
Corallococcus soli sp. Nov., a Soil Myxobacterium Isolated from Subtropical Climate, Chalus County, Iran, and Its Potential to Produce Secondary Metabolites

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Maintenance
2.2. Physiology and Chemotaxonomy
2.3. Genome and Phylogenetic Analysis
BiG-SCAPE Analysis
2.4. Extract Production, Antimicrobial Assay, and Extract Analysis
3. Results and Discussion
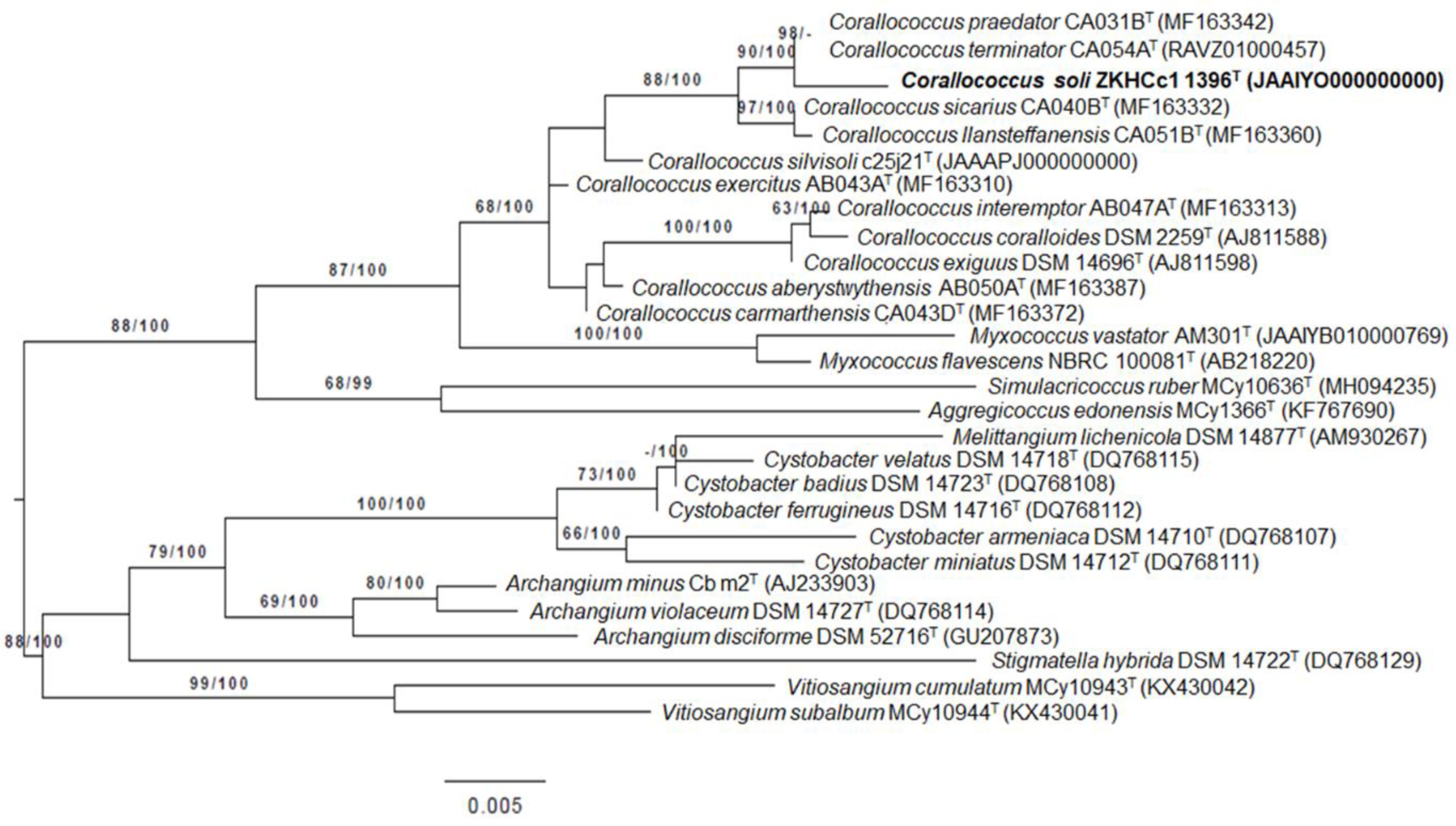
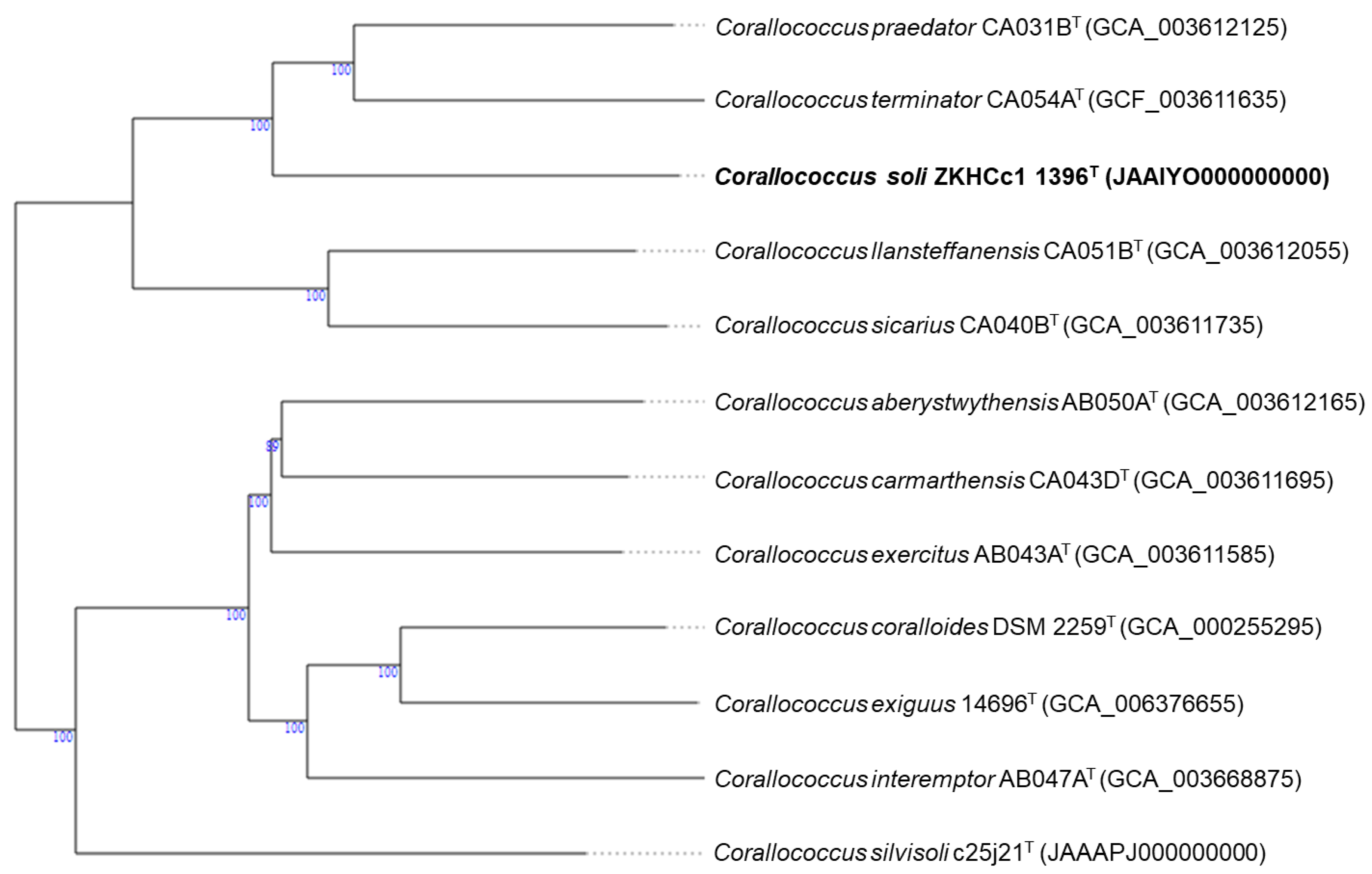
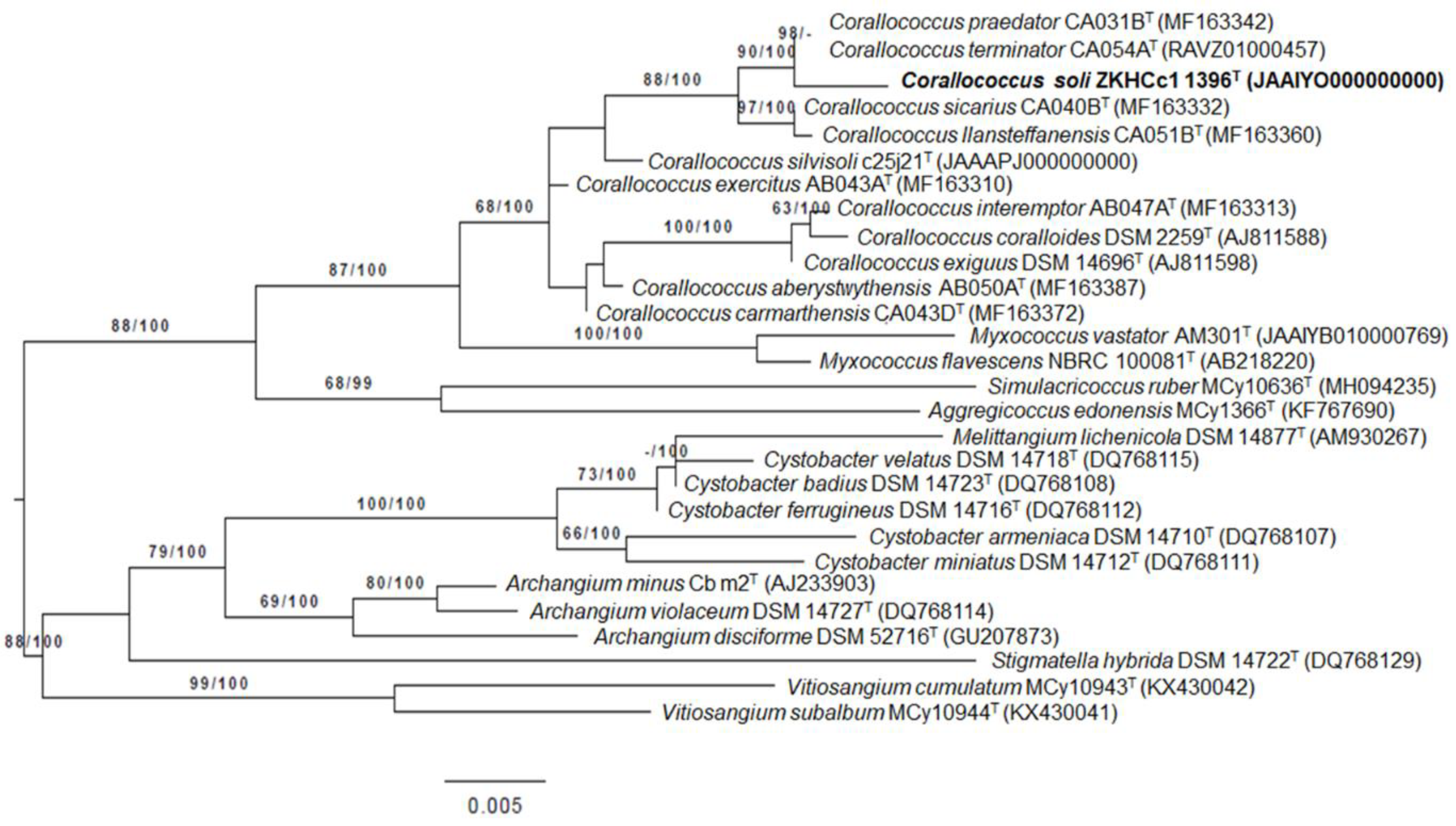
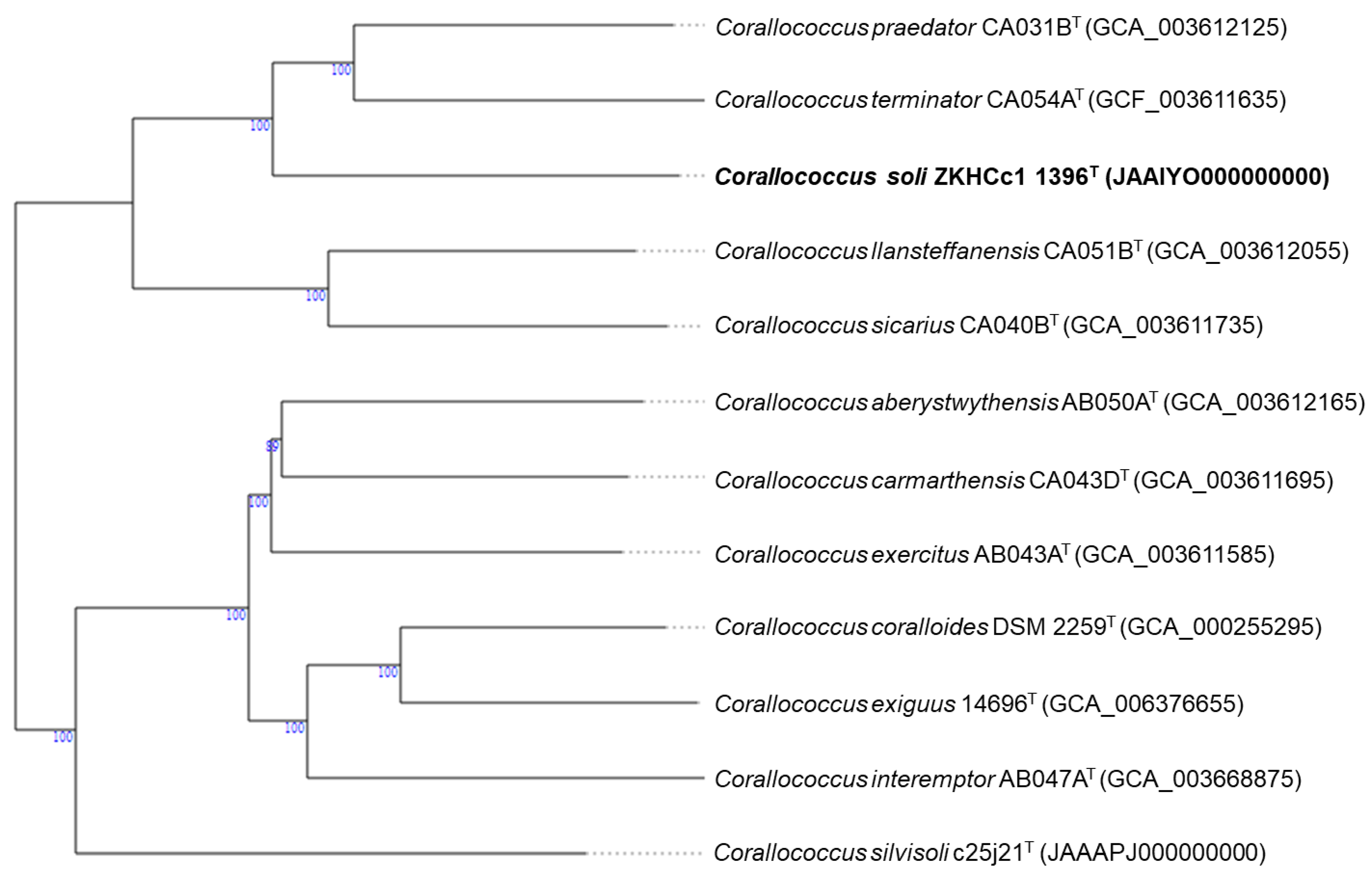
3.1. Taxonomic Identification
3.2. Comparison and Networking of the Secondary Metabolite Biosynthetic Gene Clusters (BGCs)
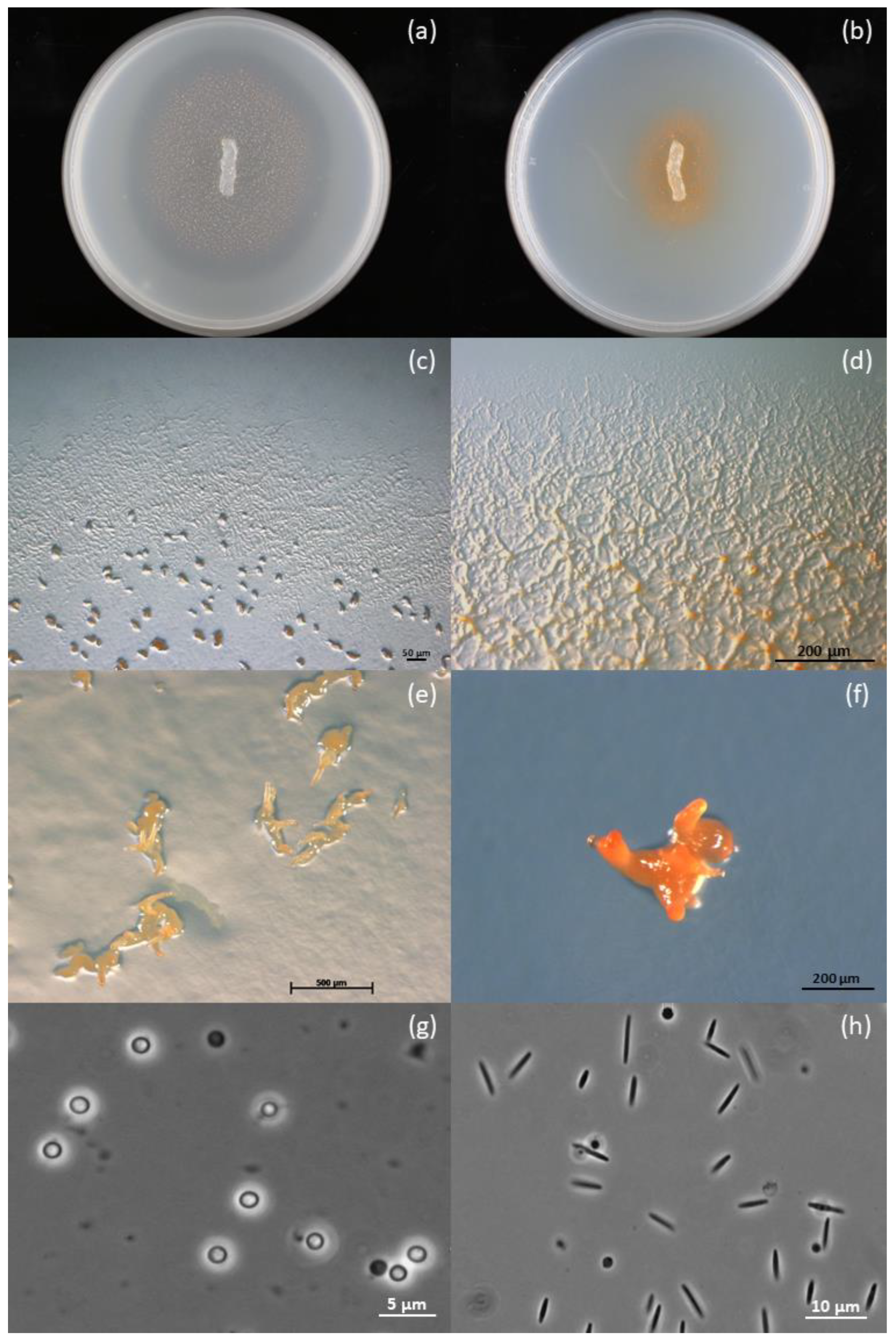
3.3. Description of Corallococcus soli sp. Nov.
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waite, D.W.; Chuvochina, M.; Pelikan, C.; Parks, D.H.; Yilmaz, P.; Wagner, M.; Loy, A.; Naganuma, T.; Nakai, R.; Whitman, W.B.; et al. Proposal to reclassify the proteobacterial classes Deltaproteobacteria and Oligoflexia, and the phylum Thermodesulfobacteria into four phyla reflecting major functional capabilities. Int. J. Syst. Evol. Microbiol. 2020, 70, 5972–6016. [Google Scholar] [CrossRef] [PubMed]
- Dawid, W. Biology and global distribution of myxobacteria in soils. FEMS Microbiol. Rev. 2000, 24, 403–427. [Google Scholar] [CrossRef] [PubMed]
- Reichenbach, H.; Dworkin, M. The myxobacteria. In The Prokaryotes, 2nd ed.; Balows, A., Trüper, H.G., Dworkin, M., Harder, W., Schleifer, K.H., Eds.; Springer: Berlin/Heidelberg, Germany, 1992; pp. 3416–3487. [Google Scholar]
- Shimkets, L.J.; Dworkin, M.; Reichenbach, H. The myxobacteria. In The Prokaryotes: A Handbook on the Biology of Bacteria; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 7, pp. 31–115. [Google Scholar]
- Garcia, R.; Müller, R. The family Polyangiaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 247–279. [Google Scholar]
- Iizuka, T.; Jojima, Y.; Fudou, R.; Yamanaka, S. Isolation of myxobacteria from the marine environment. FEMS Microbiol. Lett. 1998, 169, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.; La Clair, J.; Müller, R. Future directions of marine myxobacterial natural product discovery inferred from metagenomics. Mar. Drugs 2018, 16, 303. [Google Scholar] [CrossRef] [Green Version]
- Garcia, R.; Müller, R. The Family Haliangiaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 173–181. [Google Scholar]
- Garcia, R.; Müller, R. The Family Nannocystaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 213–229. [Google Scholar]
- Gerth, K.; Pradella, S.; Perlova, O.; Beyer, S.; Müller, R. Myxobacteria: Proficient producers of novel natural products with various biological activities–past and future biotechnological aspects with the focus on the genus Sorangium. J. Biotechnol. 2003, 106, 233–253. [Google Scholar] [CrossRef]
- Garcia, R.O.; Krug, D.; Müller, R. Chapter 3. Discovering natural products from myxobacteria with emphasis on rare producer strains in combination with improved analytical methods. In Methods in Enzymology; Hopwood, D., Ed.; Academic Press: Burlington, NJ, USA, 2009; Volume 458, pp. 59–91. [Google Scholar]
- Plaza, A.; Müller, R. Chapter 6 Myxobacteria: Chemical diversity and screening strategies. In Natural Products: Discourse, Diversity, and Design, 1st ed.; Osbourn, A., Goss, R.J., Carter, G.T., Eds.; Wiley Blackwell: Hoboken, NJ, USA, 2014; pp. 103–124. [Google Scholar]
- Weissman, K.J.; Müller, R. Myxobacterial secondary metabolites: Bioactivities and modes-of-action. Nat. Prod. Rep. 2010, 27, 1276–1295. [Google Scholar] [CrossRef]
- Shimkets, L.; Woese, C.R. A phylogenetic analysis of the myxobacteria: Basis for their classification. Proc. Natl. Acad. Sci. USA 1992, 89, 9459–9463. [Google Scholar] [CrossRef] [Green Version]
- Spröer, C.; Reichenbach, H.; Stackebrandt, E. The correlation between morphological and phylogenetic classification of myxobacteria. Int. J. Syst. Bacteriol. 1999, 49, 1255–1262. [Google Scholar] [CrossRef]
- Garcia, R.; Gerth, K.; Stadler, M.; Dogma, I.J.; Müller, R. Expanded phylogeny of myxobacteria and evidence for cultivation of the ‘unculturables’. Mol. Phylogenet. Evol. 2010, 57, 878–887. [Google Scholar] [CrossRef]
- Garcia, R.; Gemperlein, K.; Müller, R. Minicystis rosea gen. nov., sp. nov., a polyunsaturated fatty acid-rich and steroid-producing soil myxobacterium. Int. J. Syst. Evol. Microbiol. 2014, 64, 3733–3742. [Google Scholar] [CrossRef]
- Garcia, R.; Stadler, M.; Gemperlein, K.; Müller, R. Aetherobacter fasciculatus gen. nov., sp. nov. and Aetherobacter rufus sp. nov., novel myxobacteria with promising biotechnological applications. Int. J. Syst. Evol. Microbiol. 2016, 66, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.; Müller, R. Simulacricoccus ruber gen. nov., sp. nov., a microaerotolerant, non-fruiting, myxospores forming soil myxobacterium and emended description of the family Myxococcaceae. Int. J. Syst. Evol. Microbiol. 2018, 68, 3101–3110. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.; Reichenbach, H.; Ring, M.W.; Müller, R. Phaselicystis flava gen. nov., sp. nov., an arachidonic acid- containing soil myxobacterium, and the description of Phaselicystidaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 1524–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohr, K.I.; Garcia, R.O.; Gerth, K.; Irschik, H.; Müller, R. Sandaracinus amylolyticus gen. nov., sp. nov., a starch-degrading soil myxobacterium, and description of Sandaracinaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2012, 62, 1191–1198. [Google Scholar] [CrossRef] [Green Version]
- Livingstone, P.G.; Ingleby, O.; Girdwood, S.; Cookson, A.R.; Morphew, R.M.; Whitworth, D.E. Predatory organisms with untapped biosynthetic potential: Descriptions of novel Corallococcus species C. aberystwythensis sp. nov., C. carmarthensis sp. nov., C. exercitus sp. nov., C. interemptor sp. nov., C. llansteffanensis sp. nov., C. praedator sp. nov., C. sicarius sp. nov., and C. terminator sp. nov. Appl. Environ. Microbiol. 2020, 86, e01931-19. [Google Scholar]
- Garcia, R.; Müller, R. The family Myxococcaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 191–212. [Google Scholar]
- Jansen, R.; Höfle, G.; Irschik, H.; Reichenbach, H. Antibiotika aus Gleitenden Bakterien, XXIV. Corallopyronin A, B und C–drei neue Antibiotika aus Corallococcus coralloides Cc c127 (Myxobacterales). Liebigs Ann. Chem. 1985, 4, 822–836. [Google Scholar] [CrossRef]
- Schmitz, A.; Kehraus, S.; Schäberle, T.F.; Neu, E.; Almeida, C.; Roth, M.; König, G.M. Corallorazines from the myxobacterium Corallococcus coralloides. J. Nat. Prod. 2014, 77, 159–163. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, H.J.; Kim, G.W.; Cho, K.; Takahashi, S.; Koshino, H.; Kim, W.G. Isolation of coralmycins A and B, potent anti-Gram negative compounds from the myxobacteria Corallococcus coralloides M23. J. Nat. Prod. 2016, 79, 2223–2228. [Google Scholar] [CrossRef]
- Erol, Ö.; Schäberle, T.F.; Schmitz, A.; Rachid, S.; Gurgui, C.; El Omari, M.; Lohr, F.; Kehraus, S.; Piel, J.; Müller, R.; et al. Biosynthesis of the myxobacterial antibiotic corallopyronin A. ChemBioChem 2010, 11, 1253–1265. [Google Scholar] [CrossRef]
- Gerhardt, P.; Murray, R.G.E.; Costilow, R.N.; Nester, E.W.; Wood, W.A.; Krieg, N.R.; Phillips, G.E. (Eds.) Manual of Methods for General Bacteriology; American Society for Microbiology: Washington, DC, USA, 1981; pp. 415–416. [Google Scholar]
- Garcia, R.; Pistorius, D.; Stadler, M.; Müller, R. Fatty acid-related phylogeny of myxobacteria as an approach to discover polyunsaturated omega-3/6 fatty acids. J. Bacteriol. 2011, 193, 1930–1942. [Google Scholar] [CrossRef] [Green Version]
- Gemperlein, K.; Rachid, S.; Garcia, R.O.; Wenzel, S.C.; Müller, R. Polyunsaturated fatty acid biosynthesis inmyxobacteria: Different PUFA synthases and their product diversity. Chem. Sci. 2014, 5, 1733–1741. [Google Scholar] [CrossRef]
- Hall, T.A. BIOEDIT: A user-friendly biological sequence alignment editor and analysis program from windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2021, 50, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M.; Spröer, C.; Klenk, H.-P. When should a DDH experiment be mandatory in microbialtaxonomy? Arch. Microbiol. 2013, 195, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Hahnke, R.L.; Petersen, J.; Scheuner, C.; Michael, V.; Fiebig, A.; Rohde, C.; Rohde, M.; Fartmann, B.; Goodwin, L.A.; et al. Complete genome sequence of DSM 30083T, the type strain (U5/41T) of Escherichia coli, and a proposal for delineating subspecies in microbial taxonomy. Stand. Genom. Sci. 2014, 10, 2. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Farris, J.S.; Nixon, K.C. TNT, a free program for phylogenetic analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Pattengale, N.D.; Alipour, M.; Bininda-Emonds, O.R.P.; Moret, B.M.E.; Stamatakis, A. How many bootstrap replicates are necessary? J. Comput. Biol. 2010, 17, 337–354. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0 b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; O’Neill, K.R.; Haft, D.H.; DiCuccio, M.; Chetvernin, V.; Badretdin, A.; Coulouris, E.; Chitsaz, F.; Derbyshire, M.K.; Durkin, A.S.; et al. RefSeq: Expanding the Prokaryotic Genome Annotation Pipeline reach with protein family model curation. Nucleic Acids Res. 2021, 49, D1020–D1028. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Chalita, M.; Ha, S.M.; Na, S.I.; Yoon, S.H.; Chun, J. ContEst16S: An algorithm that identifies contaminated prokaryotic genomes using 16S RNA gene sequences. Int. J. Syst. Evol. Microbiol. 2017, 67, 2053–2057. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [PubMed]
- Parte, A.C. LPSN–List of Prokaryotic names with Standing in Nomenclature (bacterio. net), 20 years on. Int. J. Syst. Evol. Microbiol. 2018, 68, 1825–1829. [Google Scholar] [CrossRef] [PubMed]
- Ondov, B.D.; Treangen, T.J.; Melsted, P.; Mallonee, A.B.; Bergman, N.H.; Koren, S.; Phillippy, A.M. Mash: Fast genome and metagenome distance estimation using MinHash. Genome Biol. 2016, 17, 132. [Google Scholar] [CrossRef] [Green Version]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Staerfeldt, H.H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A comprehensive, accurate, and fast distance-based phylogeny inference program. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef] [Green Version]
- Farris, J.S. Estimating phylogenetic trees from distance matrices. Am. Nat. 1972, 106, 645–667. [Google Scholar] [CrossRef]
- Kreft, L.; Botzki, A.; Coppens, F.; Vandepoele, K.; Van Bel, M. PhyD3: A phylogenetic tree viewer with extended phyloXML support for functional genomics data visualization. Bioinformatics 2017, 33, 2946–2947. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Lim, J.M.; Kwon, S.J.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. AntiSMASH: Rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. AntiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Kautsar, S.A.; Blin, K.; Shaw, S.; Navarro-Muñoz, J.C.; Terlouw, B.R.; Van Der Hooft, J.J.J.; Van Santen, J.A.; Tracanna, V.; Duran, H.G.S.; Andreu, V.P.; et al. MIBiG 2.0: A repository for biosynthetic gene clusters of known function. Nucleic Acids Res. 2020, 48, 454–458. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Muñoz, J.C.; Selem-Mojica, N.; Mullowney, M.W.; Kautsar, S.A.; Tryon, J.H.; Parkinson, E.I.; De Los Santos, E.L.; Yeong, M.; Cruz-Morales, P.; Abubucker, S.; et al. A computational framework to explore large-scale biosynthetic diversity. Nat. Chem. Biol. 2020, 16, 60–68. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.; Tosatto, S.C.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families’ database in 2021. Nucleic Acids Res. 2021, 49, 412–419. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2013, 13, 2498–2504. [Google Scholar] [CrossRef]
- Cazin, J., Jr.; Wiemer, D.F.; Howard, J.J. Isolation, growth characteristics, and long-term storage of fungi cultivated by attine ants. Appl. Environ. Microbial. 1989, 55, 1346–1350. [Google Scholar] [CrossRef] [Green Version]
- Khosravi Babadi, Z.; Ebrahimipour, G.; Wink, J.; Narmani, A.; Risdian, C. Isolation and identification of Streptomyces sp. Act4Zk, a good producer of Staurosporine and some derivatives. Lett. Appl. Microbiol. 2021, 72, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Primahana, G.; Risdian, C.; Mozef, T.; Wink, J.; Surup, F.; Stadler, M. Amycolatomycins A and B, Cyclic Hexapeptides Isolated from an Amycolatopsis sp. 195334CR. Antibiotics 2021, 10, 261. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Feng, G.D.; Liu, Y.; Zhou, Y.; Deng, X.; Yao, Q.; Zhu, H. Corallococcus silvisoli sp. nov., a novel myxobacterium isolated from subtropical forest soil. Arch. Microbiol. 2022, 204, 141. [Google Scholar] [CrossRef] [PubMed]
- Ahearne, A.; Albataineh, H.; Dowd, S.E.; Stevens, D.C. Assessment of Evolutionary Relationships for Prioritization of Myxobacteria for Natural Product Discovery. Microorganisms 2021, 9, 1376. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains: | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temp. (°C): 30 | ++ | ++ | ++ | + | + | ++ | +++ | ++ | ++ | ++ | +++ | +++ |
| 35 | +++ | ++ | + | ++ | - | ++ | ++ | + | - | + | + | ++ |
| pH: 5 | - | + | + | - | - | - | + | - | - | - | + | ND |
| 6 | +++ | ++ | + | - | + | + | + | + | + | ++ | ++ | +++ |
| 7 | +++ | +++ | ++ | + | + | ++ | +++ | ++ | ++ | ++ | +++ | +++ |
| 8 | +++ | ++ | ++ | - | - | ++ | +++ | ++ | ++ | +++ | +++ | ++ |
| 9 | ++ | ++ | ++ | - | - | ++ | +++ | ++ | ++ | +++ | + | + |
| Biochemical | ||||||||||||
| Esculin test | - | - | + | - | + | + | + | + | + | + | - | + |
| Gelatine test | + | + | + | - | - | + | + | - | + | + | + | + |
| Glucose assimilation | - | + | - | - | - | - | - | - | - | - | - | + |
| Maltose assimilation | - | - | - | - | - | - | - | - | + | - | + | + |
| Nitrate reduction | - | - | + | - | - | - | - | - | - | - | - | - |
| Antibiotic sensitivity | ||||||||||||
| Cefotaxime | - | + | + | + | + | + | - | + | + | + | + | - |
| Ceftazidime | - | + | + | + | + | + | + | + | + | + | + | - |
| Gentamicin | + | + | + | + | + | + | - | + | + | + | + | - |
| Trimethoprim- sulfamethoxazole | + | - | - | - | - | - | - | - | - | - | - | - |
| Genome comparison: | ||||||||||||
| Contigs | 68 | 961 | 459 | 625 | 1491 | 802 | 530 | 1244 | 863 | 1 | 36 | 62 |
| Genome size (Mb) | 9.44 | 10.15 | 9.47 | 9.98 | 10.51 | 10.39 | 10.79 | 10.53 | 10.35 | 10.08 | 10.41 | 9.23 |
| Mol% GC | 69.8 | 70.2 | 70.0 | 70.0 | 69.7 | 70.2 | 69.9 | 70.3 | 69.5 | 69.9 | 69.6 | 69.8 |
| No. of Gene | 7445 | 8611 | 7892 | 8353 | 9011 | 8442 | 8959 | 8867 | 8506 | 8148 | 8416 | 7412 |
| Pseudogene | 140 | 272 | 216 | 211 | 286 | 307 | 257 | 270 | 197 | 123 | 159 | 131 |
| No. of Protein | 7248 | 8271 | 7612 | 8079 | 8661 | 8072 | 8639 | 8539 | 8247 | 7952 | 8192 | 7221 |
| rRNA | 3 | 7 | 7 | 6 | 7 | 6 | 6 | 3 | 3 | 9 | 3 | 3 |
| tRNA | 50 | 57 | 53 | 53 | 54 | 52 | 53 | 51 | 55 | 60 | 58 | 53 |
| Other RNA | 4 | 4 | 4 | 4 | 3 | 5 | 4 | 4 | 4 | 4 | 4 | 4 |
| Fatty Acid | % |
|---|---|
| C10:0 | tr |
| C14:0 | tr |
| C14:1 | tr |
| C15:0 | tr |
| C16:0 | 1.3 |
| C16:1 | 0.9 |
| C18:0 | 1.3 |
| C18:2 ω6,9 all cis | 0.2 |
| C18:3ω6,9,12 all cis | 1.3 |
| C16:0 2-OH | 0.1 |
| C16:0 3-OH | tr |
| Total SCFA: | 5.1 |
| iso-C11:0 | 0.3 |
| iso-C12:0 | tr |
| iso-C13:0 | 2.4 |
| iso-C14:0 | 1.9 |
| iso-C15:0 | 15.8 |
| iso-C15:1 | 1.9 |
| iso-C16:0 | 5.6 |
| iso-C16:1 | 0.5 |
| iso-C17:0 | 9.4 |
| iso-C17:1 | 11.7 |
| iso-C17:2 | 0.9 |
| iso-C15:0 3-OH | 5.8 |
| iso-C16:0 2-OH | 0.7 |
| iso-C16:0 3-OH | tr |
| iso-C17:0 2-OH | 31.0 |
| iso-C18:0 2-OH | 0.1 |
| iso-C15:0 OAG | 1.3 |
| iso-C16:0 OAG | tr |
| iso-C15:0 DMA | 5.0 |
| Total BCFA: | 94.2 |
| ANI/dDDH Value (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| dDDH\ANI | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| 1 | 100 | 92 | 91 | 89 | 89 | 87 | 87 | 87 | 86 | 86 | 86 | 87 |
| 2 | 44 | 100 | 91 | 88 | 87 | 84 | 84 | 84 | 85 | 86 | 87 | 87 |
| 3 | 43 | 50 | 100 | 88 | 87 | 84 | 84 | 84 | 84 | 85 | 86 | 87 |
| 4 | 36 | 36 | 35 | 100 | 92 | 85 | 85 | 86 | 85 | 86 | 86 | 87 |
| 5 | 36 | 37 | 35 | 50 | 100 | 85 | 85 | 85 | 85 | 86 | 87 | 88 |
| 6 | 31 | 32 | 31 | 32 | 32 | 100 | 91 | 91 | 90 | 92 | 92 | 89 |
| 7 | 31 | 31 | 30 | 31 | 32 | 47 | 100 | 91 | 90 | 91 | 91 | 89 |
| 8 | 31 | 31 | 31 | 31 | 33 | 48 | 48 | 100 | 90 | 91 | 92 | 89 |
| 9 | 30 | 30 | 30 | 30 | 31 | 42 | 43 | 42 | 100 | 92 | 92 | 88 |
| 10 | 30 | 31 | 30 | 30 | 31 | 44 | 44 | 44 | 46 | 100 | 94 | 88 |
| 11 | 30 | 30 | 30 | 30 | 31 | 43 | 43 | 44 | 44 | 54 | 100 | 88 |
| 12 | 32 | 32 | 31 | 32 | 33 | 36 | 35 | 36 | 34 | 35 | 34 | 100 |
| Corallococcus Species | Accession Number | Percentage Similarity of BGC | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||
| C.soli ZKHCc1 1396T | JAAIYO000000000 | 100 | 100 | 83 | 100 | 100 | 88 | 100 | 100 | 100 | - | - | - |
| C. praedatorCA031BT | RAWI01000000 | 100 | 80 | 33 | 100 | 100 | - | - | - | 100 | - | - | - |
| C. terminatorCA054AT | RAVZ01000000 | 100 | 80 | 83 | 100 | 100 | 88 | 100 | 100 | 100 | 100 | - | - |
| C. sicariusCA040BT | RAWG01000000 | 45 | 80 | 75 | 100 | - | 22 | - | - | 100 | - | - | - |
| C. llansteffanensisCA051BT | RAWB01000000 | 27 | 80 | 83 | 100 | 100 | - | - | 100 | 100 | - | 100 | - |
| C. exercitusAB043AT | RAVW01000000 | 90 | 80 | 33 | 100 | 100 | - | - | 100 | 100 | 100 | - | - |
| C. aberystwythensisAB050AT | RAWK01000000 | 63 | 100 | 75 | 100 | - | - | - | 100 | 100 | - | - | 100 |
| C. carmarthensisCA043DT | RAWE01000000 | 100 | 100 | 83 | 100 | 100 | - | - | - | 100 | - | - | - |
| C. interemptorAB047AT | RAWM01000000 | 100 | 60 | 83 | 100 | - | - | - | 100 | 100 | - | - | - |
| C. coralloidesDSM 2259T | CP003389 | 100 | 100 | 83 | 100 | - | - | - | - | - | - | - | - |
| C. exiguusDSM 14696T | JAAAPK010000000 | 100 | 100 | 83 | 100 | - | - | 100 | - | - | - | - | - |
| C. silvisoli c25j21T | JAAAPJ000000000 | 100 | 100 | 83 | 100 | 100 | - | - | 100 | 100 | 100 | - | - |
| Fraction | RT (min) | Major Ion | Number of Hits in DNP | |
|---|---|---|---|---|
| m/z | Ion | |||
| 1 | 1.87–3.99 | 211.1440 233.1257 | [M + H]+ [M + Na]+ | 28 |
| 2 | 4.10–6.00 | 245.1282 267.1100 | [M + H]+ [M + Na]+ | 61 |
| 3 | 6.09–8.02 | 209.1645 | [M + H]+ | 21 (2 from Chondromyces crocatus) |
| 4 | 8.12–10.03 | 223.1800 | [M + H]+ | 7 (2 from Chondromyces crocatus) |
| 277.2156 295.2262 313.2368 | [M-2H2O + H]+ [M-H2O + H]+ [M + H]+ | 59 | ||
| 353.2292 683.4689 | [M + Na]+ [2M + Na]+ | 552 | ||
| 5 | 10.18–11.99 | 277.2163 295.2264 | [M-H2O + H]+ [M + H]+ | 135 |
| 335.2192 647.4485 | [M + Na]+ [2M + Na]+ | 11 | ||
| 6 | 12.10–14.01 | 319.2243 | [M + H]+ | 1268 |
| 317.2086 611.4280 | [M + Na]+ [2M + Na]+ | 2 | ||
| 279.2318 557.4566 | [M + H]+ [M + Na]+ | 134 | ||
| 277.2159 295.2255 | [M-H2O + H]+ [M + H]+ | 135 | ||
| 293.2086 | [M + H]+ | 113 | ||
| 7 | 14.12–15.99 | 255.2317 277.2136 237.2210 509.4564 | [M + H]+ [M + Na]+ [M-H2O + H]+ [2M + H]+ | 91 |
| 325.2712 | [M + H]+ | 169 | ||
| 8 | 16.11–18.01 | 353.2661 313.2736 | [M + Na]+ [M-H2O + H]+ | 18 |
| 257.2471 239.2365 | [M + H]+ [M-H2O + H]+ | 40 | ||
| 441.3549 | [M + H]+ | 5 | ||
| 485.3808 | [M + H]+ | 1 | ||
| 524.4517 | [M + H]+ | 1 | ||
| 529.4070 | [M + H]+ | 3 | ||
| 9 | 18.11–19.98 | 686.4746 708.4566 1371.9401 | [M + H]+ [M + Na]+ [2M + H]+ | 0 |
| 589.4437 1155.8960 | [M + Na]+ [2M + Na]+ | 3 | ||
| 353.3021 | [M + H]+ | 5 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Babadi, Z.K.; Garcia, R.; Ebrahimipour, G.H.; Risdian, C.; Kämpfer, P.; Jarek, M.; Müller, R.; Wink, J. Corallococcus soli sp. Nov., a Soil Myxobacterium Isolated from Subtropical Climate, Chalus County, Iran, and Its Potential to Produce Secondary Metabolites. Microorganisms 2022, 10, 1262. https://doi.org/10.3390/microorganisms10071262
Babadi ZK, Garcia R, Ebrahimipour GH, Risdian C, Kämpfer P, Jarek M, Müller R, Wink J. Corallococcus soli sp. Nov., a Soil Myxobacterium Isolated from Subtropical Climate, Chalus County, Iran, and Its Potential to Produce Secondary Metabolites. Microorganisms. 2022; 10(7):1262. https://doi.org/10.3390/microorganisms10071262
Chicago/Turabian StyleBabadi, Zahra Khosravi, Ronald Garcia, Gholam Hossein Ebrahimipour, Chandra Risdian, Peter Kämpfer, Michael Jarek, Rolf Müller, and Joachim Wink. 2022. "Corallococcus soli sp. Nov., a Soil Myxobacterium Isolated from Subtropical Climate, Chalus County, Iran, and Its Potential to Produce Secondary Metabolites" Microorganisms 10, no. 7: 1262. https://doi.org/10.3390/microorganisms10071262
APA StyleBabadi, Z. K., Garcia, R., Ebrahimipour, G. H., Risdian, C., Kämpfer, P., Jarek, M., Müller, R., & Wink, J. (2022). Corallococcus soli sp. Nov., a Soil Myxobacterium Isolated from Subtropical Climate, Chalus County, Iran, and Its Potential to Produce Secondary Metabolites. Microorganisms, 10(7), 1262. https://doi.org/10.3390/microorganisms10071262