
In Vitro and In Silico Based Approaches to Identify Potential Novel Bacteriocins from the Athlete Gut Microbiome of an Elite Athlete Cohort

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Subject Recruitment and Sample Collection
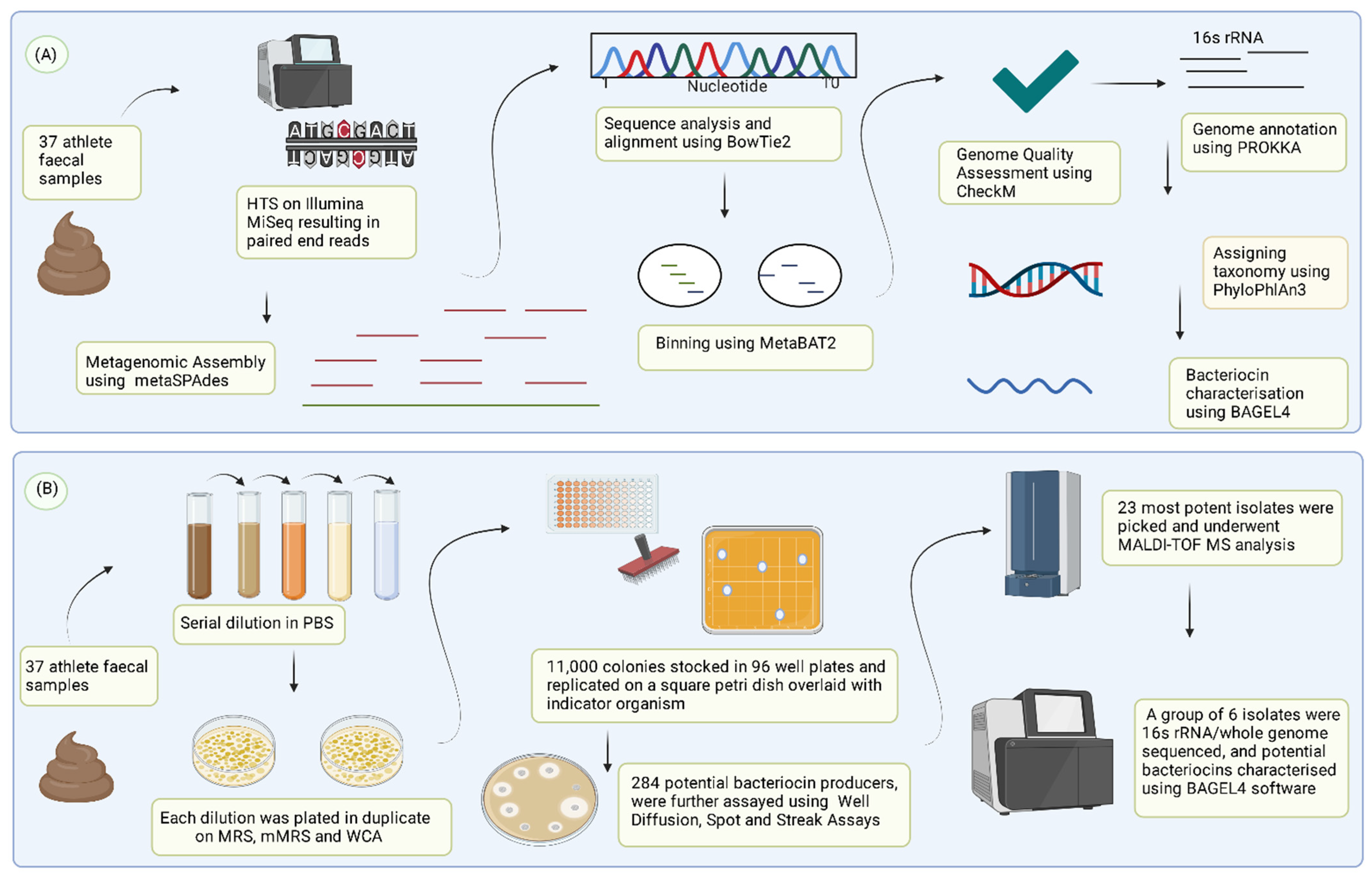
2.2. Recovery of Metagenome Assembled Genomes (MAGs) and Antimicrobial Peptide Production Analysis
2.3. Isolation of Bacterial Isolates Producing Antimicrobial Peptides
2.4. Antimicrobial Activity Assays
2.5. Characterisation of Antimicrobial Activity of Putative Bacteriocin-Producers
2.5.1. Well Diffusion Assay (WDA)
2.5.2. Identification of Putative Bacteriocin-Producing Strains Using 16S rDNA Analysis
2.5.3. Shotgun Whole-Genome Sequencing (WGS) and Analysis
2.5.4. MALDI-TOF Mass Spectrometry
2.6. Targeted Assembly of Metagenome Assembled Genomes
2.7. Statistical Analysis
3. Results
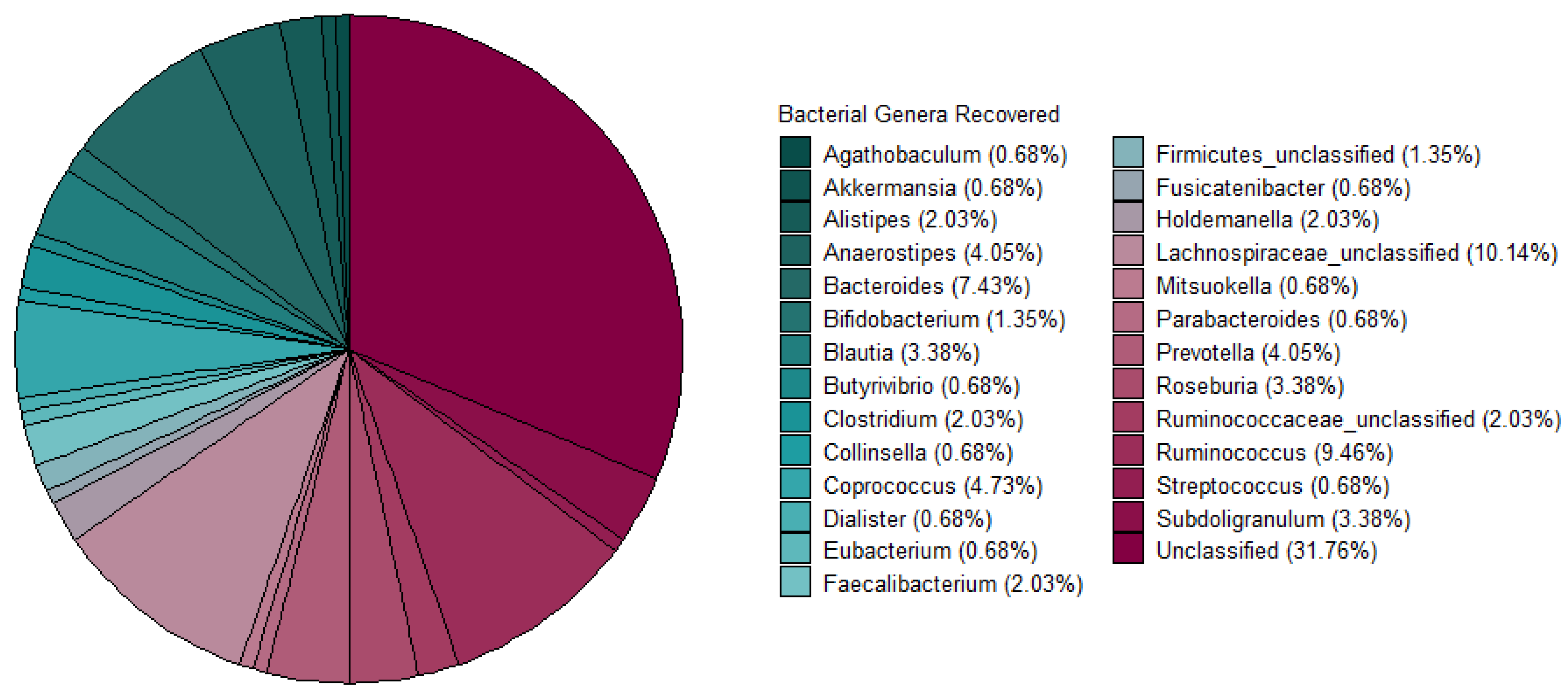
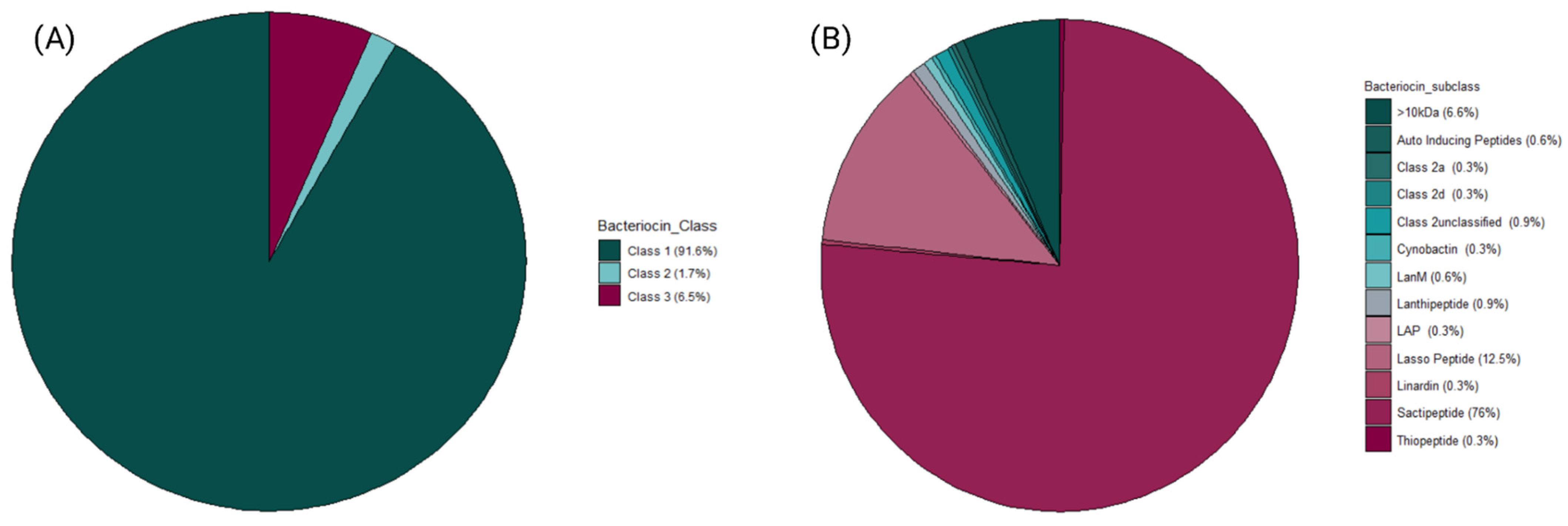
3.1. Assessment and Recovery of Metagenome Assembled Genomes (MAGs) for Bacteriocinogenic Potential
3.2. Detection of Bacteriocin-Producing Bacterial Isolates from Athlete Faecal Samples
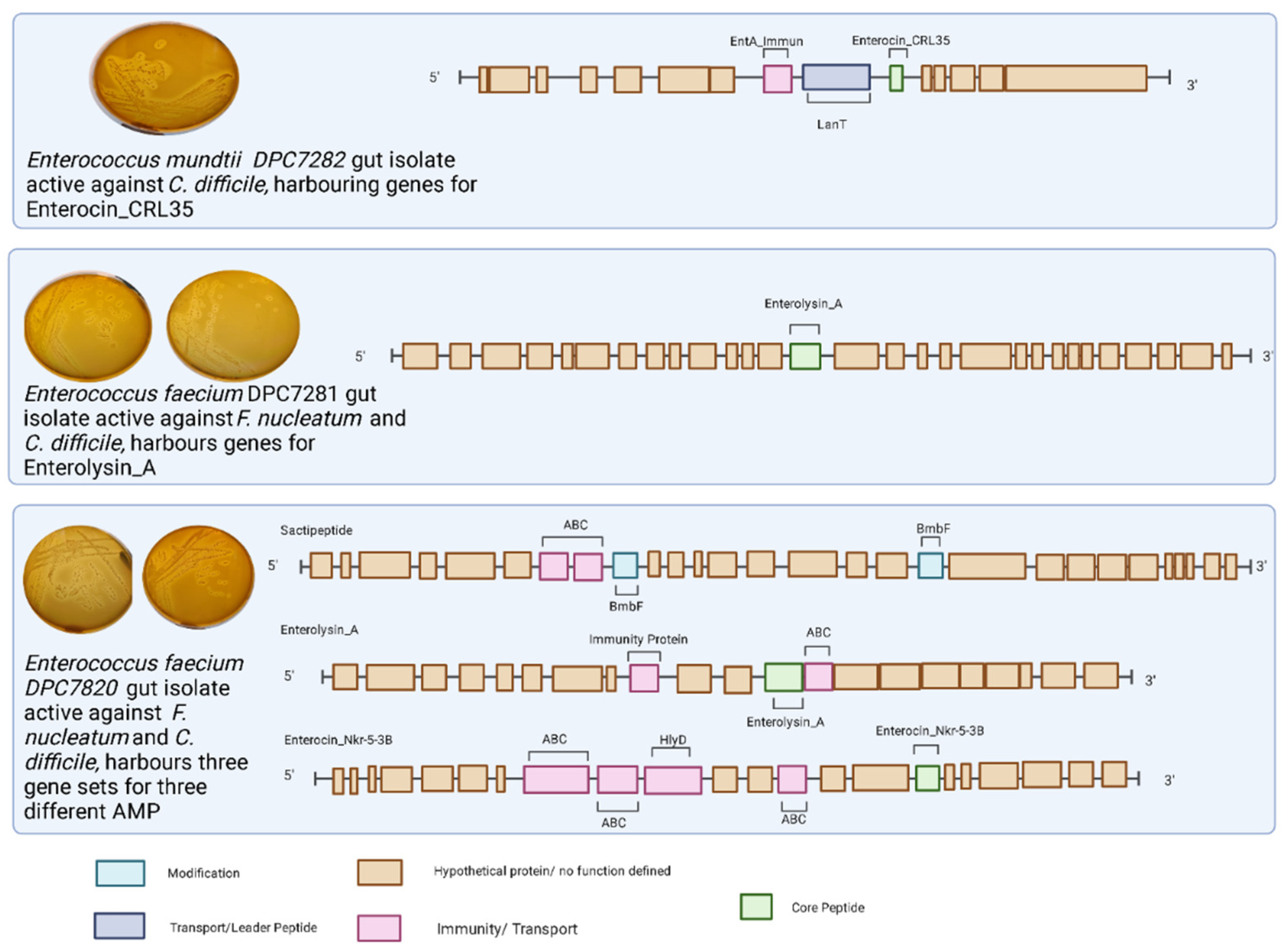
3.3. Identification of Putative Bacteriocin-Producing Strains Isolated from Faecal Samples
3.4. Assessment of Potential Bacteriocin-Producing Strains for Antimicrobial Resistance Genes (ARGs)
3.5. Targeted Assembly of Metagenome Assembled Genomes (MAGs)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brown, D.W.; Heath, G.W.; Balluz, L.; Giles, W.H.; Ford, E.S.; Mokdad, A.H. Associations between Physical Activity Dose and Health-Related Quality of Life. Med. Sci. Sports Exerc. 2004, 36, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Siscovick, D.S.; Fried, L.; Mittelmark, M.; Rutan, G.; Bild, D.; O’Leary, D.H.; Cardiovascular Health Study Research Group. Exercise Intensity and Subclinical Cardiovascular Disease in the Elderly: The Cardiovascular Health Study. Am. J. Epidemiology 1997, 145, 977–986. [Google Scholar] [CrossRef]
- Ostrowski, K.; Rohde, T.; Asp, S.; Schjerling, P.; Pedersen, B.K. Pro- and anti-inflammatory cytokine balance in strenuous exercise in humans. J. Physiol. 1999, 515, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Craft, L.L.; Landers, D.M. The Effect of Exercise on Clinical Depression and Depression Resulting from Mental Illness: A Meta-Analysis. J. Sport Exerc. Psychol. 1998, 20, 339–357. [Google Scholar] [CrossRef]
- Clarke, S.; Murphy, E.F.; O’Sullivan, O.; Lucey, A.; Humphreys, M.; Hogan, A.; Hayes, P.; O’Reilly, M.; Jeffery, I.; Wood-Martin, R.; et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 2014, 63, 1913–1920. [Google Scholar] [CrossRef]
- Cronin, O.; Barton, W.; Skuse, P.; Penney, N.C.; Garcia-Perez, I.; Murphy, E.F.; Woods, T.; Nugent, H.; Fanning, A.; Melgar, S.; et al. A Prospective Metagenomic and Metabolomic Analysis of the Impact of Exercise and/or Whey Protein Supplementation on the Gut Microbiome of Sedentary Adults. mSystems 2018, 3, e00044-18. [Google Scholar] [CrossRef]
- O’Donovan, C.M.; Madigan, S.M.; Garcia-Perez, I.; Rankin, A.; Sullivan, O.O.; Cotter, P. Distinct microbiome composition and metabolome exists across subgroups of elite Irish athletes. J. Sci. Med. Sport 2019, 23, 63–68. [Google Scholar] [CrossRef]
- Petersen, L.M.; Bautista, E.J.; Nguyen, H.; Hanson, B.M.; Chen, L.; Lek, S.H.; Sodergren, E.; Weinstock, G.M. Community characteristics of the gut microbiomes of competitive cyclists. Microbiome 2017, 5, 98. [Google Scholar] [CrossRef]
- Scheiman, J.; Luber, J.M.; Chavkin, T.; Macdonald, T.; Tung, A.; Pham, L.-D.; Wibowo, M.C.; Wurth, R.C.; Punthambaker, S.; Tierney, B.; et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat. Med. 2019, 25, 1104–1109. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Rainey, P.; Rainey, K. Evolution of cooperation and conflict in experimental bacterial populations. Nature 2003, 425, 72–74. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.; Hill, C.; Ross, R. Bacteriocins: Developing innate immunity for food. Nat. Rev. Genet. 2005, 3, 777–788. [Google Scholar] [CrossRef]
- Hatakka, K.; Saxelin, M. Probiotics in Intestinal and Non-Intestinal Infectious Diseases—Clinical Evidence. Curr. Pharm. Des. 2008, 14, 1351–1367. [Google Scholar] [PubMed]
- Rea, M.C.; Dobson, A.; O’Sullivan, O.; Crispie, F.; Fouhy, F.; Cotter, P.D.; Ross, R.P. Effect of broad- and narrow-spectrum antimicrobials on Clostridium difficile and microbial diversity in a model of the distal colon. Proc. Natl. Acad. Sci. USA 2011, 108, 4639–4644. [Google Scholar] [PubMed]
- Walsh, M.C.; Gardiner, G.E.; Hart, O.; Lawlor, P.G.; Daly, M.; Lynch, B.; Richert, B.T.; Radcliffe, S.; Giblin, L.; Hill, C.; et al. Predominance of a bacteriocin-producing Lactobacillus salivarius component of a five-strain probiotic in the porcine ileum and effects on host immune phenotype. FEMS Microbiol. Ecol. 2008, 64, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Corr, S.C.; Li, Y.; Riedel, C.U.; O’Toole, P.W.; Hill, C.; Gahan, C.G.M. Bacteriocin production as a mechanism for the antiinfective activity of Lactobacillus salivarius UCC118. Proc. Natl. Acad. Sci. USA 2007, 104, 7617–7621. [Google Scholar] [CrossRef] [PubMed]
- Van Hemert, S.; Meijerink, M.; Molenaar, D.; Bron, P.A.; De Vos, P.; Kleerebezem, M.; Marco, M.L. Identification of Lactobacillus plantarum genes modulating the cytokine response of human peripheral blood mononuclear cells. BMC Microbiol. 2010, 10, 293. [Google Scholar]
- Deng, Z.; Luo, X.M.; Liu, J.; Wang, H. Quorum Sensing, Biofilm, and Intestinal Mucosal Barrier: Involvement the Role of Probiotic. Front. Cell. Infect. Microbiol. 2020, 10, 504. [Google Scholar] [CrossRef]
- Khalaf, H.; Nakka, S.S.; Sandén, C.; Svärd, A.; Hultenby, K.; Scherbak, N.; Aili, D.; Bengtsson, T. Antibacterial effects of Lactobacillus and bacteriocin PLNC8 αβ on the periodontal pathogen Porphyromonas gingivalis. BMC Microbiol. 2016, 16, 188. [Google Scholar] [CrossRef]
- Ovchinnikov, K.V.; Kranjec, C.; Thorstensen, T.; Carlsen, H.; Diep, D.B. Successful Development of Bacteriocins into Therapeutic Formulation for Treatment of MRSA Skin Infection in a Murine Model. Antimicrob. Agents Chemother. 2020, 64, e00829-20. [Google Scholar] [CrossRef]
- Nascimento, J.S.; Ceotto, H.; Nascimento, S.B.; Giambiagi-Demarval, M.; Santos, K.R.N.; Bastos, M.C.F. Bacteriocins as alternative agents for control of multiresistant staphylococcal strains. Lett. Appl. Microbiol. 2006, 42, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Mathiesen, G.; Huehne, K.; Kroeckel, L.; Axelsson, L.; Eijsink, V.G. Characterization of a New Bacteriocin Operon in Sakacin P-Producing Lactobacillus sakei, Showing Strong Translational Coupling between the Bacteriocin and Immunity Genes. Appl. Environ. Microbiol. 2005, 71, 3565–3574. [Google Scholar] [PubMed]
- Nes, I.F.; Diep, D.B.; Brurberg, M.B.; Eijsink, V.; Holo, H. Biosynthesis of bacteriocins in lactic acid bacteria. Antonie van Leeuwenhoek 1996, 70, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Drissi, F.; Buffet, S.; Raoult, D.; Merhej, V. Common occurrence of antibacterial agents in human intestinal microbiota. Front. Microbiol. 2015, 6, 441. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.J.; Guinane, C.; Hill, C.; Ross, R.P.; O’Toole, P.W.; Cotter, P.D. In silico identification of bacteriocin gene clusters in the gastrointestinal tract, based on the Human Microbiome Project’s reference genome database. BMC Microbiol. 2015, 15, 1–11. [Google Scholar] [CrossRef]
- Zheng, J.; Gänzle, M.; Lin, X.B.; Ruan, L.; Sun, M. Diversity and dynamics of bacteriocins from human microbiome. Environ. Microbiol. 2014, 17, 2133–2143. [Google Scholar] [CrossRef]
- Kommineni, S.; Bretl, D.J.; Lam, V.; Chakraborty, R.; Hayward, M.; Simpson, P.M.; Cao, Y.; Bousounis, P.; Kristich, C.J.; Salzman, N.H. Bacteriocin production augments niche competition by enterococci in the mammalian gastrointestinal tract. Nature 2015, 526, 719–722. [Google Scholar] [CrossRef]
- Rea, M.; Clayton, E.; O’Connor, P.M.; Shanahan, F.; Kiely, B.; Ross, R.; Hill, C. Antimicrobial activity of lacticin 3147 against clinical Clostridium difficile strains. J. Med Microbiol. 2007, 56, 940–946. [Google Scholar] [CrossRef]
- Casey, P.; Casey, G.; Gardiner, G.; Tangney, M.; Stanton, C.; Ross, R.; Hill, C.; Fitzgerald, G. Isolation and characterization of anti-Salmonella lactic acid bacteria from the porcine gastrointestinal tract. Lett. Appl. Microbiol. 2004, 39, 431–438. [Google Scholar] [CrossRef]
- Reenen, V.; Dicks, L.M.; Chikindas, M.L. Isolation, purification and partial characterization of plantaricin 423, a bacteriocin produced by Lactobacillus plantarum. J. Appl. Microbiol. 1998, 84, 1131–1137. [Google Scholar] [CrossRef]
- Kim, T.-S.; Hur, J.-W.; Yu, M.-A.; Cheigh, C.-I.; Kim, K.-N.; Hwang, J.-K.; Pyun, Y.-R. Antagonism of Helicobacter pylori by Bacteriocins of Lactic Acid Bacteria. J. Food Prot. 2003, 66, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Soria, M.C.; Audisio, M.C. Inhibition of Bacillus cereus Strains by Antimicrobial Metabolites from Lactobacillus johnsonii CRL1647 and Enterococcus faecium SM21. Probiotics Antimicrob. Proteins 2014, 6, 208–216. [Google Scholar] [PubMed]
- Chaveerach, P.; Lipman, L.; van Knapen, F. Antagonistic activities of several bacteria on in vitro growth of 10 strains of Campylobacter jejuni/coli. Int. J. Food Microbiol. 2003, 90, 43–50. [Google Scholar] [CrossRef]
- Vedantam, G.; Clark, A.; Chu, M.; McQuade, R.; Mallozzi, M.; Viswanathan, V.K. Clostridium difficile infection: Toxins and non-toxin virulence factors, and their contributions to disease establishment and host response. Gut Microbes 2012, 3, 121–134. [Google Scholar]
- George, W.L.; Goldstein, E.; Sutter, V.; Ludwig, S.; Finegold, S. ÆTIOLOGY OF ANTIMICROBIAL-AGENT-ASSOCIATED COLITIS. Lancet 1978, 311, 802–803. [Google Scholar] [CrossRef]
- Lim, S.; Knight, D.; Riley, T. Clostridium difficile and One Health. Clin. Microbiol. Infect. 2019, 26, 857–863. [Google Scholar] [CrossRef]
- Pépin, J.; Valiquette, L.; Alary, M.-E.; Villemure, P.; Pelletier, A.; Forget, K.; Pépin, K.; Chouinard, D. Clostridium difficile-associated diarrhea in a region of Quebec from 1991 to 2003: A changing pattern of disease severity. Can. Med. Assoc. J. 2004, 171, 466–472. [Google Scholar] [CrossRef]
- Starr, J. Clostridium difficile associated diarrhoea: Diagnosis and treatment. BMJ 2005, 331, 498–501. [Google Scholar] [CrossRef][Green Version]
- Han, Y.W. Fusobacterium nucleatum: A commensal-turned pathogen. Curr. Opin. Microbiol. 2015, 23, 141–147. [Google Scholar] [CrossRef]
- Li, X.; Huang, J.; Yu, T.; Fang, X.; Lou, L.; Xin, S.; Ji, L.; Jiang, F.; Lou, Y. Fusobacterium nucleatum Promotes the Progression of Colorectal Cancer Through Cdk5-Activated Wnt/β-Catenin Signaling. Front. Microbiol. 2021, 11, 3231. [Google Scholar] [CrossRef]
- Rotmistrovsky, K.; Agarwala, R. BMTagger: Best Match Tagger for Removing Human Reads from Metagenomics Datasets. 2011. Available online: ftp://ftp.ncbi.nlm.nih.gov/pub/agarwala/bmtagger/ (accessed on 2 June 2021).
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Nurk, S.; Meleshko, D.; Korobeynikov, A.; Pevzner, P.A. metaSPAdes: A new versatile metagenomic assembler. Genome Res. 2017, 27, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.D.; Li, F.; Kirton, E.; Thomas, A.; Egan, R.; An, H.; Wang, Z. MetaBAT 2: An adaptive binning algorithm for robust and efficient genome reconstruction from metagenome assemblies. PeerJ 2019, 7, e7359. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef]
- Bowers, R.M.; Kyrpides, N.C.; Stepanauskas, R.; Harmon-Smith, M.; Doud, D.; Reddy, T.B.K.; Schulz, F.; Jarett, J.; Rivers, A.R.; Eloe-Fadrosh, E.A.; et al. Minimum information about a single amplified genome (MISAG) and a metagenome-assembled genome (MIMAG) of bacteria and archaea. Nat. Biotechnol. 2017, 35, 725–731. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Asnicar, F.; Thomas, A.M.; Beghini, F.; Mengoni, C.; Manara, S.; Manghi, P.; Zhu, Q.; Bolzan, M.; Cumbo, F.; May, U.; et al. Precise phylogenetic analysis of microbial isolates and genomes from metagenomes using PhyloPhlAn 3.0. Nat. Commun. 2020, 11, 2500. [Google Scholar] [CrossRef]
- van Heel, A.J.; de Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar]
- Ryan, M.P.; Rea, M.C.; Hill, C.; Ross, R.P. An application in cheddar cheese manufacture for a strain of Lactococcus lactis producing a novel broad-spectrum bacteriocin, lacticin 3147. Appl. Environ. Microbiol. 1996, 62, 612–619. [Google Scholar]
- Hammami, R.; Zouhir, A.; Le Lay, C.; Ben Hamida, J.; Fliss, I. BACTIBASE second release: A database and tool platform for bacteriocin characterization. BMC Microbiol. 2010, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Pevzner, P.A. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. J. Comput. Mol. Cell Biol. 2012, 19, 455–477. [Google Scholar]
- Chaumeil, P.-A.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk: A toolkit to classify genomes with the Genome Taxonomy Database. Bioinformatics 2020, 36, 1925–1927. [Google Scholar] [CrossRef]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L.; et al. The Comprehensive Antibiotic Resistance Database. Antimicrob. Agents Chemother. 2013, 57, 3348–3357. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.; Igrejas, G.; Gonçalves, A.; Poeta, P. Commensal gut bacteria: Distribution of Enterococcus species and prevalence of Escherichia coli phylogenetic groups in animals and humans in Portugal. Ann. Microbiol. 2011, 62, 449–459. [Google Scholar] [CrossRef]
- Barton, W.; Penney, N.C.; Cronin, O.; Garcia-Perez, I.; Molloy, M.G.; Holmes, E.; Shanahan, F.; Cotter, P.D.; O’Sullivan, O. The microbiome of professional athletes differs from that of more sedentary subjects in composition and particularly at the functional metabolic level. Gut 2018, 67, 625–633. [Google Scholar] [PubMed]
- Rosberg-Cody, E.; Ross, R.; Hussey, S.; Ryan, C.A.; Murphy, B.P.; Fitzgerald, G.F.; Devery, R.; Stanton, C. Mining the Microbiota of the Neonatal Gastrointestinal Tract for Conjugated Linoleic Acid-Producing Bifidobacteria. Appl. Environ. Microbiol. 2004, 70, 4635–4641. [Google Scholar] [CrossRef]
- Wall, R.; Hussey, S.G.; Ryan, C.A.; O’Neill, M.; Fitzgerald, G.; Stanton, C.; Ross, R.P. Presence of two Lactobacillus and Bifidobacterium probiotic strains in the neonatal ileum. ISME J. 2007, 2, 83–91. [Google Scholar] [CrossRef]
- Jost, T.; Lacroix, C.; Braegger, C.P.; Chassard, C. New insights in gut microbiota establishment in healthy breast fed neonates. PLoS ONE 2012, 7, e44595. [Google Scholar]
- Brazier, J.S.; Goldstein, E.J.; Citron, D.M.; Ostovari, M.I. Fastidious anaerobe agar compared with Wilkins-Chalgren agar, brain heart infusion agar, and brucella agar for susceptibility testing of Fusobacterium species. Antimicrob. Agents Chemother. 1990, 34, 2280–2282. [Google Scholar] [CrossRef]
- Nilsen, T.; Nes, I.F.; Holo, H. Enterolysin A, a cell wall-degrading bacteriocin from Enterococcus faecalis LMG 2333. Appl. Environ. Microbiol. 2003, 69, 2975–2984. [Google Scholar]
- Jang, L.-G.; Choi, G.; Kim, S.-W.; Kim, B.-Y.; Lee, S.; Park, H. The combination of sport and sport-specific diet is associated with characteristics of gut microbiota: An observational study. J. Int. Soc. Sports Nutr. 2019, 16, 21. [Google Scholar] [CrossRef]
- Jaago, M.; Timmusk, U.S.; Timmusk, T.; Palm, K. Drastic Effects on the Microbiome of a Young Rower Engaged in High-Endurance Exercise After a Month Usage of a Dietary Fiber Supplement. Front. Nutr. 2021, 8, 654008. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.B.; Marques, C.; Abreu, R.; Figueiredo, P.; Calhau, C.; Brito, J.; Sousa, M. Gut microbiota of elite female football players is not altered during an official international tournament. Scand. J. Med. Sci. Sports 2021. published ahead of print. [Google Scholar] [CrossRef]
- Bressa, C.; Bailén-Andrino, M.; Pérez-Santiago, J.; González-Soltero, R.; Pérez, M.; Montalvo-Lominchar, M.G.; Maté-Muñoz, J.L.; Domínguez, R.; Moreno, D.; Larrosa, M. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE 2017, 12, e0171352. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, Z.; Hu, B.; Huang, W.; Yuan, C.; Zou, L. Response of Gut Microbiota to Metabolite Changes Induced by Endurance Exercise. Front. Microbiol. 2018, 9, 765. [Google Scholar] [CrossRef]
- Queipo-Ortuño, M.I.; Seoane, L.M.; Murri, M.; Pardo, M.; Gomez-Zumaquero, J.M.; Cardona, F.; Casanueva, F.; Tinahones, F.J. Gut Microbiota Composition in Male Rat Models under Different Nutritional Status and Physical Activity and Its Association with Serum Leptin and Ghrelin Levels. PLoS ONE 2013, 8, e65465. [Google Scholar] [CrossRef]
- Lakshminarayanan, B.; Guinane, C.; O’Connor, P.; Coakley, M.; Hill, C.; Stanton, C.; O’Toole, P.; Ross, R. Isolation and characterization of bacteriocin-producing bacteria from the intestinal microbiota of elderly Irish subjects. J. Appl. Microbiol. 2012, 114, 886–898. [Google Scholar] [CrossRef]
- O’Sullivan, J.N.; Rea, M.C.; O’Connor, P.M.; Hill, C.; Ross, R.P. Human skin microbiota is a rich source of bacteriocin-producing staphylococci that kill human pathogens. FEMS Microbiol. Ecol. 2018, 95, fiy241. [Google Scholar] [CrossRef]
- Khattab, R.A.; Ahmed, N.A.; Ragab, Y.M.; Rasmy, S.A. Bacteria producing antimicrobials against Clostridium difficile isolated from human stool. Anaerobe 2020, 63, 102206. [Google Scholar] [CrossRef]
- Basanta, A.; Sánchez, J.; Gómez-Sala, B.; Herranz, C.; Hernández, P.E.; Cintas, L.M. Antimicrobial activity of Enterococcus faecium L50, a strain producing enterocins L50 (L50A and L50B), P and Q, against beer-spoilage lactic acid bacteria in broth, wort (hopped and unhopped), and alcoholic and non-alcoholic lager beers. Int. J. Food Microbiol. 2008, 125, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Dezwaan, D.C.; Mequio, M.J.; Littell, J.S.; Allen, J.P.; Rossbach, S.; Pybus, V. Purification and characterization of enterocin 62-6, a two-peptide bacteriocin produced by a vaginal strain of Enterococcus faecium: Potential significance in bacterial vaginosis. Microb. Ecol. Health Dis. 2007, 19, 241–250. [Google Scholar] [PubMed]
- Umu, Ö.C.O.; Bäuerl, C.; Oostindjer, M.; Pope, P.B.; Hernández, P.E.; Pérez-Martínez, G.; Diep, D.B. The Potential of Class II Bacteriocins to Modify Gut Microbiota to Improve Host Health. PLoS ONE 2016, 11, e0164036. [Google Scholar] [CrossRef]
- Minahk, C.; Farãas, M.E.; Sesma, F.; Morero, R.D. Effect of Enterocin CRL35 on Listeria monocytogenes cell membrane. FEMS Microbiol. Lett. 2000, 192, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Todorov, S.D.; Kang, H.-J.; Ivanova, I.V.; Holzapfel, W.H. Bacteriocins From LAB and Other Alternative Approaches for the Control of Clostridium and Clostridiodes Related Gastrointestinal Colitis. Front. Bioeng. Biotechnol. 2020, 8, 1088. [Google Scholar] [CrossRef]
- Lawrence, G.W.; Begley, M.; Cotter, P.D.; Guinane, C.M. Potential Use of Biotherapeutic Bacteria to Target Colorectal Cancer-Associated Taxa. Int. J. Mol. Sci. 2020, 21, 924. [Google Scholar] [CrossRef]
- Gao, Z.; Guo, B.; Gao, R.; Zhu, Q.; Wu, W.; Qin, H. Probiotics modify human intestinal mucosa-associated microbiota in patients with colorectal cancer. Mol. Med. Rep. 2015, 12, 6119–6127. [Google Scholar] [CrossRef]
- Hickey, R.M.; Twomey, D.P.; Ross, R.P.; Hill, C. Production of enterolysin A by a raw milk enterococcal isolate exhibiting multiple virulence factors. Microbiology 2003, 149, 655–664. [Google Scholar] [CrossRef][Green Version]
- Rea, M.C.; Sit, C.S.; Clayton, E.; O’Connor, P.M.; Whittal, R.M.; Zheng, J.; Vederas, J.C.; Ross, R.P.; Hill, C. Thuricin CD, a posttranslationally modified bacteriocin with a narrow spectrum of activity against Clostridium difficile. Proc. Natl. Acad. Sci. USA 2010, 107, 9352–9357. [Google Scholar] [CrossRef]
- Himeno, K.; Rosengren, K.J.; Inoue, T.; Perez, R.H.; Colgrave, M.L.; Lee, H.S.; Chan, L.Y.; Henriques, S.T.; Fujita, K.; Ishibashi, N.; et al. Identification, Characterization, and Three-Dimensional Structure of the Novel Circular Bacteriocin, Enterocin NKR-5-3B, from Enterococcus faecium. Biochemistry 2015, 54, 4863–4876. [Google Scholar] [CrossRef]
- McKay, A.M. Antimicrobial activity of Enterococcus faecium against Listeria spp. Lett. Appl. Microbiol. 1990, 11, 15–17. [Google Scholar] [CrossRef]
- Siragusa, G.R. Production of bacteriocin inhibitory to Listeria species by Enterococcus hirae. Appl. Environ. Microbiol. 1992, 58, 3508–3513. [Google Scholar] [CrossRef] [PubMed]
- Parente, E.; Hill, C. Inhibition of Listeria in Buffer, Broth, and Milk by Enterocin 1146, a Bacteriocin Produced by Enterococcus faecium. J. Food Prot. 1992, 55, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Vlaemynck, G.; Herman, L.; Coudijzer, K. Isolation and characterization of two bacteriocins produced by Enterococcus faecium strains inhibitory to Listeria monocytogenes. Int. J. Food Microbiol. 1994, 24, 211–225. [Google Scholar] [CrossRef]
- Hammerum, A.M. Enterococci of animal origin and their significance for public health. Clin. Microbiol. Infect. 2012, 18, 619–625. [Google Scholar] [CrossRef]
- Abriouel, H.; Ben Omar, N.; Molinos, A.C.; López, R.L.; Grande, M.J.; Martínez-Viedma, P.; Ortega, E.; Cañamero, M.M.M.; Galvez, A. Comparative analysis of genetic diversity and incidence of virulence factors and antibiotic resistance among enterococcal populations from raw fruit and vegetable foods, water and soil, and clinical samples. Int. J. Food Microbiol. 2008, 123, 38–49. [Google Scholar] [CrossRef]
- Nawaz, F.; Khan, M.N.; Javed, A.; Ahmed, I.; Ali, N.; Ali, M.I.; Bakhtiar, S.M.; Imran, M. Genomic and Functional Characterization of Enterococcus mundtii QAUEM2808, Isolated From Artisanal Fermented Milk Product Dahi. Front. Microbiol. 2019, 10, 434. [Google Scholar] [CrossRef]
- Foka, F.E.T.; Ateba, C.N. Detection of Virulence Genes in Multidrug Resistant Enterococci Isolated from Feedlots Dairy and Beef Cattle: Implications for Human Health and Food Safety. BioMed Res. Int. 2019, 2019, 5921840. [Google Scholar] [CrossRef]
- Tian, Y.; Yu, H.; Wang, Z. Distribution of acquired antibiotic resistance genes among Enterococcus spp. isolated from a hospital in Baotou, China. BMC Res. Notes 2019, 12, 27. [Google Scholar] [CrossRef]
- Portillo, A.; Larrea, F.R.; Zarazaga, M.; Alonso, A.; Martinez, J.L.; Torres, C. Macrolide Resistance Genes in Enterococcus spp. Antimicrob. Agents Chemother. 2000, 44, 967–971. [Google Scholar] [CrossRef]
- Jensen, L.B.; Frimodt-Møller, N.; Aarestrup, F. Presence of erm gene classes in Gram-positive bacteria of animal and human origin in Denmark. FEMS Microbiol. Lett. 1999, 170, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Trieu-Cuot, P.; de Cespédès, G.; Bentorcha, F.; Delbos, F.; Gaspar, E.; Horaud, T. Study of heterogeneity of chloramphenicol acetyltransferase (CAT) genes in streptococci and enterococci by polymerase chain reaction: Characterization of a new CAT determinant. Antimicrob. Agents Chemother. 1993, 37, 2593–2598. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Werner, G.; Hildebrandt, B.; Witte, W. Aminoglycoside-Streptothricin Resistance Gene Cluster aadE–sat4–aphA-3 Disseminated among Multiresistant Isolates of Enterococcus faecium. Antimicrob. Agents Chemother. 2001, 45, 3267–3269. [Google Scholar] [CrossRef] [PubMed]
- Diarra, M.S.; Rempel, H.; Champagne, J.; Masson, L.; Pritchard, J.; Topp, E. Distribution of Antimicrobial Resistance and Virulence Genes in Enterococcus spp. and Characterization of Isolates from Broiler Chickens. Appl. Environ. Microbiol. 2010, 76, 8033–8043. [Google Scholar] [PubMed]
- Woegerbauer, M.; Zeinzinger, J.; Springer, B.; Hufnagl, P.; Indra, A.; Korschineck, I.; Allerberger, F. Prevalence of the aminoglycoside phosphotransferase genes aph(3′)-IIIa and aph(3′)-IIa in Escherichia coli, Enterococcus faecalis, Enterococcus faecium, Pseudomonas aeruginosa, Salmonella enterica subsp. enterica and Staphylococcus aureus isolates in Austria. J. Med. Microbiol. 2014, 63, 210–217. [Google Scholar]
- Coque, T.M.; Singh, K.V.; Weinstock, G.; Murray, B.E. Characterization of Dihydrofolate Reductase Genes from Trimethoprim-Susceptible and Trimethoprim-Resistant Strains of Enterococcus faecalis. Antimicrob. Agents Chemother. 1999, 43, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, H.; Ortega-Polo, R.; Zaheer, R.; Goji, N.; Amoako, K.K.; Brown, R.S.; Majury, A.; Liss, S.N.; McAllister, T.A. Comparative genomics of multidrug-resistant Enterococcus spp. isolated from wastewater treatment plants. BMC Microbiol. 2020, 20, 20. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, Y.; Pan, B.; Hao, P.; Li, Y.; Shao, Z.; Xu, X.; Li, X. Optimizing hybrid assembly of next-generation sequence data from Enterococcus faecium: A microbe with highly divergent genome. BMC Syst. Biol. 2012, 6, S21. [Google Scholar] [CrossRef]
- Zhong, Z.; Kwok, L.-Y.; Hou, Q.; Sun, Y.; Li, W.; Zhang, H.; Sun, Z. Comparative genomic analysis revealed great plasticity and environmental adaptation of the genomes of Enterococcus faecium. BMC Genom. 2019, 20, 602. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Media for Cultivation | Growth Atmosphere | Temperature (°C) |
|---|---|---|---|
| Lactobacillus bulgaricus LMG6901 * | MRS | Anaerobic ****** | 37 |
| Listeria innocua DPC3572 ** | BHI *** | Aerobic | 37 |
| Fusobacterium nucleatum DPC6999 | FAA/WCA **** | Anaerobic | 37 |
| Clostridioides difficile DPC6509 | RCA/BHI ***** | Anaerobic | 37 |
| Indicator Organism | Number of Isolates Screened | Isolates with Antagonistic Activity against Indicator | Frequency of Isolation % |
|---|---|---|---|
| Listeria innocua DPC3572 | 5000 | 2 | 0.04% |
| Lactobacillus bulgaricus LMG6901 | 5000 | 1 | 0.02% |
| Fusobacterium nucleatum DPC6999 | 6000 | 136 | 2.26% |
| Clostridioides difficile DPC6509 | 6000 | 145 | 2.42% |
| Strain Designation | Media | Indicator | Taxonomy Method | Genus/Species | Method of Bacteriocin Prediction | Bacteriocin Predicted | AMR Genes Identified |
|---|---|---|---|---|---|---|---|
| LW001 | mMRS × 2 [Mupirocin] | Listeria innocua | 16s rRNA | Enterococcus | MALDI-TOF | Enterocin Q | n/a |
| LW002 | mMRS × 2 [Mupirocin] | Listeria innocua | 16s rRNA | Enterococcus | MALDI-TOF | Enterocin Q | n/a |
| LW003 | mMRS × 2 [Mupirocin] | Lactobacillus bulgaricus | 16s rRNA | Enterococcus | MALDI-TOF | Enterocin 62-6 | n/a |
| DPC7281 | WCA | Fusobacterium nucleatum | WGS | Enterococcus faecalis | WGS + BAGEL4 | Enterolysin A | dfrE, efrA, tetM, ermB, E. faecalis chloramphenicol acetyltransferase, aad(6), SAT-4, APH(3′)-IIIa |
| DPC7280 | WCA | Fusobacterium nucleatum | WGS | Enterococcus faecalis | WGS + BAGEL4 | Enterolysin A Enterocin NKR-5-3B Sactipeptide | dfrE, efrA |
| DPC7282 | WCA | Clostridioides difficile | WGS | Enterococcus mundtii | WGS + BAGEL4 | Enterocin CRL35 | No AMR genes identified |
| Species | Total Number of BLAST Hits |
|---|---|
| Enterococcus mundtii | 2,732,299 |
| Enterococcus hirae | 13,166,169 |
| Enterococcus durans | 9,623,516 |
| Enterococcus faecalis | 73,372,776 |
| Enterococcus faecium | 670,879,682 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wosinska, L.; Walsh, C.J.; O’Connor, P.M.; Lawton, E.M.; Cotter, P.D.; Guinane, C.M.; O’Sullivan, O. In Vitro and In Silico Based Approaches to Identify Potential Novel Bacteriocins from the Athlete Gut Microbiome of an Elite Athlete Cohort. Microorganisms 2022, 10, 701. https://doi.org/10.3390/microorganisms10040701
Wosinska L, Walsh CJ, O’Connor PM, Lawton EM, Cotter PD, Guinane CM, O’Sullivan O. In Vitro and In Silico Based Approaches to Identify Potential Novel Bacteriocins from the Athlete Gut Microbiome of an Elite Athlete Cohort. Microorganisms. 2022; 10(4):701. https://doi.org/10.3390/microorganisms10040701
Chicago/Turabian StyleWosinska, Laura, Calum J. Walsh, Paula M. O’Connor, Elaine M. Lawton, Paul D. Cotter, Caitriona M. Guinane, and Orla O’Sullivan. 2022. "In Vitro and In Silico Based Approaches to Identify Potential Novel Bacteriocins from the Athlete Gut Microbiome of an Elite Athlete Cohort" Microorganisms 10, no. 4: 701. https://doi.org/10.3390/microorganisms10040701
APA StyleWosinska, L., Walsh, C. J., O’Connor, P. M., Lawton, E. M., Cotter, P. D., Guinane, C. M., & O’Sullivan, O. (2022). In Vitro and In Silico Based Approaches to Identify Potential Novel Bacteriocins from the Athlete Gut Microbiome of an Elite Athlete Cohort. Microorganisms, 10(4), 701. https://doi.org/10.3390/microorganisms10040701