
Native Heavy Metal-Tolerant Plant Growth Promoting Rhizobacteria Improves Sulla spinosissima (L.) Growth in Post-Mining Contaminated Soils



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Testing Resistance to Heavy Metals in Bacterial Isolates from Stipa Tenuissima Rhizosphere
2.3. Evaluation of PGP Traits in Bacterial Strains from Stipa Tenuissima Rhizosphere
2.4. In Planta Evaluation of the PGP Traits of the Selected Strains
2.4.1. Measurement of Plant Biomass and Chlorophyll/Carotenoid Content
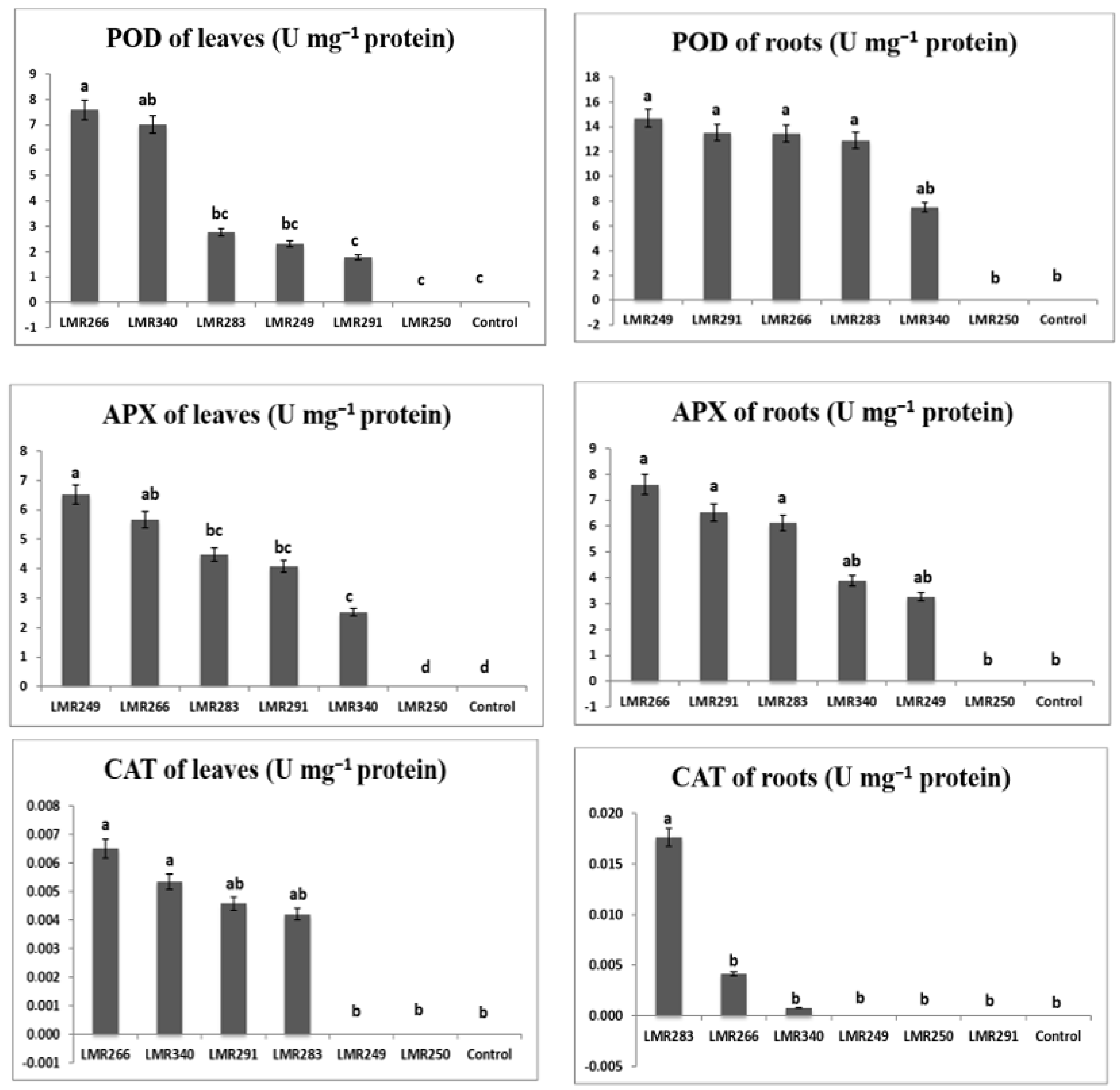
2.4.2. Antioxidant Enzyme Assays
2.5. Effect of Inoculation with Selected PGPR Strains on Sulla Growth under Heavy Metal Stress Conditions
2.6. Statistical Analysis
3. Results
3.1. Comparative Heavy Metal Resistance of Bacterial Strains Used as Inocula
3.2. PGP Activities in Bacterial Strains Used as Inocula
3.3. Beneficial Effect of Selected Tolerant PGPR Strains on Sulla spinossisima (L.) Growing in Sterilized Sand
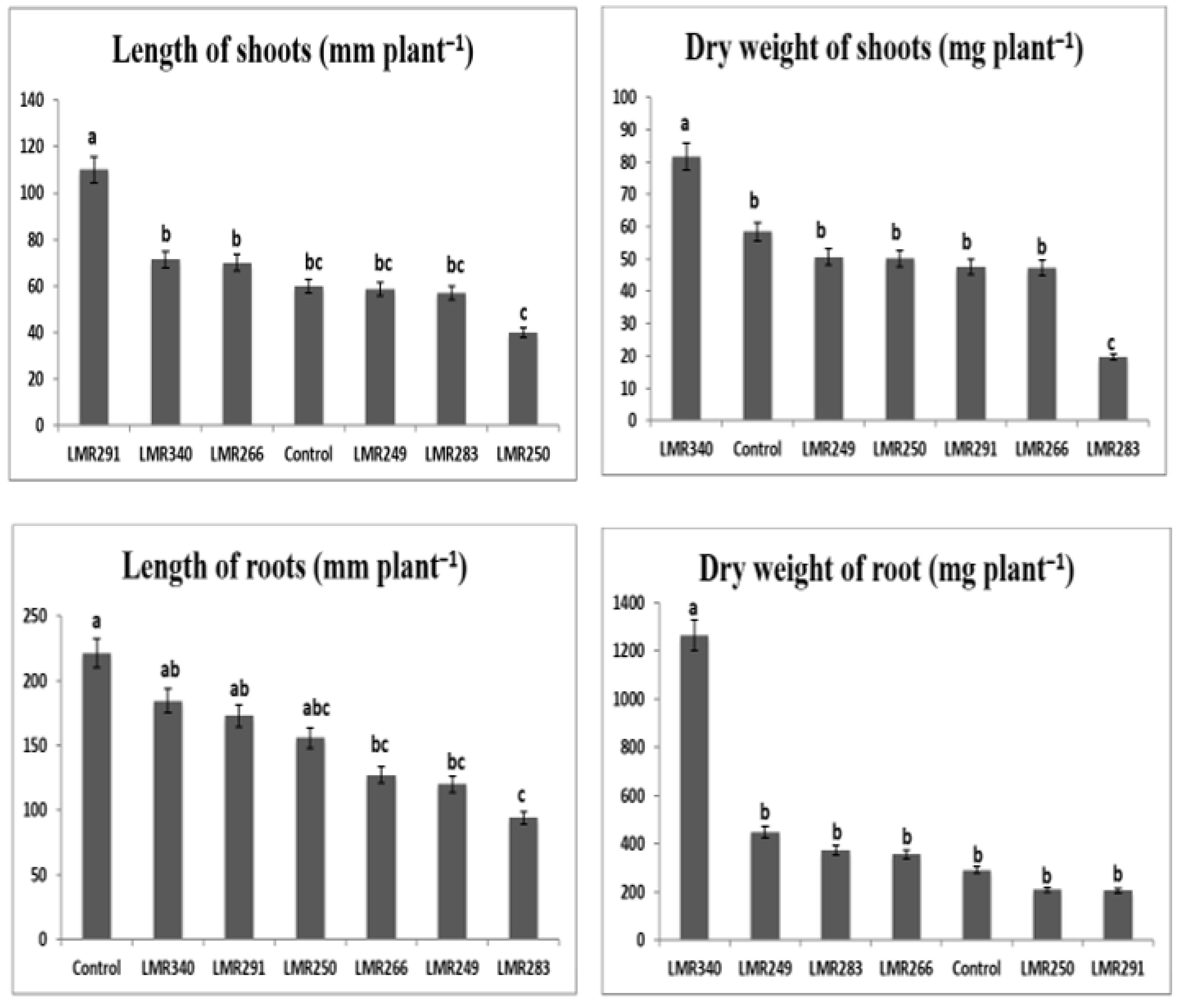
3.3.1. Plant Growth Parameter Measurements
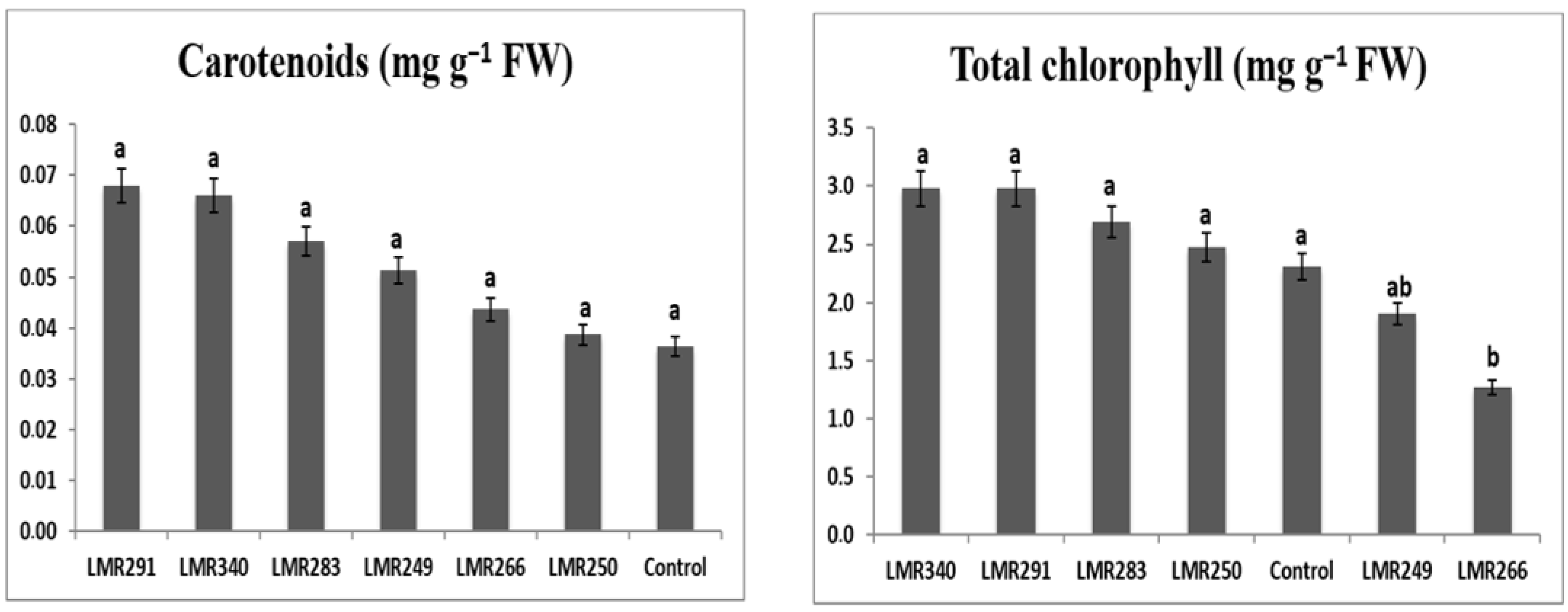
3.3.2. Plant Chlorophyll and Carotenoid Content
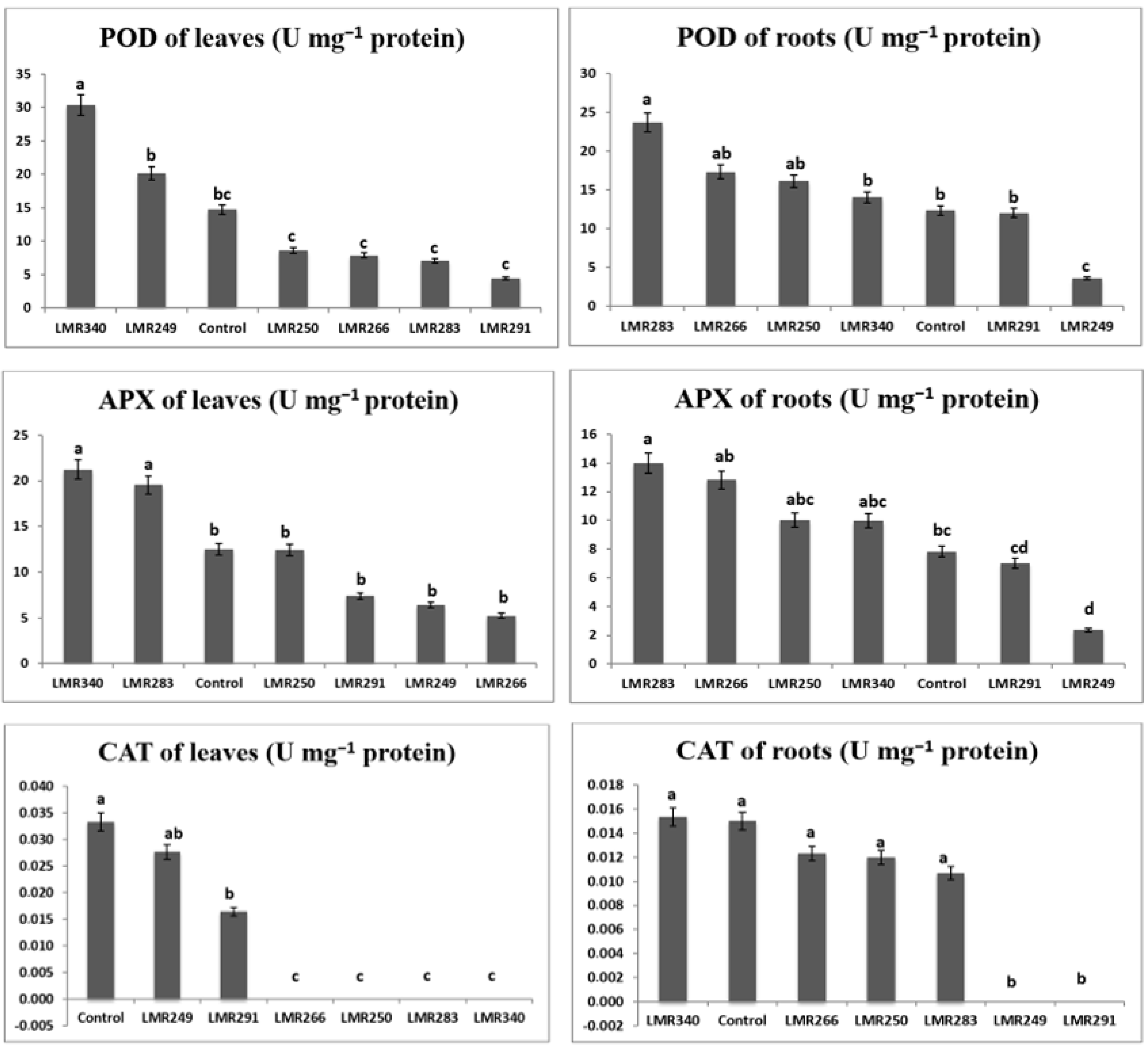
3.3.3. Plant Antioxidant Enzymes Activities
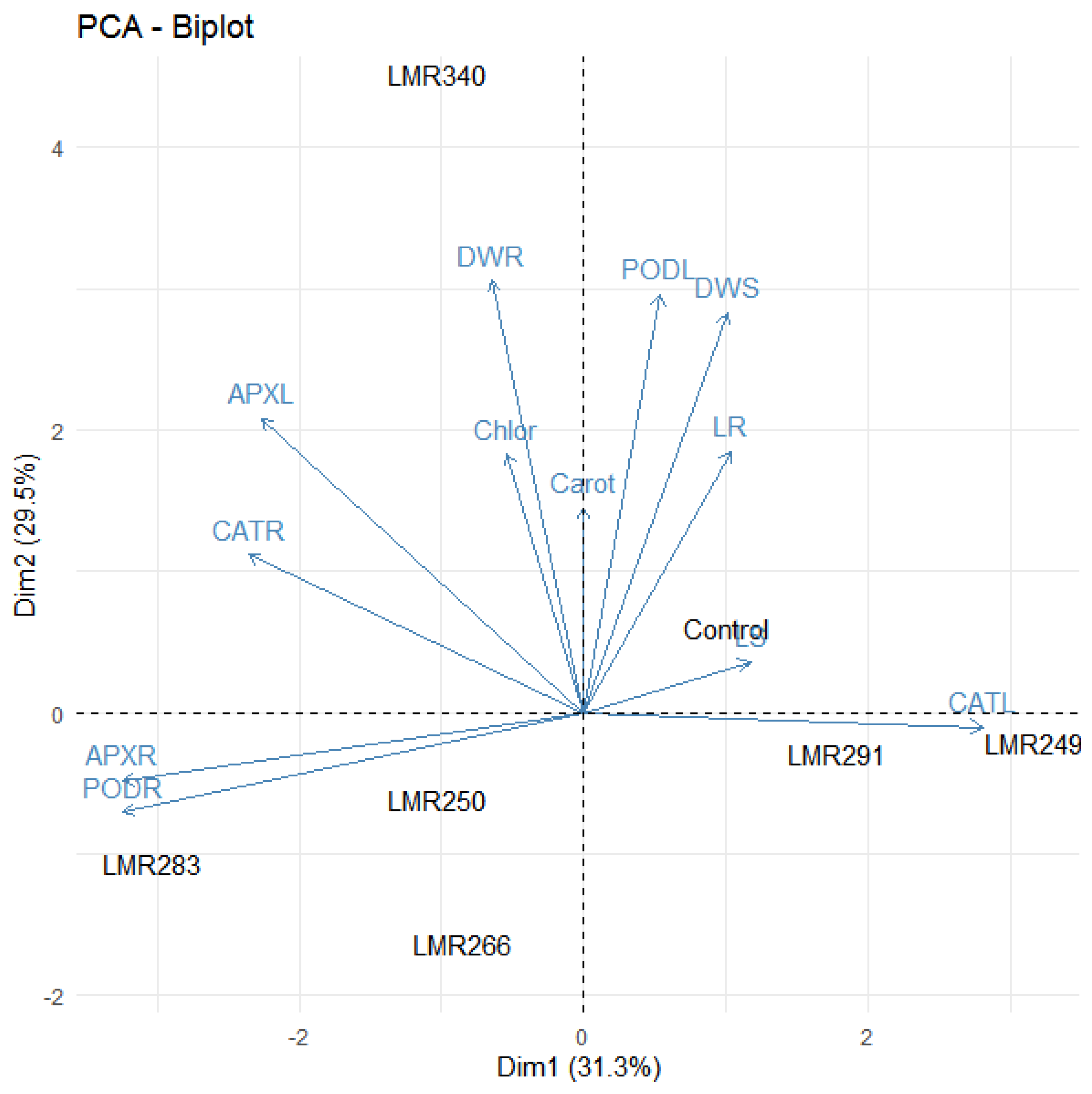
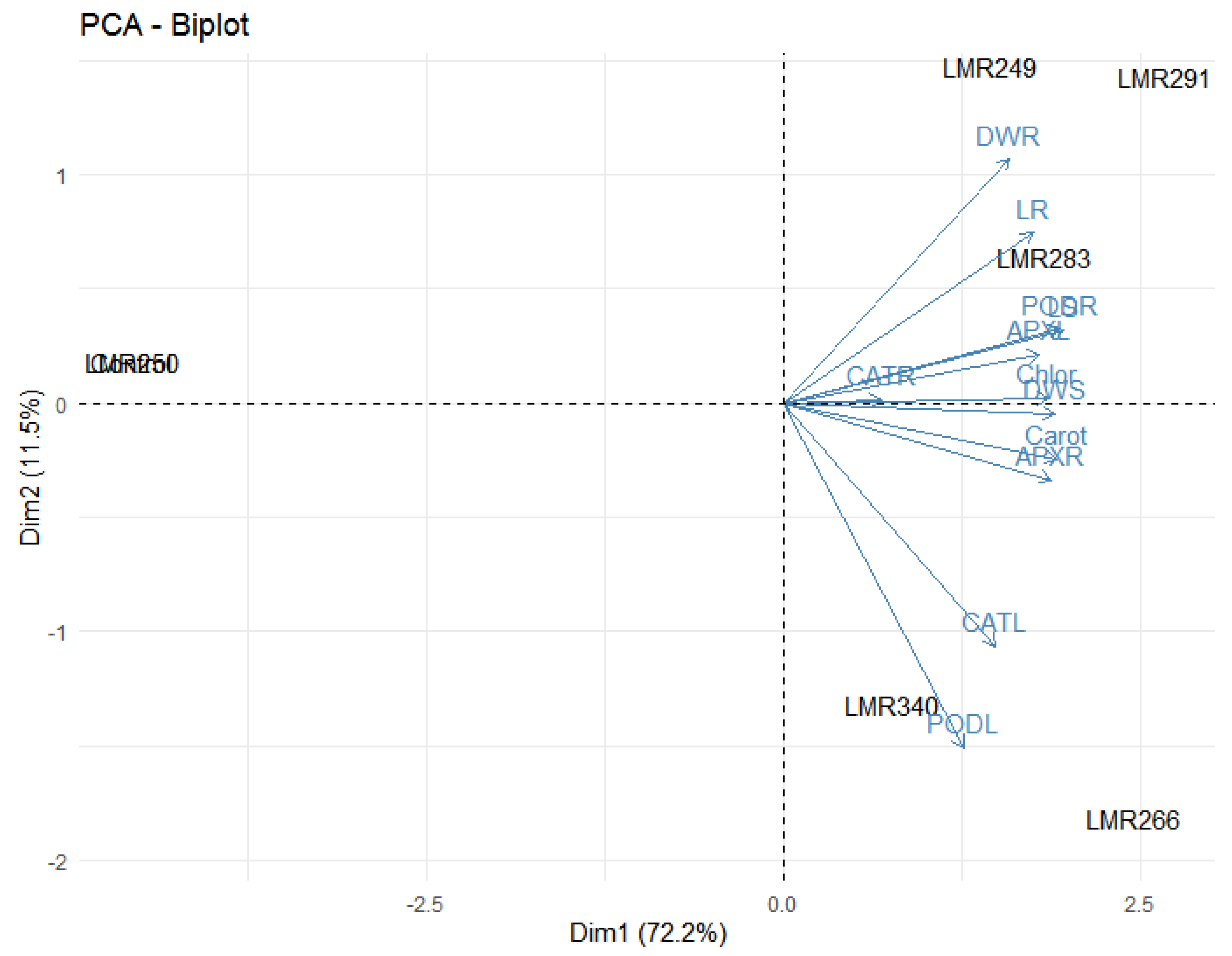
3.3.4. PCA Analysis
3.4. Effect of Selected Heavy Metal-Tolerant PGPR Strains on Sulla spinosissima (L.) Growing in a Contaminated Soil
3.4.1. Plant Growth Parameter Measurements
3.4.2. Plant Chlorophyll and Carotenoid Content
3.4.3. Plant Antioxidant Enzymes Activities
3.4.4. PCA Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pilon-Smits, E. Phytoremediation. Annu. Rev. Plant Biol. 2005, 56, 15–39. [Google Scholar] [CrossRef] [PubMed]
- Tekere, M. Biological strategies for heavy metal remediation. In Methods for Bioremediation of Water and Wastewater Pollution, Environmental Chemistry for a Sustainable World; Ahamed, M.I.I., Lichtfouse, E., Asiri, A.M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 393–413. [Google Scholar]
- Agnello, A.C.; Bagard, M.; van Hullebusch, E.D.; Esposito, G.; Huguenot, D. Comparative bioremediation of heavy metals and petroleum hydrocarbons co-contaminated soil by natural attenuation, phytoremediation, bioaugmentation and bioaugmentation-assisted phytoremediation. Sci. Total Environ. 2016, 563–564, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, A.; Ji, C.; Joseph, S.; Bian, R.; Li, L.; Pan, G.; Paz-Ferreiro, J. Biochar’s effect on crop productivity and the dependence on experimental conditions—A meta-analysis of literature data. Plant Soil 2013, 373, 583–594. [Google Scholar] [CrossRef]
- Ojuederie, O.B.; Babalola, O.O. Microbial and plant-assisted bioremediation of heavy metal polluted environments: A Review. Int. J. Environ. Res. Public Health 2017, 14, 1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dilnashin, H.; Birla, H.; Hoat, T.X.; Singh, H.B.; Singh, S.P.; Keswani, C. Applications of agriculturally important microorganisms for sustainable crop production. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Sharma, V., Salwan, R., Al-Ani, L.K.T., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 403–415. [Google Scholar] [CrossRef]
- Aeron, A.; Kumar, S.; Pandey, P.; Maheshwari, D.K. Emerging role of plant growth promoting rhizobacteria in agrobiology. In Bacteria in Agrobiology: Crop Ecosystems; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1–36. [Google Scholar]
- Gamalero, E.; Glick, B.R. Recent advances in aacterial amelioration of plant drought and salt stress. Biology 2022, 11, 437. [Google Scholar] [CrossRef]
- Zhuang, X.; Chen, J.; Shim, H.; Bai, Z. New advances in plant growth-promoting rhizobacteria for bioremediation. Environ. Int. 2007, 33, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kloepper, J.W.; Ryu, C.-M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef]
- Valls, M.; De Lorenzo, V. Exploiting the genetic and biochemical capacities of bacteria for the remediation of heavy metal pollution. FEMS Microbiol. Rev. 2002, 26, 327–338. [Google Scholar] [CrossRef]
- Guo, J.K.; Ding, Y.Z.; Feng, R.W.; Wang, R.G.; Xu, Y.M.; Chen, C.; Wei, X.L.; Chen, W.M. Burkholderia metalliresistens sp. nov., a multiple metal-resistant and phosphate-solubilising species isolated from heavy metal-polluted soil in Southeast China. Antonie Leeuwenhoek 2015, 107, 1591–1598. [Google Scholar] [CrossRef]
- Alotaibi, F.; Hijri, M.; St-Arnaud, M. Overview of approaches to improve rhizoremediation of petroleum hydrocarbon-contaminated soils. Appl. Microbiol. 2021, 1, 329–351. [Google Scholar] [CrossRef]
- Gurska, J.; Wang, W.; Gerhardt, K.E.; Khalid, A.M.; Isherwood, D.M.; Huang, X.-D.; Glick, B.R.; Greenberg, B.M. Three years field test of a plant growth promoting rhizobacteria inhanced phytoremediation system at a land farm for treatment of hydrocarbon waste. Environ. Sci. Technol. 2009, 43, 4472–4479. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Zhu, Y.; Li, Z.; Huang, B.; Luo, N.; Liu, C.; Zeng, G. Compost as a soil amendment to remediate heavy metal-contaminated agricultural soil: Mechanisms, efficacy, problems, and strategies. Water Air Soil Pollut. 2016, 227, 359. [Google Scholar] [CrossRef]
- Lemmel, F.; Maunoury-Danger, F.; Fanesi, A.; Leyval, C.; Cébron, A. Soil properties and multi-pollution affect taxonomic and functional bacterial diversity in a range of French soils displaying an anthropisation gradient. Microb. Ecol. 2019, 77, 993–1013. [Google Scholar] [CrossRef] [PubMed]
- Auguy, F. Identification de Gènes Impliqués Dans la Tolérance au Plomb Chez la Brassicacée Hirschfeldia incana. Ph.D. Thesis, Université de Montpellier II—Sciences et Techniques du Languedoc, Montpellier, France, 2012; 202p. [Google Scholar]
- Sujkowska-Rybkowska, M.; Banasiewicz, J.; Rekosz-Burlaga, H.; Stępkowski, T. Anthyllis vulneraria and Lotus corniculatus on calamine heaps form nodules with Bradyrhizobium liaoningense-related strains harboring novel in Europe symbiotic nifD haplotypes. Appl. Soil Ecol. 2020, 151, 103539. [Google Scholar] [CrossRef]
- Sbabou, L.; Idir, Y.; Bruneel, O.; Quere, A.L.; Aurag, J.; Bena, G.; Filali-Maltouf, A. Characterization of root-nodule bacteria isolated from Hedysarum spinosissimum L, growing in mining sites of Northeastern region of Morocco. SOJ Microbiol. Infect. Dis. 2016, 4, 1–8. [Google Scholar] [CrossRef]
- Ibañez, S.; Medina, M.I.; Agostini, E. Vicia: A green bridge to clean up polluted environments. Appl. Microbiol. Biotechnol. 2020, 104, 13–21. [Google Scholar] [CrossRef]
- Lamin, H.; Alami, S.; Bouhnik, O.; Bennis, M.; Benkritly, S.; Abdelmoumen, H.; Bedmar, E.J.; Idrissi, M.M.-E. Identification of the endosymbionts from Sulla spinosissima growing in a lead mine tailing in Eastern Morocco as Mesorhizobium camelthorni sv. aridi. J. Appl. Microbiol. 2020, 130, 948–959. [Google Scholar] [CrossRef]
- Oubohssaine, M.; Dahmani, I.; Sbabou, L.; Bruneel, O.; Aurag, J. The rhizosphere of Sulla spinosissima growing in abandoned mining soils is a reservoir of heavy metals tolerant plant growth-promoting rhizobacteria. Biocatal. Agric. Biotechnol. 2022, 39, 1878–8181. [Google Scholar] [CrossRef]
- Becerra-Castro, C.; Prieto-Fernández, A.; Alvarez-Lopez, V.; Monterroso, C.; Cabello-Conejo, M.I.; Acea, M.J.; Kidd, P.S. Nickel solubilizing capacity and characterization of rhizobacteria isolated from hyperaccumulating and non-hyperaccumulating subspecies of Alyssum serpyllifolium. Int. J. Phytoremediat. 2011, 13 (Suppl. S1), 229–244. [Google Scholar] [CrossRef]
- Modi, M.; Shah, K.S.; Modi, V.V. Isolation and characterization of catechol-like siderophore from cowpea Rhizobium RA-1. Arch. Microbiol. 1985, 141, 156–158. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Manjanatha, M.G.; Loynachan, T.E.; Atherly, A.G. Tn5 mutagenesis of chinese Rhizobium fredii for siderophore overproduction. Soil Biol. Biochem. 1992, 24, 151–155. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of phosphorus in soil in connection with the vital activity of some microbial species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Zaidi, S.; Usmani, S.; Singh, B.R.; Musarrat, J. Significance of Bacillus subtilis strain SJ 101 as a bioinoculant for concurrent plant growth promotion and nickel accumulation in Brassica juncea. Chemosphere 2006, 64, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, F.S.; Olsen, S.R. Test of an ascorbic acid method for determining phosphorus in water and NaHCO3 extracts from soil. Soil Sci. Soc. Am. J. 1965, 29, 677–678. [Google Scholar] [CrossRef]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Sheng, X.; He, L.; Wang, Q.; Ye, H.; Jiang, C. Effects of inoculation of biosurfactant-producing Bacillus sp. J119 on plant growth and cadmium uptake in a cadmium-amended soil. J. Hazard. Mater. 2008, 155, 17–22. [Google Scholar] [CrossRef]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Ramakrishna, W. Effect of multiple metal resistant bacteria from contaminated lake sediments on metal accumulation and plant growth. J. Hazard. Mater. 2011, 189, 531–539. [Google Scholar] [CrossRef]
- Döbereiner, J. Isolation and identification of aerobic nitrogen-fixing bacteria from soil and plants. In Methods in Applied Soil Microbiology and Biochemistry; Alef, K., Nannipieri, P., Eds.; Academic Press: London, UK, 1995; pp. 134–141. [Google Scholar]
- Quecine, M.C.; Araújo, W.L.; Rossetto, P.B.; Ferreira, A.; Tsui, S.; Lacava, P.T.; Mondin, M.; Azevedo, J.L.; Pizzirani-Kleiner, A.A. Sugarcane growth promotion by the endophytic bacterium Pantoea agglomerans. Appl. Environ. Microbiol. 2012, 78, 7511–7518. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, D.R.; Arnon, D.I. The Water Culture Method for Growing Plants without Soil; Circular 347; California Agricultural Experiment Station: Berkeley, CA, USA, 1950. [Google Scholar]
- Mackinney, Q. Absorption of light by chlorophyll solutions. J. Biol. Chem. 1941, 140, 315–322. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Chen, G.X.; Asada, K. Ascorbate peroxidase in tea leaves: Occurrence of two isozymes and the differences in their enzymatic and molecular properties. Plant Cell Physiol. 1989, 30, 897–998. [Google Scholar]
- Chance, B.; Maehly, A.C. Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Ahmad, F.; Ahmad, I.; Khan, M.S. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef]
- Wei, G.H.; Fan, L.M.; Zhu, W.F.; Fu, Y.Y.; Yu, J.F.; Tang, M. Isolation and characterization of the heavy metal resistant bacteria CCNWRS33-2 isolated from root nodule of Lespedeza cuneata in gold mine tailings in China. J. Hazard. Mater. 2009, 162, 50–56. [Google Scholar] [CrossRef]
- Sher, S.; Ghani, A.; Sultan, S.; Rehman, A. Bacterial strains isolated from heavy metals contaminated soil and wastewater with potential to oxidize arsenite. Environ. Process. 2021, 8, 333–347. [Google Scholar] [CrossRef]
- Payne, S.M. Detection, isolation, and characterization of siderophores. Methods Enzymol. 1994, 235, 329–344. [Google Scholar]
- Valencia-Cantero, E.; Hernández-Calderón, E.; Velázquez-Becerra, C.; López-Meza, J.E.; Alfaro-Cuevas, R.; López-Bucio, J. Role of dissimilatory fermentative iron-reducing bacteria in Fe uptake by common bean (Phaseolus vulgaris L.) plants grown in alkaline soil. Plant Soil 2007, 291, 263–273. [Google Scholar] [CrossRef]
- Glick, B.R. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Neubauer, U.; Nowack, B.; Furrer, G.; Schulin, R. Heavy metal sorption on clay minerals affected by the siderophore Desferrioxamine B. Environ. Sci. Technol. 2000, 34, 2749–2755. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; Merten, D.; Svatos, A.; Büchel, G.; Kothe, E. Siderophores mediate reduced and increased uptake of cadmium by Streptomyces tendae F4 and sunflower (Helianthus annuus), respectively. J. Appl. Microbiol. 2009, 107, 1687–1696. [Google Scholar] [CrossRef]
- Ji, C.; Juarez-Hernandez, R.E.; Miller, M.J. Exploiting bacterial iron acquisition: Siderophore conjugates. Future Med. Chem. 2012, 4, 297–313. [Google Scholar] [CrossRef]
- Hider, R.C.; Kong, X. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637–657. [Google Scholar] [CrossRef]
- Barua, S.; Tripathi, S.; Chakraborty, A.; Ghosh, S.; Chakrabarti, K. Characterization and crop production efficiency of diazotrophic bacterial isolates from coastal saline soils. Microbiol. Res. 2012, 167, 95–102. [Google Scholar] [CrossRef]
- Garcia de Salamone, I.E.; Funes, J.M.; Di Salvo, L.P.; Escobar-Ortega, J.S.; D’Auria, F.; Ferrando, L.; Fernandez-Scavino, A. Inoculation of paddy rice with Azospirillum brasilense and Pseudomonas fluorescens: Impact of plant genotypes on rhizosphere microbial communities and field crop production. Appl. Soil Ecol. 2012, 61, 196–204. [Google Scholar] [CrossRef]
- Tang, A.; Haruna, A.O.; Majid, N.M.A. Potential PGPR properties of cellulolytic, nitrogen-fixing, and phosphate-solubilizing bacteria of a rehabilitated tropical forest soil. Microorganisms 2020, 8, 442. [Google Scholar] [CrossRef] [Green Version]
- Govindasamy, V.; Senthilkumar, M.; Magheshwaran, V.; Kumar, U.; Bose, P.; Sharma, V.; Annapurna, K. Bacillus and Paenibacillus spp.: Potential PGPR for Sustainable Agriculture. In Plant Growth and Health Promoting Bacteria; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 333–364. [Google Scholar]
- Goswami, D.; Vaghela, H.; Parmar, S.; Dhandhukia, P.; Thakker, J.N. Plant growth promoting potentials of Pseudomonas spp. strain OG isolated from marine water. J. Plant Interact. 2013, 8, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Patten, C.L.; Glick, B.R. Bacterial biosynthesis of indole-3-acetic acid. Can. J. Microbiol. 1996, 42, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Mirza, M.S.; Ahmad, W.; Latif, F.; Haurat, J.; Bally, R.; Normand, P.; Malik, K.A. Isolation, partial characterization, and the effect of plant growth-promoting bacteria (PGPB) on micro-propagated sugarcane in vitro. Plant Soil 2001, 237, 47–54. [Google Scholar] [CrossRef]
- Gravel, V.; Antoun, H.; Tweddell, R.J. Growth stimulation and fruit yield improvement of greenhouse tomato plants by inoculation with Pseudomonas putida or Trichoderma atroviride: Possible role of indole acetic acid (IAA). Soil Biol. Biochem. 2007, 39, 1968–1977. [Google Scholar] [CrossRef]
- Glick, B.R.; Penrose, D.M.; Li, J. A Model for the lowering of plant ethylene concentrations by plant growth-promoting bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Arshad, M.; Hussain, S.; Bhatti, A.S. Perspective of plant growth promoting rhizobacteria (PGPR) containing ACC deaminase in stress agriculture. J. Ind. Microbiol. Biotechnol. 2007, 34, 635–648. [Google Scholar] [CrossRef]
- Onofre-Lemus, J.; Hernandez-Lucas, I.; Girard, L.; Caballero-Mellado, J. ACC (1-aminocyclopropane-1- carboxylate) deaminase activity, a widespread trait in Burkholderia species, and its growth-promoting effect on tomato plants. Appl. Environ. Microbiol. 2009, 75, 6581–6590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahid, M.; Ahmed, T.; Noman, M.; Manzoor, N.; Hussain, S.; Mahmood, F.; Muhammad, S. Microbe-mediated reclamation of contaminated soils: Current status and future perspectives. In Microbial Interventions in Agriculture and Environment: Volume 1: Research Trends, Priorities and Prospects; Singh, D.P., Gupta, V.K., Prabha, R., Eds.; Springer: Singapore, 2019; pp. 261–279. [Google Scholar]
- Barman, D.; Jha, D.K. Metallotolerant microorganisms and microbe-assisted phytoremediation for a sustainable clean environment. In Microbes in Microbial Communities: Ecological and Applied Perspectives; Singh, R.P., Manchanda, G., Bhattacharjee, K., Panosyan, H., Eds.; Springer: Singapore, 2021; pp. 307–336. [Google Scholar]
- Ke, T.; Guo, G.; Liu, J.; Zhang, C.; Tao, Y.; Wang, P.; Xu, Y.; Chen, L. Improvement of the Cu and Cd phytostabilization efficiency of perennial ryegrass through the inoculation of three metal-resistant PGPR strains. Environ. Pollut. 2021, 271, 116314. [Google Scholar] [CrossRef]
- Benidire, L.; Madline, A.; Pereira, S.I.A.; Castro, P.M.L.; Boularbah, A. Synergistic effect of organo-mineral amendments and plant growth-promoting rhizobacteria (PGPR) on the establishment of vegetation cover and amelioration of mine tailings. Chemosphere 2021, 262, 127803. [Google Scholar] [CrossRef]
- Amna; Xia, Y.; Farooq, M.A.; Javed, M.T.; Kamran, M.A.; Mukhtar, T.; Ali, J.; Tabassum, T.; Rehman, S.; Hussain Munis, M.F.; et al. Multi-stress tolerant PGPR Bacillus xiamenensis PM14 activating sugarcane (Saccharum officinarum L.) red rot disease resistance. Plant Physiol. Biochem. 2020, 151, 640–649. [Google Scholar] [CrossRef]
- Amirjani, M. Effects of cadmium on wheat growth and some physiological factors. Int. J. For. Soil Eros. 2012, 2, 50–58. [Google Scholar]
- Zayed, A.M.; Terry, N. Chromium in the environment: Factors affecting biological remediation. Plant Soil 2003, 249, 139–156. [Google Scholar] [CrossRef]
- Khanna, K.; Jamwal, V.L.; Gandhi, S.G.; Ohri, P.; Bhardwaj, R. Metal resistant PGPR lowered Cd uptake and expression of metal transporter genes with improved growth and photosynthetic pigments in Lycopersicon esculentum under metal toxicity. Sci. Rep. 2019, 9, 5855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, D.; Sharma, N.L.; Singh, C.K.; Sarkar, S.K.; Singh, I.; Dotaniya, M.L. Effect of chromium (VI) toxicity on morpho-physiological characteristics, yield, and yield components of two chickpea (Cicer arietinum L.) varieties. PLoS ONE 2020, 15, e0243032. [Google Scholar]
- Hashem, H.A. Cadmium toxicity induces lipid peroxidation and alters cytokinin content and antioxidant enzyme activities in soybean. Botany 2013, 92, 1–7. [Google Scholar] [CrossRef]
- Tripathi, P.; Singh, P.C.; Mishra, A.; Srivastava, S.; Chauhan, R.; Awasthi, S.; Mishra, S.; Dwivedi, S.; Tripathi, P.; Kalra, A.; et al. Arsenic tolerant Trichoderma sp. reduces arsenic induced stress in chickpea (Cicer arietinum). Environ. Pollut. 2017, 223, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Pramanik, K.; Sarkar, A.; Ghosh, P.K.; Soren, T.; Maiti, T.K. Bioaccumulation of cadmium by Enterobacter sp. and enhancement of rice seedling growth under cadmium stress. Ecotoxicol. Environ. Saf. 2018, 156, 183–196. [Google Scholar] [CrossRef]
- Tanaka, A.; Ito, H.; Tanaka, R.; Tanaka, N.K.; Yoshida, K.; Okada, K. Chlorophyll an oxygenase (CAO) is involved in chlorophyll b formation from chlorophyll a. Proc. Natl. Acad. Sci. USA 1998, 95, 12719–12723. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.Y.; Wang, Y.S. Physiological and biochemical responses in the leaves of two mangrove plant seedlings (Kandelia candel and Bruguiera gymnorrhiza) exposed to multiple heavy metals. J. Hazard. Mater. 2010, 182, 848–854. [Google Scholar] [CrossRef]
- Bilal, S.; Shahzad, R.; Imran, M.; Jan, R.; Kim, K.Y.; Lee, I.-J. Synergistic association of endophytic fungi enhances Glycine max L. resilience to combined abiotic stresses: Heavy metals, high temperature and drought stress. Ind. Crop. Prod. 2020, 143, 111931. [Google Scholar] [CrossRef]
- Rahbari, A.; Fatemi, H.; Esmaiel Pour, B.; Rizwan, M.; Soltani, A.-A. Lead (Pb)-resistant bacteria inhibit Pb accumulation in dill (Anethum graveolens L.) by improving biochemical, physiological, and antioxidant enzyme response of plants. Environ. Sci. Pollut. Res. 2021, 28, 5704–5713. [Google Scholar] [CrossRef]
- Demir, S. Influence of arbuscular mycorrhiza on some physiological growth parameters of pepper. Turk. J. Biol. 2005, 28, 85–90. [Google Scholar]
- Singh, R.P.; Mishra, S.; Jha, P.; Raghuvanshi, S.; Jha, P.N. Effect of inoculation of zinc-resistant bacterium Enterobacter ludwigii CDP-14 on growth, biochemical parameters and zinc uptake in wheat (Triticum aestivum L.). Plant. Ecol. Eng. 2018, 116, 163–173. [Google Scholar] [CrossRef]
- Babu, A.G.; Shea, P.J.; Sudhakar, D.; Jung, I.B.; Oh, B.T. Potential use of Pseudomonas koreensis AGB-1 in association with Miscanthus sinensis to remediate heavy metal (loid)-contaminated mining site soil. J. Environ. Manag. 2015, 151, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Almeselmani, M.; Deshmukh, P.S.; Sairam, R.K.; Kushwaha, S.R.; Singh, T.P. Protective role of antioxidant enzymes under high temperature stress. Plant Sci. 2006, 171, 382–388. [Google Scholar] [CrossRef]
- Gupta, D.K.; Palma, J.M.; Corpas, F.J. Generation and scavenging of reactive oxygen species (ROS) in plant cells: An overview. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Sharma, S.S.; Dietz, K.J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2008, 14, 43–50. [Google Scholar] [CrossRef]
- Ju, W.; Liu, L.; Fang, L.; Cui, Y.; Duan, C.; Wu, H. Impact of co-inoculation with plant-growth-promoting rhizobacteria and rhizobium on the biochemical responses of alfalfa-soil system in copper contaminated soil. Ecotoxicol. Environ. Saf. 2019, 167, 218–226. [Google Scholar] [CrossRef]
- Shah, A.A.; Yasin, N.A.; Akram, K.; Ahmad, A.; Khan, W.U.; Akram, W.; Akbar, M. Ameliorative role of Bacillus subtilis FBL-10 and silicon against lead induced stress in Solanum melongena. Plant Physiol. Biochem. 2021, 158, 486–496. [Google Scholar] [CrossRef]
- Hosseini, S.; Kafi, M.; Arghavani, M. The effect of Salicylic acid on physiological characteristics of Lolium grass (Lolium perenne cv. “Numan”) under drought stress. Int. J. Agron. Agric. Res. 2015, 7, 7–14. [Google Scholar]
- Khan, M.M.A.; Haque, E.; Paul, N.C.; Khaleque, M.A.; Al-Garni, S.M.S.; Rahman, M.; Islam, T.F. Enhancement of growth and grain yield of rice in nutrient deficient soils by rice probiotic bacteria. Rice Sci. 2017, 24, 264–273. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Babar, M.A. The root growth of wheat plants, the water conservation and fertility status of sandy soils influenced by plant growth promoting rhizobacteria. Symbiosis 2017, 72, 195–205. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Zandi, P. Effects of exogenously applied plant growth regulators in combination with PGPR on the physiology and root growth of chickpea (Cicer arietinum) and their role in drought tolerance. J. Plant Interact. 2018, 13, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Paredes-Páliz, K.; Rodríguez-Vázquez, R.; Duarte, B.; Caviedes, M.A.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Caçador, M.I.; Rodríguez-Llorente, I.D.; Pajuelo, E. Investigating the mechanisms underlying phytoprotection by plant growth-promoting rhizobacteria in Spartina densiflora under metal stress. Plant Biol. 2018, 20, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Chaturvedi, P.; Chandra, R.; Kumar, S. Identification of heavy metals tolerant Brevundimonas sp. from rhizospheric zone of Saccharum munja L. and their efficacy in in-situ phytoremediation. Chemosphere 2022, 295, 133823. [Google Scholar] [CrossRef] [PubMed]
- Tirry, N.; Kouchou, A.; El Omari, B.; Ferioun, M.; El Ghachtouli, N. Improved chromium tolerance of Medicago sativa by plant growth-promoting rhizobacteria (PGPR). J. Genet. Eng. Biotechnol. 2021, 19, 149. [Google Scholar] [CrossRef]
- Zainab, N.; Amna; Khan, A.A.; Azeem, M.A.; Ali, B.; Wang, T.; Shi, F.; Alghanem, S.M.; Hussain Munis, M.F.; Hashem, M.; et al. PGPR-mediated plant growth attributes and metal extraction ability of Sesbania sesban L. in industrially contaminated soils. Agronomy 2021, 11, 1820. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | LMR249 | LMR250 | LMR266 | LMR283 | LMR291 | LMR340 |
|---|---|---|---|---|---|---|
| Genus/species | Pseudarthrobacter phenanthrenivorans | Pseudarthrobacter oxydans | Stenotrophomonastumulicola | Pseudomonas brassicacearum | Pseudarthrobacter oxydans | Rhodococcus qingshengii |
| Sites | Sidi Boubker | Sidi Boubker | Sidi Boubker | Oued El Heimer | Oued El Heimer | Touissit |
| Origin | Rhizosphere of Stipa tenuissima | Rhizosphere of Stipa tenuissima | Rhizosphere of Sulla spinosissima | Rhizosphere of Sulla spinosissima | Rhizosphere of Sulla spinosissima | Rhizosphere of Sulla spinosissima |
| MIC of Pb (mM) * | 5 | 5 | 10 † | 5 † | 15 † | 25 † |
| MIC of Zn (mM) * | 10 | 5 | 20 † | 10 † | 3 † | 30 † |
| MIC of As (mM) * | 5 | 5 | 10 † | 5 † | 5 † | 35 † |
| P (mg L−1) * | 2.31 ± 0.1 | 0 | 0 | 2.44 ± 0.3 † | 67.63 ± 0.1 † | 0 |
| Quantity (nmol mg−1 h−1) of α cetobutyrate (ACC Activity) * | 128 ± 0.8 | 4 ± 0.5 | 0 | 0 | 71.1 ± 1.4 † | 58 ± 2.3 † |
| Auxin production (µg mL−1) * | 66.66 ± 1.6 | 127 ± 0.1 | 19.5 ± 1 † | 144.98 ± 0.9 † | 134.15 ± 0.6 † | 26.67 ± 1.2 † |
| Siderophore production (%) * | 59.71 ± 2.3 | 97.4 ± 1.4 | 86.77 ± 0.9 † | 75.63 ± 2.4 † | 0 | 0 |
| Nitrogen fixation | - | + | - | - | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oubohssaine, M.; Sbabou, L.; Aurag, J. Native Heavy Metal-Tolerant Plant Growth Promoting Rhizobacteria Improves Sulla spinosissima (L.) Growth in Post-Mining Contaminated Soils. Microorganisms 2022, 10, 838. https://doi.org/10.3390/microorganisms10050838
Oubohssaine M, Sbabou L, Aurag J. Native Heavy Metal-Tolerant Plant Growth Promoting Rhizobacteria Improves Sulla spinosissima (L.) Growth in Post-Mining Contaminated Soils. Microorganisms. 2022; 10(5):838. https://doi.org/10.3390/microorganisms10050838
Chicago/Turabian StyleOubohssaine, Malika, Laila Sbabou, and Jamal Aurag. 2022. "Native Heavy Metal-Tolerant Plant Growth Promoting Rhizobacteria Improves Sulla spinosissima (L.) Growth in Post-Mining Contaminated Soils" Microorganisms 10, no. 5: 838. https://doi.org/10.3390/microorganisms10050838
APA StyleOubohssaine, M., Sbabou, L., & Aurag, J. (2022). Native Heavy Metal-Tolerant Plant Growth Promoting Rhizobacteria Improves Sulla spinosissima (L.) Growth in Post-Mining Contaminated Soils. Microorganisms, 10(5), 838. https://doi.org/10.3390/microorganisms10050838