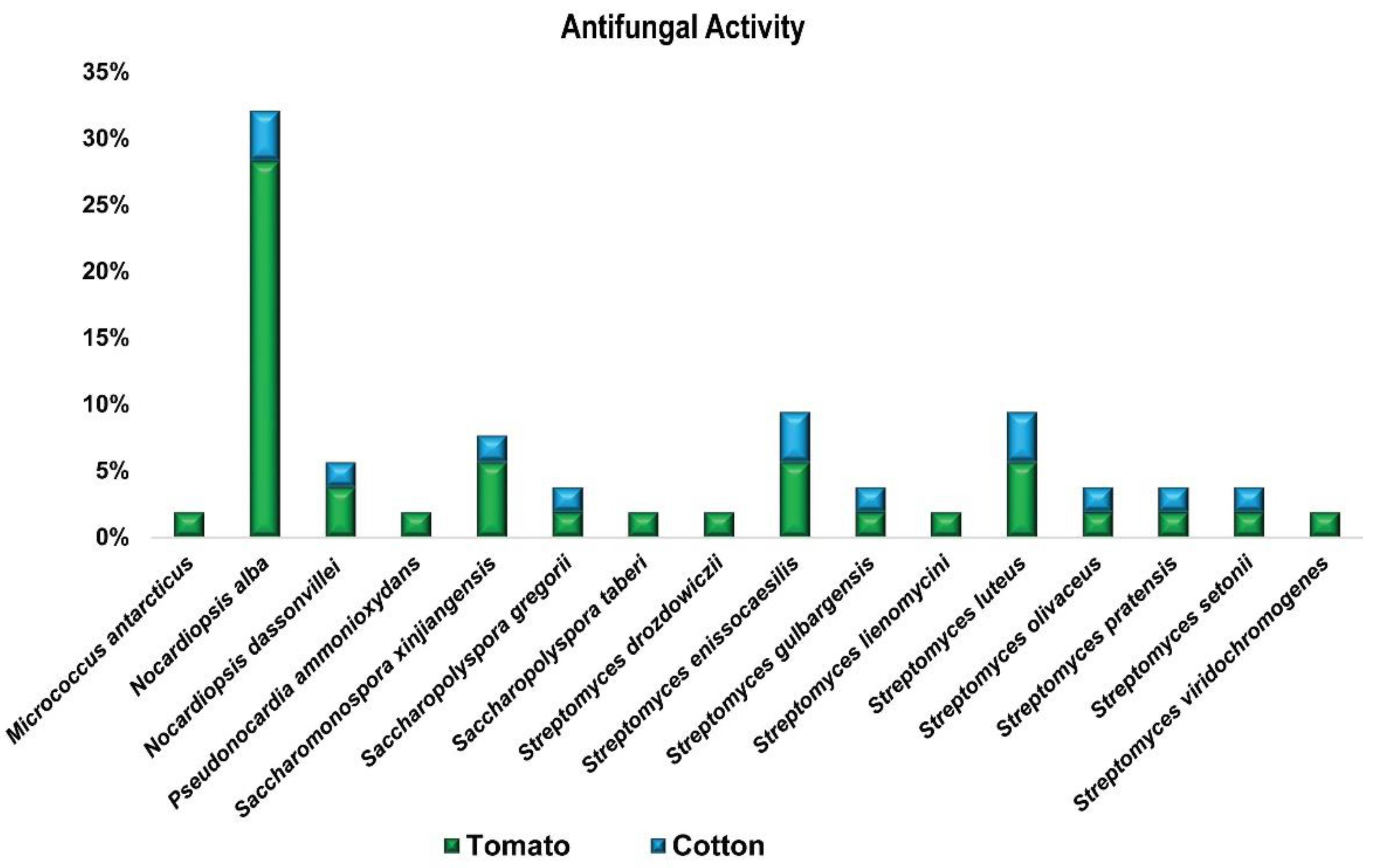
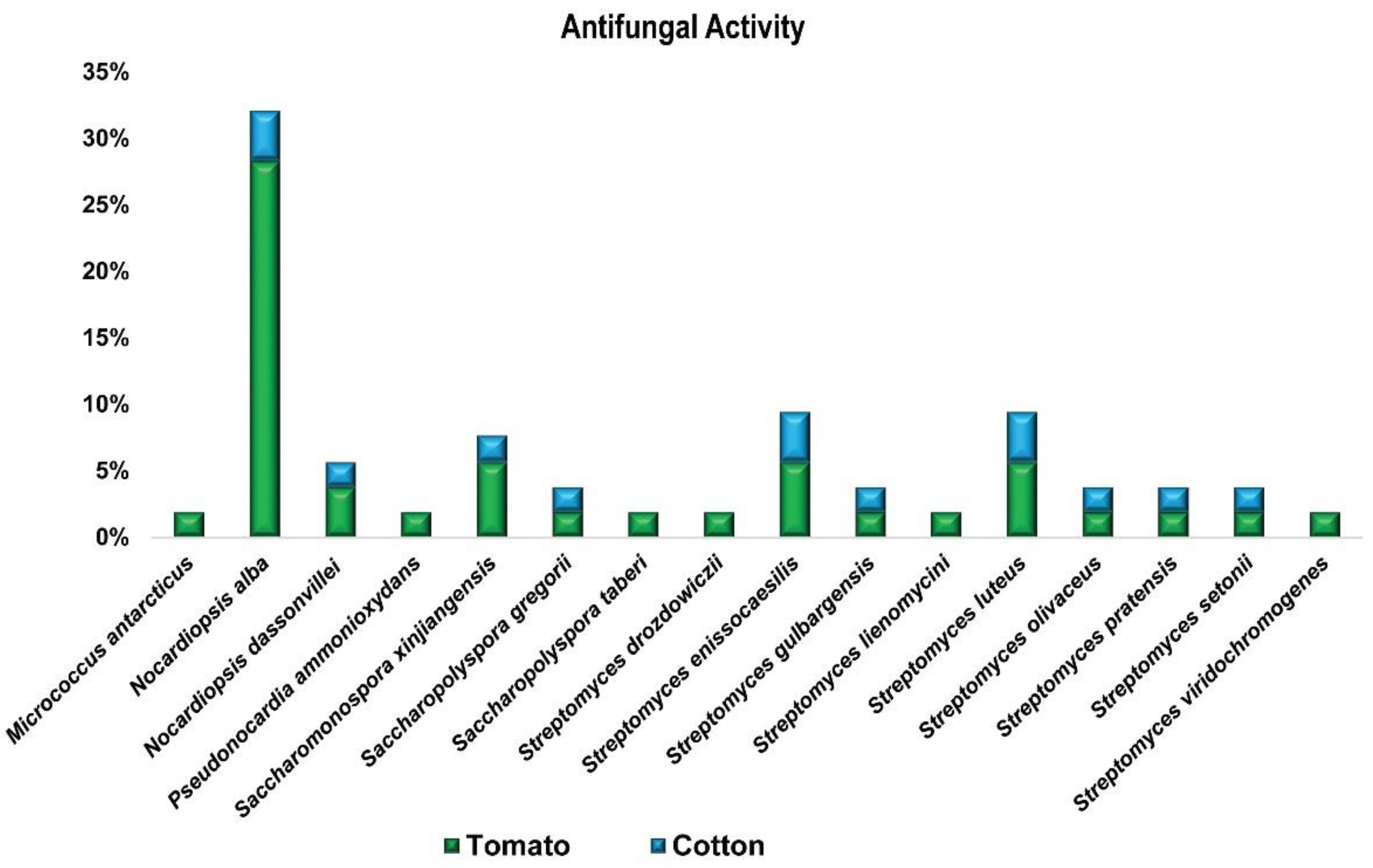
3.1.2. In-Vitro Antagonistic Bioassay
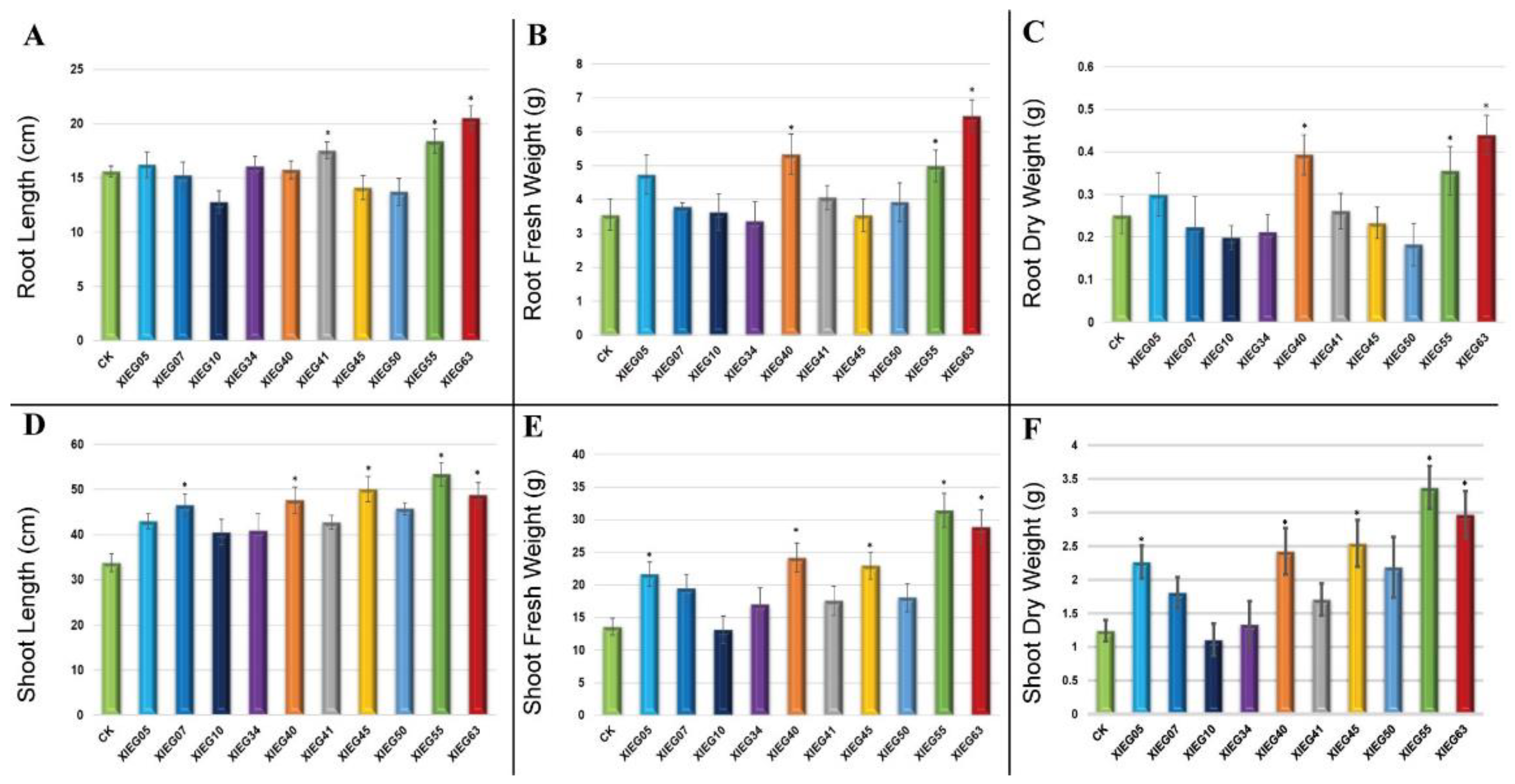
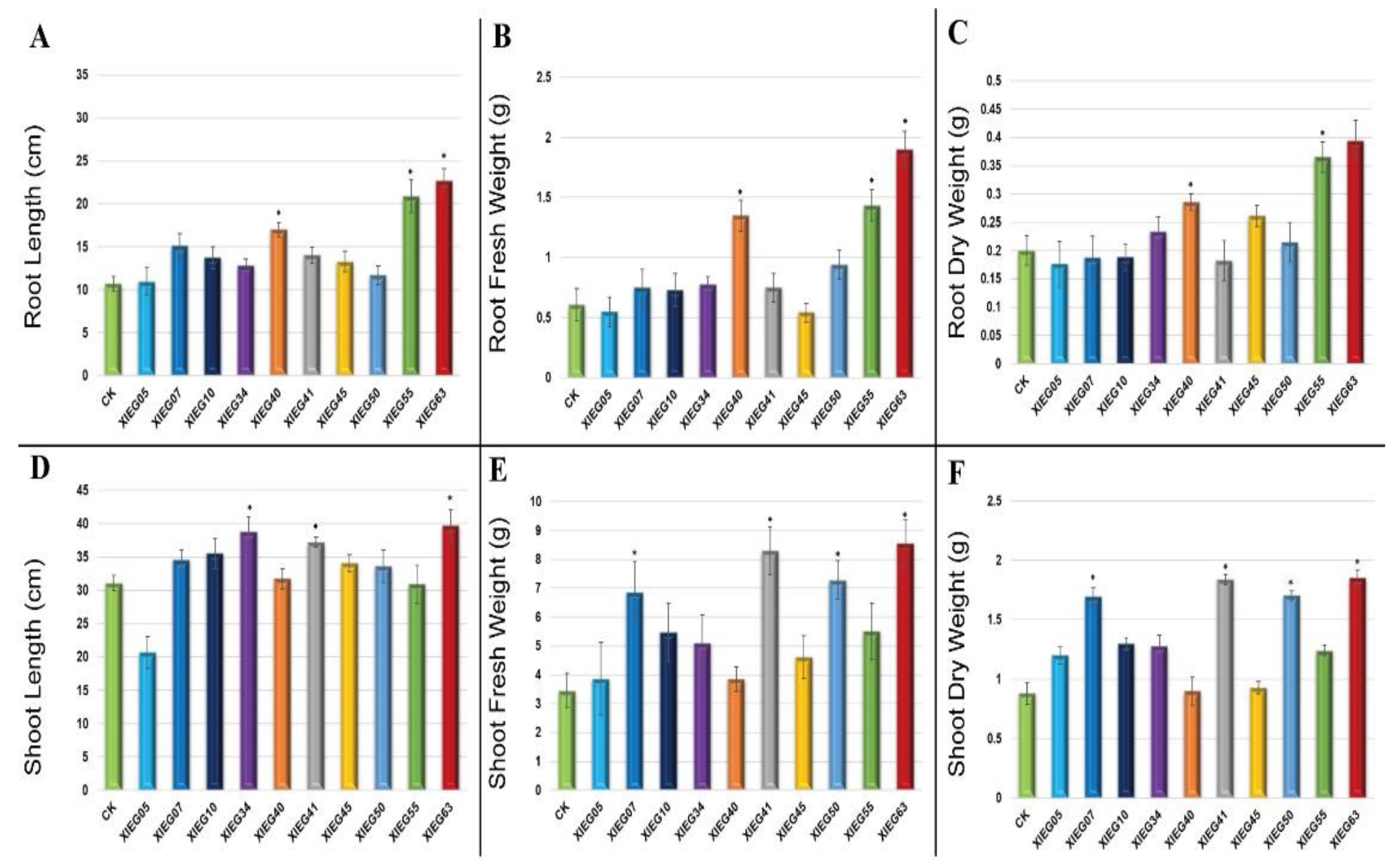
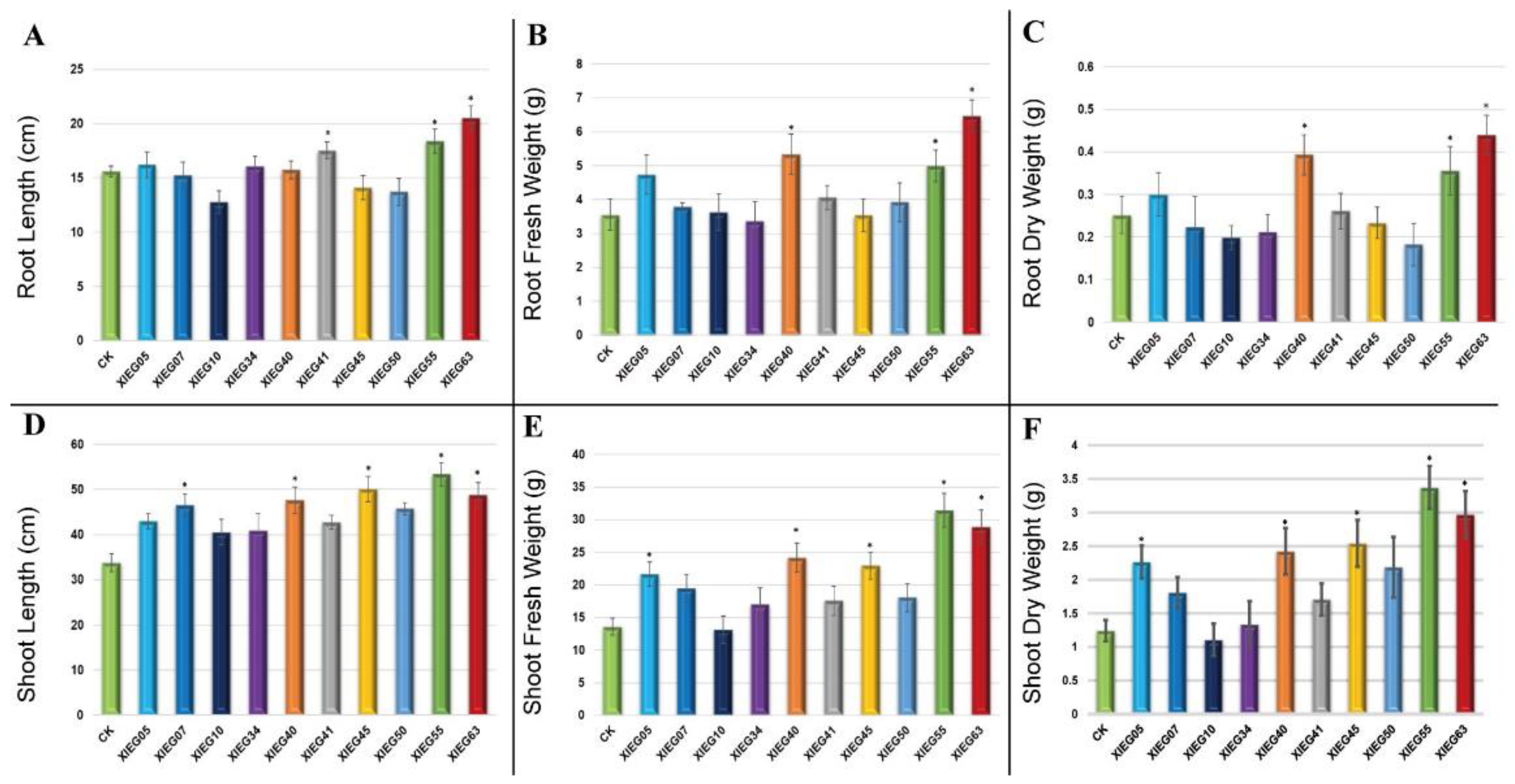
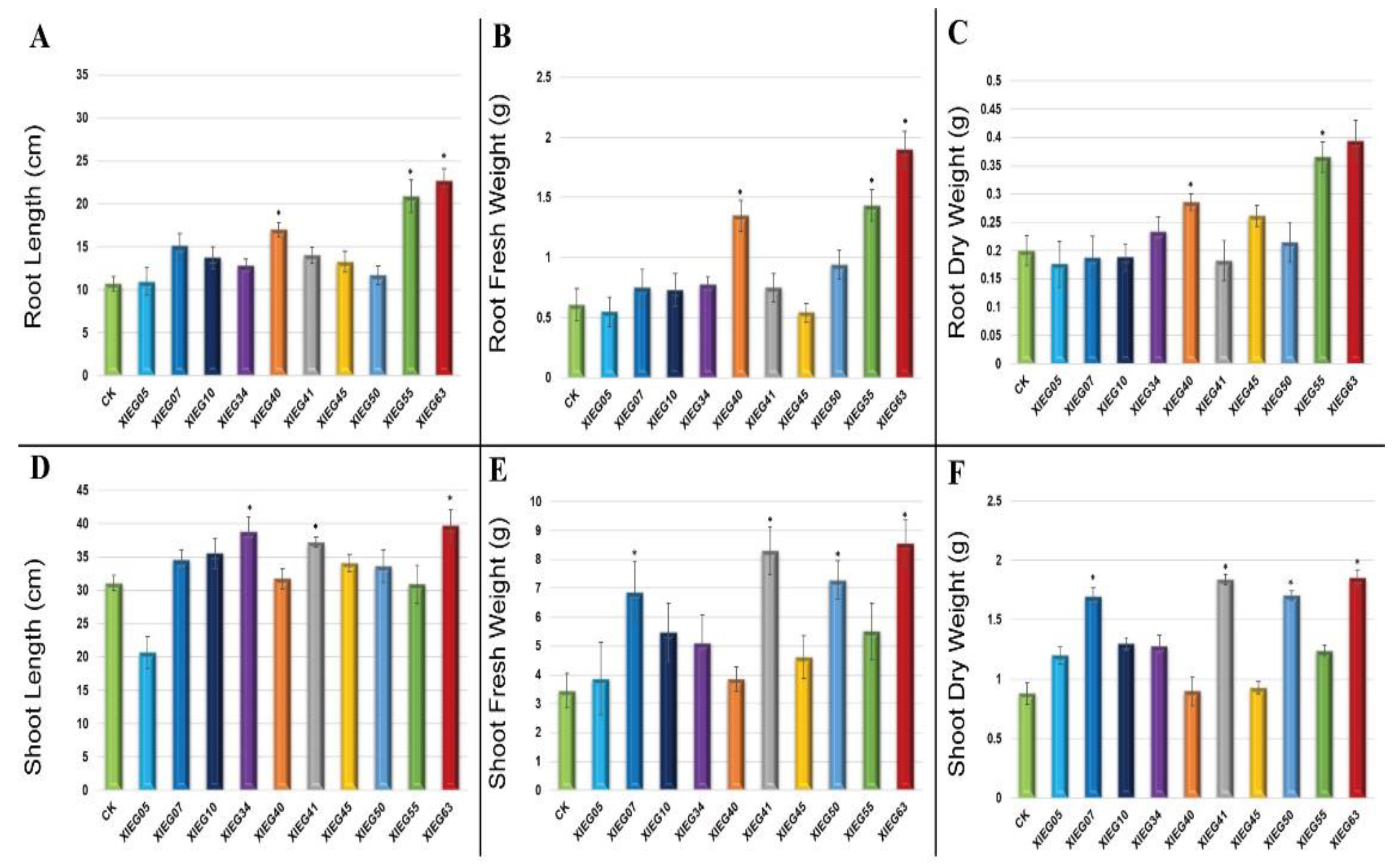
3.2. In-Planta Assay for Plant Growth Promotion Parameters by Selected Actinobacterial Strains
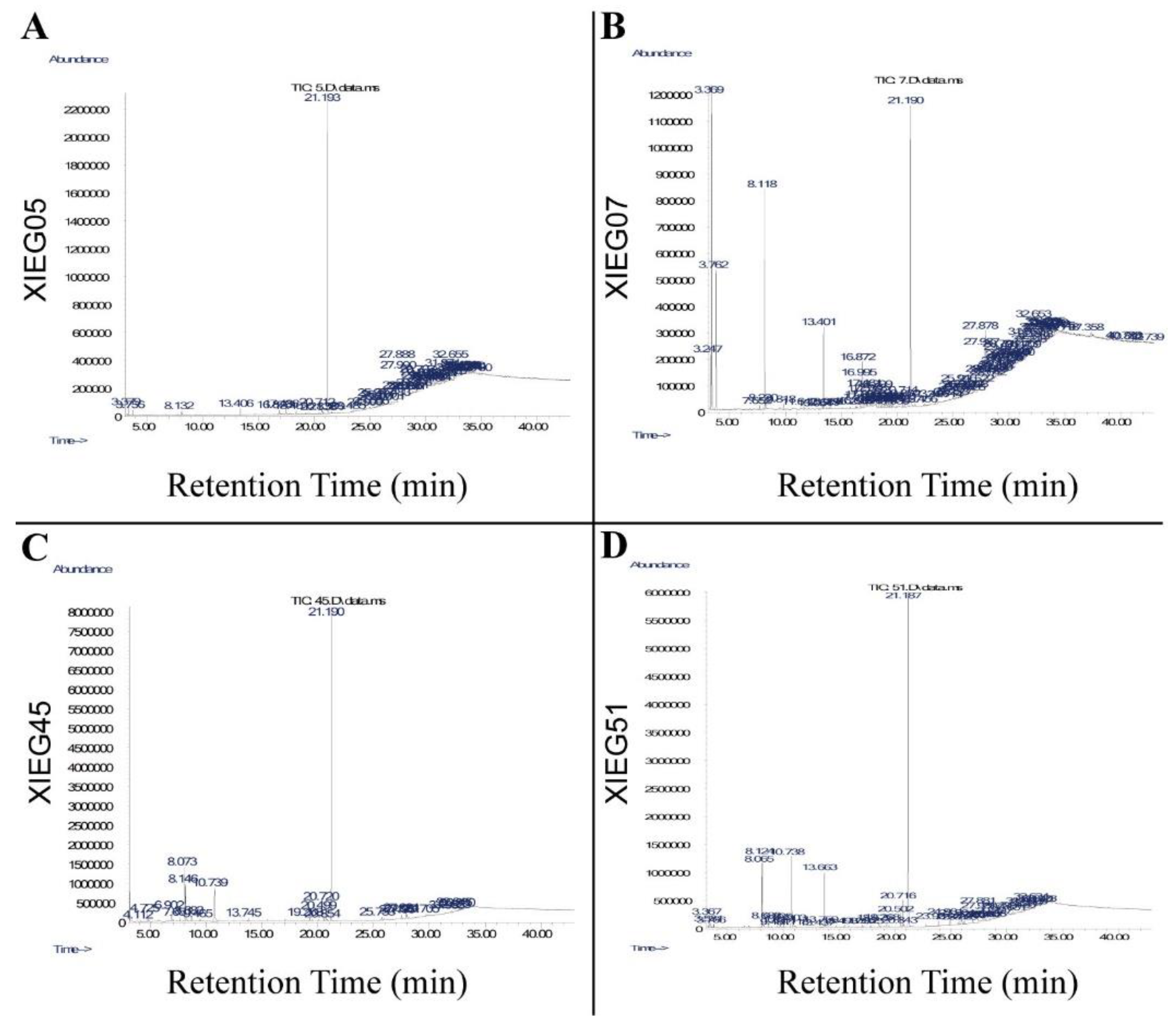
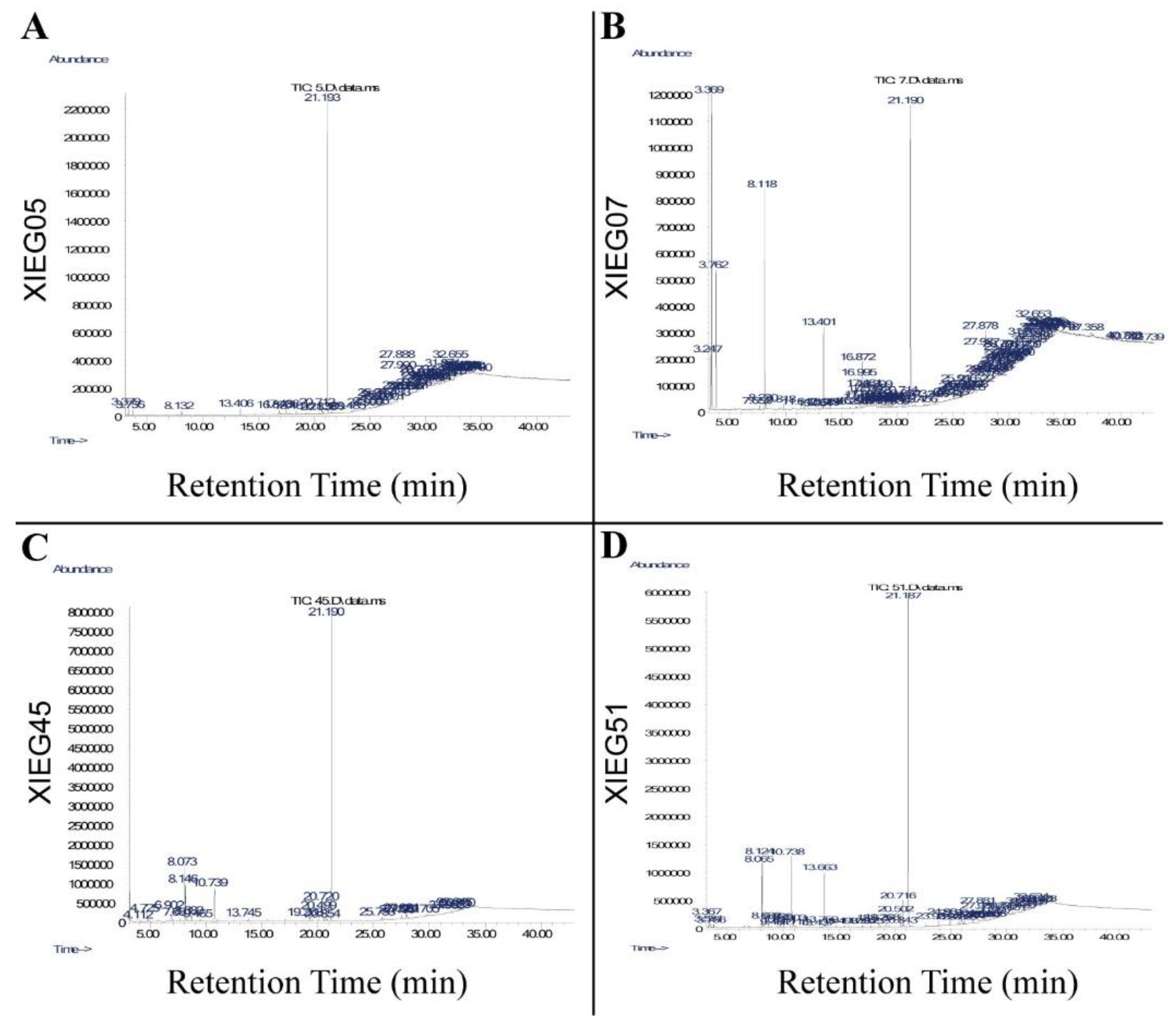
3.3. Detection of Bioactive Compounds by GC-MS Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- UN. World Population Prospects: The 2019 Revision; UN: New York, NY, USA, 2019. [Google Scholar]
- FAO. The Future of Food and Agriculture Alternative Pathways to 2050; FAO: Rome, Italy, 2018. [Google Scholar]
- FAO. The State of the World’s Land and Water Resources for Food and Agriculture: Managing Systems at Risk; FAO: Rome, Italy, 2021. [Google Scholar]
- Chang, Q.; Wang, W.; Regev-Yochay, G.; Lipsitch, M.; Hanage, W.P. Antibiotics in agriculture and the risk to human health: How worried should we be? Evol. Appl. 2015, 8, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yan, C.; Guo, Q.; Zhang, J.; Ruiz-Menjivar, J. The impact of agricultural chemical inputs on environment: Global evidence from informetrics analysis and visualization. Int. J. Low-Carbon Technol. 2018, 13, 338–352. [Google Scholar] [CrossRef]
- Kopittke, P.M.; Menzies, N.W.; Wang, P.; McKenna, B.A.; Lombi, E. Soil and the intensification of agriculture for global food security. Environ. Int. 2019, 132, 105078. [Google Scholar] [CrossRef]
- Mueller, U.G.; Sachs, J.L. Engineering microbiomes to improve plant and animal health. Trends Microbiol. 2015, 23, 606–617. [Google Scholar] [CrossRef]
- Scagliola, M.; Valentinuzzi, F.; Mimmo, T.; Cesco, S.; Crecchio, C.; Pii, Y. Bioinoculants as promising complement of chemical fertilizers for a more sustainable agricultural practice. Front. Sustain. Food Syst. 2021, 4, 305. [Google Scholar] [CrossRef]
- Saharan, B.; Nehra, V. Plant growth promoting rhizobacteria: A critical review. Life Sci. Med. Res. 2011, 21, 30. [Google Scholar]
- Fasusi, O.A.; Cruz, C.; Babalola, O.O. Agricultural Sustainability: Microbial Biofertilizers in Rhizosphere Management. Agriculture 2021, 11, 163. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, W.; Lang, D.; Zhang, X.; Cui, G.; Zhang, X. Interactions between endophytes and plants: Beneficial effect of endophytes to ameliorate biotic and abiotic stresses in plants. J. Plant Biol. 2019, 62, 1–13. [Google Scholar]
- Kushwaha, P.; Kashyap, P.L.; Bhardwaj, A.K.; Kuppusamy, P.; Srivastava, A.K.; Tiwari, R.K. Bacterial endophyte mediated plant tolerance to salinity: Growth responses and mechanisms of action. World J. Microbiol. Biotechnol. 2020, 36, 26. [Google Scholar] [CrossRef]
- Kushwaha, P.; Kashyap, P.L.; Srivastava, A.K.; Tiwari, R.K. Plant growth promoting and antifungal activity in endophytic Bacillus strains from pearl millet (Pennisetum glaucum). Braz. J. Microbiol. 2020, 51, 229–241. [Google Scholar] [CrossRef]
- Galletti, S.; Paris, R.; Cianchetta, S. Selected isolates of Trichoderma gamsii induce different pathways of systemic resistance in maize upon Fusarium verticillioides challenge. Microbiol. Res. 2020, 233, 126406. [Google Scholar] [CrossRef]
- Borah, A.; Thakur, D. Phylogenetic and functional characterization of culturable endophytic actinobacteria associated with Camellia spp. for growth promotion in commercial tea cultivars. Front. Microbiol. 2020, 11, 318. [Google Scholar] [CrossRef] [Green Version]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Kashyap, P.L.; Solanki, M.K.; Kushwaha, P.; Kumar, S.; Srivastava, A.K. Biocontrol Potential of Salt-Tolerant Trichoderma and Hypocrea Isolates for the Management of Tomato Root Rot Under Saline Environment. J. Soil Sci. Plant Nutr. 2019, 20, 160–176. [Google Scholar] [CrossRef]
- Rana, K.L.; Kour, D.; Kaur, T.; Devi, R.; Yadav, A.; Yadav, A.N. Bioprospecting of endophytic bacteria from the Indian Himalayas and their role in plant growth promotion of maize (Zea mays L.). J. Appl. Biol. Biotechnol. 2021, 9, 41–50. [Google Scholar]
- Bona, E.; Lingua, G.; Manassero, P.; Cantamessa, S.; Marsano, F.; Todeschini, V.; Copetta, A.; D’Agostino, G.; Massa, N.; Avidano, L.; et al. AM fungi and PGP pseudomonads increase flowering, fruit production, and vitamin content in strawberry grown at low nitrogen and phosphorus levels. Mycorrhiza 2015, 25, 181–193. [Google Scholar] [CrossRef]
- Pii, Y.; Graf, H.; Valentinuzzi, F.; Cesco, S.; Mimmo, T. The effects of plant growth-promoting rhizobacteria (PGPR) on the growth and quality of strawberries. Acta Hortic. 2018, 1217, 231–238. [Google Scholar] [CrossRef]
- Kumar, A.; Vandana; Singh, M.; Singh, P.P.; Singh, S.K.; Singh, P.K.; Pandey, K.D. Isolation of plant growth promoting rhizobacteria and their impact on growth and curcumin content in Curcuma longa L. Biocatal. Agric. Biotechnol. 2016, 8, 1–7. [Google Scholar] [CrossRef]
- Saeed, K.S.; Ahmed, S.A.; Hassan, I.A.; Ahmed, P.J. Effect of bio-fertilizer and chemical fertilizer on growth and yield in cucumber (Cucumis sativus L.) in green house condition. Pak. J. Biol. Sci. 2015, 18, 129–134. [Google Scholar]
- Arif, M.S.; Riaz, M.; Shahzad, S.M.; Yasmeen, T.; Akhtar, M.J.; Riaz, M.A.; Jassey, V.E.J.; Bragazza, L.; Buttler, A. Associative interplay of plant growth promoting rhizobacteria (Pseudomonas aeruginosa QS40) with nitrogen fertilizers improves sunflower (Helianthus annuus L.) productivity and fertility of aridisol. Appl. Soil Ecol. 2016, 108, 238–247. [Google Scholar] [CrossRef]
- Patel, J.K.; Madaan, S.; Archana, G. Antibiotic producing endophytic Streptomyces spp. colonize above-ground plant parts and promote shoot growth in multiple healthy and pathogen-challenged cereal crops. Microbiol. Res. 2018, 215, 36–45. [Google Scholar] [CrossRef]
- Vijayabharathi, R.; Gopalakrishnan, S.; Sathya, A.; Srinivas, V.; Sharma, M. Deciphering the tri-dimensional effect of endophytic Streptomyces sp. on chickpea for plant growth promotion, helper effect with Mesorhizobium ciceri and host-plant resistance induction against Botrytis cinerea. Microb. Pathog. 2018, 122, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Selim, M.S.M.; Abdelhamid, S.A.; Mohamed, S.S. Secondary metabolites and biodiversity of actinomycetes. J. Genet. Eng. Biotechnol. 2021, 19, 72. [Google Scholar] [CrossRef]
- Abdelshafy Mohamad, O.A.; Li, L.; Ma, J.-B.; Hatab, S.; Rasulov, B.A.; Musa, Z.; Liu, Y.-H.; Li, W.-J. Halophilic Actinobacteria Biological Activity and Potential Applications. In Extremophiles in Eurasian Ecosystems: Ecology, Diversity, and Applications; Egamberdieva, D., Birkeland, N.-K., Panosyan, H., Li, W.-J., Eds.; Springer Singapore: Singapore, 2018; pp. 333–364. [Google Scholar]
- Nafis, A.; Raklami, A.; Bechtaoui, N.; El Khalloufi, F.; El Alaoui, A.; Glick, B.R.; Hafidi, M.; Kouisni, L.; Ouhdouch, Y.; Hassani, L. Actinobacteria from Extreme Niches in Morocco and Their Plant Growth-Promoting Potentials. Diversity 2019, 11, 139. [Google Scholar] [CrossRef] [Green Version]
- Narsing Rao, M.P.; Li, W.-J. Diversity of Actinobacteria in Various Habitats. In Actinobacteria; Springer: Berlin/Heidelberg, Germany, 2022; pp. 37–58. [Google Scholar]
- Singh, N.A.; Jain, R. Diversity and Bioactive Potential of Endophytic Bacteria from High-Value Medicinal Plants. In Bacterial Endophytes for Sustainable Agriculture and Environmental Management; Singh, A.K., Tripathi, V., Shukla, A.K., Kumar, P., Eds.; Springer: Singapore, 2022; pp. 45–69. [Google Scholar]
- Singh, R.; Dubey, A.K. Diversity and Applications of Endophytic Actinobacteria of Plants in Special and Other Ecological Niches. Front. Microbiol. 2018, 9, 1767. [Google Scholar] [CrossRef] [PubMed]
- Sathya, A.; Vijayabharathi, R.; Gopalakrishnan, S. Plant growth-promoting actinobacteria: A new strategy for enhancing sustainable production and protection of grain legumes. 3 Biotech 2017, 7, 102. [Google Scholar] [CrossRef]
- Golinska, P.; Wypij, M.; Agarkar, G.; Rathod, D.; Dahm, H.; Rai, M. Endophytic actinobacteria of medicinal plants: Diversity and bioactivity. Antonie van Leeuwenhoek 2015, 108, 267–289. [Google Scholar] [CrossRef]
- Kamboj, V.P. Herbal medicine. Curr. Sci. 2000, 78, 35–39. [Google Scholar]
- Saad, M.M.; Eida, A.A.; Hirt, H. Tailoring plant-associated microbial inoculants in agriculture: A roadmap for successful application. J. Exp. Bot. 2020, 71, 3878–3901. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, A.A.; Upadhyay, S.K.; Srivastava, A.K.; Bhojiya, A.A. Biofertilizers: A Nexus between soil fertility and crop productivity under abiotic stress. Curr. Res. Environ. Sustain. 2021, 3, 100063. [Google Scholar] [CrossRef]
- Batista, B.D.; Singh, B.K. Realities and hopes in the application of microbial tools in agriculture. Microb. Biotechnol. 2021, 14, 1258–1268. [Google Scholar] [CrossRef] [PubMed]
- Koskey, G.; Mburu, S.W.; Awino, R.; Njeru, E.M.; Maingi, J.M. Potential Use of Beneficial Microorganisms for Soil Amelioration, Phytopathogen Biocontrol, and Sustainable Crop Production in Smallholder Agroecosystems. Front. Sustain. Food Syst. 2021, 5, 130. [Google Scholar] [CrossRef]
- Hammoudi Halat, D.; Krayem, M.; Khaled, S.; Younes, S. A Focused Insight into Thyme: Biological, Chemical, and Therapeutic Properties of an Indigenous Mediterranean Herb. Nutrients 2022, 14, 2104. [Google Scholar] [CrossRef] [PubMed]
- Nieto, G. A Review on Applications and Uses of Thymus in the Food Industry. Plants 2020, 9, 961. [Google Scholar] [CrossRef]
- McGovern, R.J. Management of tomato diseases caused by Fusarium oxysporum. Crop Prot. 2015, 73, 78–92. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, X.; Zhang, C.; Zhang, Q.; Sun, Y.; Zhu, H.; Tang, C. Pectin lyase enhances cotton resistance to Verticillium wilt by inducing cell apoptosis of Verticillium dahliae. J. Hazard. Mater. 2021, 404, 124029. [Google Scholar] [CrossRef] [PubMed]
- Musa, Z.; Ma, J.; Egamberdieva, D.; Abdelshafy Mohamad, O.A.; Abaydulla, G.; Liu, Y.; Li, W.-J.; Li, L. Diversity and antimicrobial potential of cultivable endophytic actinobacteria associated with the medicinal plant Thymus roseus. Front. Microbiol. 2020, 11, 191. [Google Scholar] [CrossRef]
- Li, L.; Mohamad, O.A.A.; Ma, J.B.; Friel, A.D.; Su, Y.G.; Wang, Y.; Musa, Z.; Liu, Y.H.; Hedlund, B.P.; Li, W.J. Synergistic plant-microbe interactions between endophytic bacterial communities and the medicinal plant Glycyrrhiza uralensis F. Anton Leeuw Int. J G 2018, 111, 1735–1748. [Google Scholar] [CrossRef] [PubMed]
- Abdelshafy Mohamad, O.A.; Ma, J.-B.; Liu, Y.-H.; Zhang, D.; Hua, S.; Bhute, S.; Hedlund, B.P.; Li, W.-J.; Li, L. Beneficial Endophytic Bacterial Populations Associated With Medicinal Plant Thymus vulgaris Alleviate Salt Stress and Confer Resistance to Fusarium oxysporum. Front. Plant Sci. 2020, 11, 47. [Google Scholar] [CrossRef] [PubMed]
- da Cunha Ferreira, S.; Nakasone, A.K.; do Nascimento, S.M.C.; de Oliveira, D.A.; Siqueira, A.S.; Cunha, E.F.M.; de Castro, G.L.S.; de Souza, C.R.B. Isolation and characterization of cassava root endophytic bacteria with the ability to promote plant growth and control the in vitro and in vivo growth of Phytopythium sp. Physiol. Mol. Plant Pathol. 2021, 116, 101709. [Google Scholar] [CrossRef]
- Ismail, M.A.; Amin, M.A.; Eid, A.M.; Hassan, S.E.-D.; Mahgoub, H.A.; Lashin, I.; Abdelwahab, A.T.; Azab, E.; Gobouri, A.A.; Elkelish, A.J.C. Comparative Study between exogenously applied plant growth hormones versus metabolites of microbial endophytes as plant growth-promoting for Phaseolus vulgaris L. Cells 2021, 10, 1059. [Google Scholar] [CrossRef]
- Dubey, A.; Saiyam, D.; Kumar, A.; Hashem, A.; Abd_Allah, E.F.; Khan, M.L. Bacterial root endophytes: Characterization of their competence and plant growth promotion in soybean (Glycine max (L.) Merr.) under drought stress. Int. J. Environ. Res. Public Health 2021, 18, 931. [Google Scholar] [CrossRef]
- Fouda, A.; Eid, A.M.; Elsaied, A.; El-Belely, E.F.; Barghoth, M.G.; Azab, E.; Gobouri, A.A.; Hassan, S.E.-D. Plant growth-promoting endophytic bacterial community inhabiting the leaves of Pulicaria incisa (Lam.) DC inherent to arid regions. Plants 2021, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, T.; Kotasthane, A.S. Chitinolytic assay of indigenous Trichoderma isolates collected from different geographical locations of Chhattisgarh in Central India. SpringerPlus 2012, 1, 73. [Google Scholar] [CrossRef]
- Mohamad, O.A.; Li, L.; Ma, J.-B.; Hatab, S.; Xu, L.; Guo, J.-W.; Rasulov, B.A.; Liu, Y.-H.; Hedlund, B.P.; Li, W.-J. Evaluation of the antimicrobial activity of endophytic bacterial populations from Chinese traditional medicinal plant licorice and characterization of the bioactive secondary metabolites produced by Bacillus atrophaeus against Verticillium dahliae. Front. Microbiol. 2018, 9, 924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahgoub, H.A.; Fouda, A.; Eid, A.M.; Ewais, E.E.-D.; Hassan, S.E.-D. Biotechnological application of plant growth-promoting endophytic bacteria isolated from halophytic plants to ameliorate salinity tolerance of Vicia faba L. Plant Biotechnol. Rep. 2021, 15, 819–843. [Google Scholar] [CrossRef]
- Mohamad, O.A.A.; Ma, J.-B.; Liu, Y.-H.; Li, L.; Hatab, S.; Li, W.-J. Medicinal plant-associated microbes as a source of protection and production of crops. In Medically Important Plant Biomes: Source of Secondary Metabolites; Springer: Berlin/Heidelberg, Germany, 2019; pp. 239–263. [Google Scholar]
- Dede, A.; Güven, K. Plant growth-promoting of olive and walnut actinobacteria: Isolation, screening PGP traits, antifungal activities, identification, and hydroponic production of wheat. Arch. Agron. Soil Sci. 2022. [Google Scholar] [CrossRef]
- Alblooshi, A.A.; Purayil, G.P.; Saeed, E.E.; Ramadan, G.A.; Tariq, S.; Altaee, A.S.; El-Tarabily, K.A.; AbuQamar, S.F. Biocontrol Potential of Endophytic Actinobacteria against Fusarium solani, the Causal Agent of Sudden Decline Syndrome on Date Palm in the UAE. J. Fungi 2022, 8, 8. [Google Scholar] [CrossRef]
- de Jesus Sousa, J.A.; Olivares, F.L. Plant growth promotion by streptomycetes: Ecophysiology, mechanisms and applications. Chem. Biol. Technol. Agric. 2016, 3, 1–12. [Google Scholar]
- Zhao, L.; Xu, Y.; Lai, X.-H.; Shan, C.; Deng, Z.; Ji, Y. Screening and characterization of endophytic Bacillus and Paenibacillus strains from medicinal plant Lonicera japonica for use as potential plant growth promoters. Braz. J. Microbiol. 2015, 46, 977–989. [Google Scholar] [CrossRef]
- Maldonado, S.; Rodríguez, A.; Ávila, B.; Morales, P.; González, M.P.; Araya Angel, J.P.A.; Olalde, V.; Bravo, J.; Jana, C.; Sierra, C.; et al. Enhanced Crop Productivity and Sustainability by Using Native Phosphate Solubilizing Rhizobacteria in the Agriculture of Arid Zones. Front. Sustain. Food Syst. 2020, 4, 607355. [Google Scholar] [CrossRef]
- Pandey, A.; Trivedi, P.; Kumar, B.; Palni, L.M.S. Characterization of a phosphate solubilizing and antagonistic strain of Pseudomonas putida (B0) isolated from a sub-alpine location in the Indian Central Himalaya. Curr. Microbiol. 2006, 53, 102–107. [Google Scholar] [CrossRef]
- Goswami, D.; Dhandhukia, P.; Patel, P.; Thakker, J.N. Screening of PGPR from saline desert of Kutch: Growth promotion in Arachis hypogea by Bacillus licheniformis A2. Microbiol. Res. 2014, 169, 66–75. [Google Scholar] [CrossRef]
- Shirinbayan, S.; Khosravi, H.; Malakouti, M.J. Alleviation of drought stress in maize (Zea mays) by inoculation with Azotobacter strains isolated from semi-arid regions. Appl. Soil Ecol. 2019, 133, 138–145. [Google Scholar] [CrossRef]
- Ekin, Z. Performance of phosphate solubilizing bacteria for improving growth and yield of sunflower (Helianthus annuusL.) in the presence of phosphorus fertilizer. Afr. J. Biotechnol. 2010, 9, 3794–3800. [Google Scholar]
- Rasul, M.; Yasmin, S.; Suleman, M.; Zaheer, A.; Reitz, T.; Tarkka, M.T.; Islam, E.; Mirza, M.S. Glucose dehydrogenase gene containing phosphobacteria for biofortification of Phosphorus with growth promotion of rice. Microbiol. Res. 2019, 223, 1–12. [Google Scholar] [CrossRef]
- Ogbe, A.A.; Finnie, J.F.; Van Staden, J. The role of endophytes in secondary metabolites accumulation in medicinal plants under abiotic stress. S. Afr. J. Bot. 2020, 134, 126–134. [Google Scholar] [CrossRef]
- Pfeilmeier, S.; Caly, D.L.; Malone, J.G. Bacterial pathogenesis of plants: Future challenges from a microbial perspective: Challenges in Bacterial Molecular Plant Pathology. Mol. Plant Pathol. 2016, 17, 1298–1313. [Google Scholar] [CrossRef] [PubMed]
- Passari, A.K.; Mishra, V.K.; Saikia, R.; Gupta, V.K.; Singh, B.P. Isolation, abundance and phylogenetic affiliation of endophytic actinomycetes associated with medicinal plants and screening for their in vitro antimicrobial biosynthetic potential. Front. Microbiol. 2015, 6, 273. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P.; Ferreira, B.S.; Cabral, J.M.S. Solvent tolerance in bacteria: Role of efflux pumps and cross-resistance with antibiotics. Int. J. Antimicrob. Agents 2003, 22, 211–216. [Google Scholar] [CrossRef]
- Yule, I.A.; Czaplewski, L.G.; Pommier, S.; Davies, D.T.; Narramore, S.K.; Fishwick, C.W. Pyridine-3-carboxamide-6-yl-ureas as novel inhibitors of bacterial DNA gyrase: Structure based design, synthesis, SAR and antimicrobial activity. Eur. J. Med. Chem. 2014, 86, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Sikora, K.; Jaśkiewicz, M.; Neubauer, D.; Bauer, M.; Bartoszewska, S.; Barańska-Rybak, W.; Kamysz, W. Counter-ion effect on antistaphylococcal activity and cytotoxicity of selected antimicrobial peptides. Amino Acids 2018, 50, 609–619. [Google Scholar] [CrossRef]
- Vlad, I.M.; Nuta, D.C.; Chirita, C.; Caproiu, M.T.; Draghici, C.; Dumitrascu, F.; Bleotu, C.; Avram, S.; Udrea, A.M.; Missir, A.V.; et al. In Silico and In Vitro Experimental Studies of New Dibenz[b,e]oxepin-11(6H)one O-(arylcarbamoyl)-oximes Designed as Potential Antimicrobial Agents. Molecules 2020, 25, 321. [Google Scholar] [CrossRef] [PubMed]
- Naeim, H.; El-Hawiet, A.; Abdel Rahman, R.A.; Hussein, A.; El Demellawy, M.A.; Embaby, A.M. Antibacterial activity of Centaurea pumilio L. root and aerial part extracts against some multidrug resistant bacteria. BMC Complement. Med. Ther. 2020, 20, 79. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Mo, J.; Guo, T.; Huang, S.; Li, Q.; Ning, P.; Hsiang, T. In vitro antifungal activity of dimethyl trisulfide against Colletotrichum gloeosporioides from mango. World J. Microbiol. Biotechnol. 2020, 36, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Al-Samman, A.M.M.A.; Siddique, N.A. Gas Chromatography–Mass Spectrometry (GC-MS/MS) Analysis, Ultrasonic Assisted Extraction, Antibacterial and Antifungal Activity of Emblica officinalis Fruit Extract. Pharmacogn. Mag. 2019, 11, 315–323. [Google Scholar] [CrossRef]
- Orienti, I.; Zuccari, G.; Bergamante, V.; Fini, A.; Carosio, R.; Montaldo, P.G. Enhancement of Oleyl Alcohol Anti Tumor Activity through Complexation in Polyvinyl alcohol Amphiphilic Derivatives. Drug Deliv. 2007, 14, 209–217. [Google Scholar] [CrossRef]
- Kalpana, V.N.; Rajeswari, V.D. 1—Preservatives in Beverages: Perception and Needs. In Preservatives and Preservation Approaches in Beverages; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 1–30. [Google Scholar]
- Priyanka, R.; Nakkeeran, S. Ochrobactrum ciceri mediated induction of defence genes and antifungal metabolites enhance the biocontrol efficacy for the management of Botrytis leaf blight of Lilium under protected conditions. J. Plant Pathol. 2019, 101, 323–337. [Google Scholar] [CrossRef]
- Awla, H.K.; Kadir, J.; Othman, R.; Rashid, T.S.; Wong, M.-Y. Bioactive compounds produced by Streptomyces sp. isolate UPMRS4 and antifungal activity against Pyricularia oryzae. Am. J. Plant Sci. 2016, 7, 1077. [Google Scholar]
- Manilal, A.; Tsalla, T.; Zerdo, Z.; Ameya, G.; Merdekios, B.; John, S.E. Evaluating the antibacterial and anticandidal potency of mangrove, Avicennia marina. Asian Pac. J. Trop. Dis. 2016, 6, 136–140. [Google Scholar] [CrossRef]
- Yogeswari, S.; Ramalakshmi, S.; Neelavathy, R.; Muthumary, J.y. Identification and comparative studies of different volatile fractions from Monochaetia kansensis by GCMS. Global J. Pharmacol. 2012, 6, 65–71. [Google Scholar]
- Khaled, J.M.; Alharbi, N.S.; Mothana, R.A.; Kadaikunnan, S.; Alobaidi, A.S. Biochemical Profile by GC–MS of Fungal Biomass Produced from the Ascospores of Tirmania nivea as a Natural Renewable Resource. J. Fungi 2021, 7, 1083. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamad, O.A.A.; Liu, Y.-H.; Huang, Y.; Li, L.; Ma, J.-B.; Egamberdieva, D.; Gao, L.; Fang, B.-Z.; Hatab, S.; Jiang, H.-C.; et al. The Metabolic Potential of Endophytic Actinobacteria Associated with Medicinal Plant Thymus roseus as a Plant-Growth Stimulator. Microorganisms 2022, 10, 1802. https://doi.org/10.3390/microorganisms10091802
Mohamad OAA, Liu Y-H, Huang Y, Li L, Ma J-B, Egamberdieva D, Gao L, Fang B-Z, Hatab S, Jiang H-C, et al. The Metabolic Potential of Endophytic Actinobacteria Associated with Medicinal Plant Thymus roseus as a Plant-Growth Stimulator. Microorganisms. 2022; 10(9):1802. https://doi.org/10.3390/microorganisms10091802
Chicago/Turabian StyleMohamad, Osama Abdalla Abdelshafy, Yong-Hong Liu, Yin Huang, Li Li, Jin-Biao Ma, Dilfuza Egamberdieva, Lei Gao, Bao-Zhu Fang, Shaimaa Hatab, Hong-Chen Jiang, and et al. 2022. "The Metabolic Potential of Endophytic Actinobacteria Associated with Medicinal Plant Thymus roseus as a Plant-Growth Stimulator" Microorganisms 10, no. 9: 1802. https://doi.org/10.3390/microorganisms10091802
APA StyleMohamad, O. A. A., Liu, Y.-H., Huang, Y., Li, L., Ma, J.-B., Egamberdieva, D., Gao, L., Fang, B.-Z., Hatab, S., Jiang, H.-C., & Li, W.-J. (2022). The Metabolic Potential of Endophytic Actinobacteria Associated with Medicinal Plant Thymus roseus as a Plant-Growth Stimulator. Microorganisms, 10(9), 1802. https://doi.org/10.3390/microorganisms10091802