A Novel Subunit Vaccine Based on Outer Capsid Proteins of Grass Carp Reovirus (GCRV) Provides Protective Immunity against GCRV Infection in Rare Minnow (Gobiocypris rarus)

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
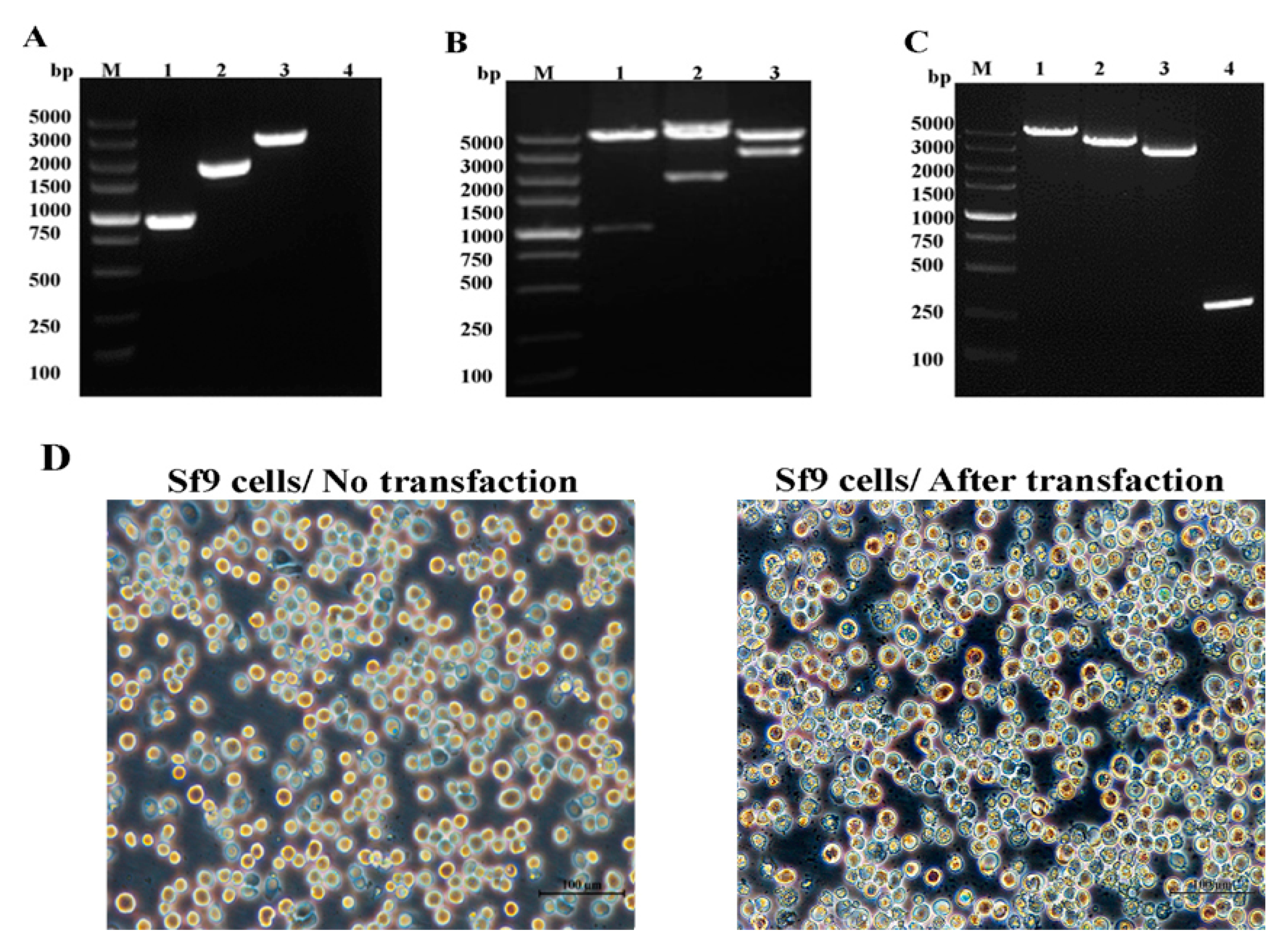
2.1. VP35 and VP4 Gene Cloning and Identification of Donor Vectors pFastBac-VP35, pFastBac-VP4, and pFastBac-VP35-T2A-VP4
2.2. Identification of Recombinant Baculoviruses
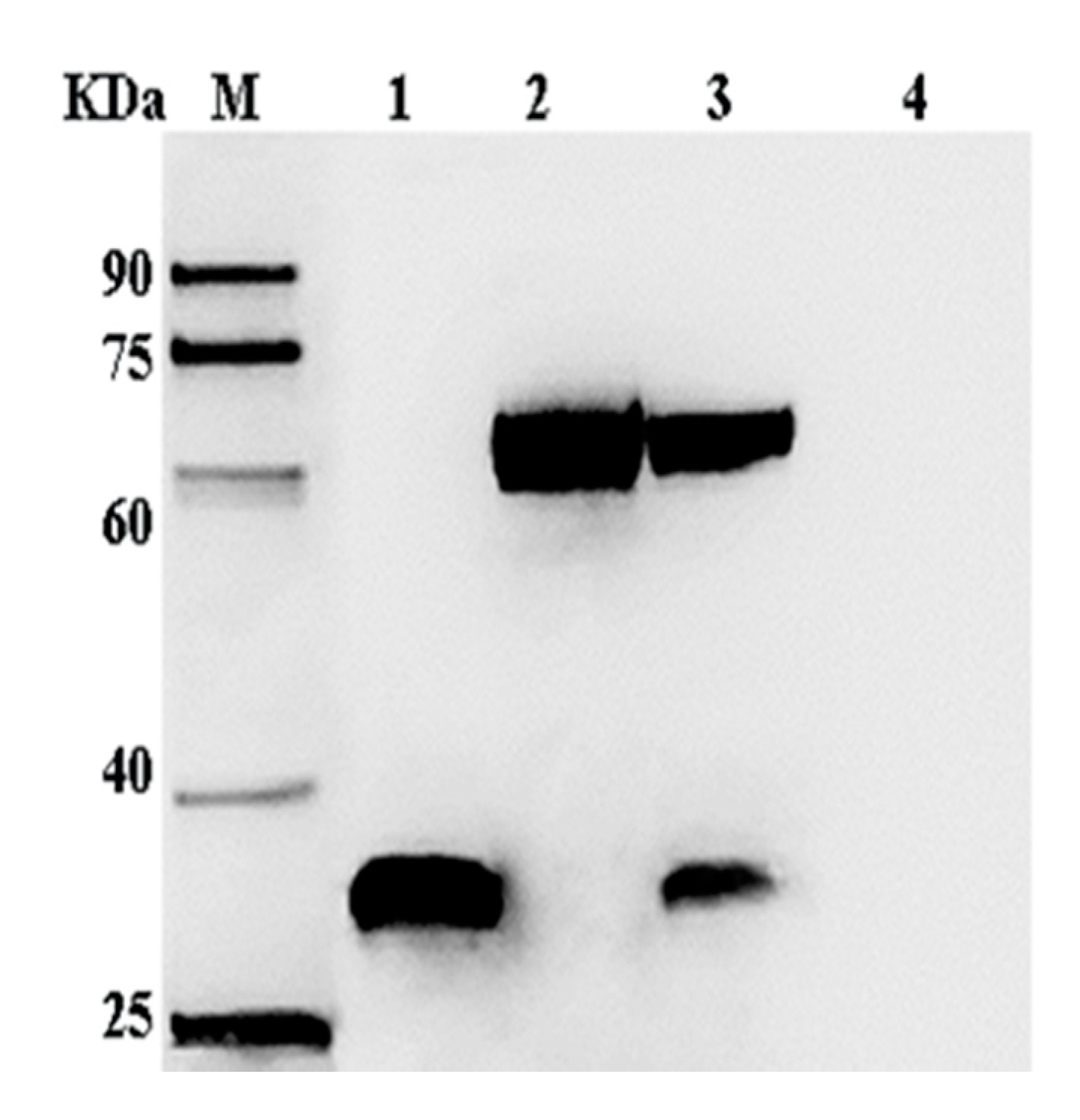
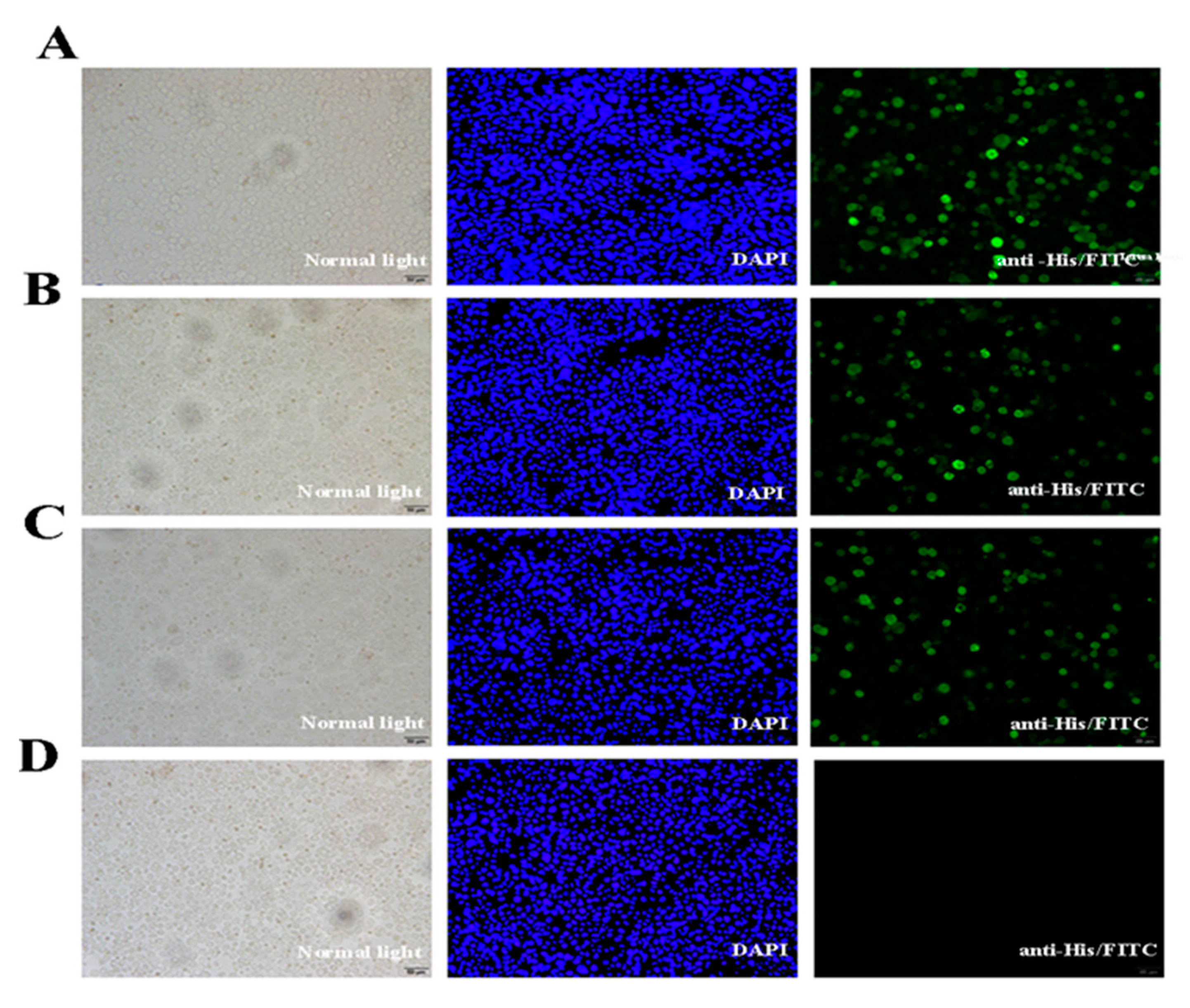
2.3. Expression of VP35 and VP4 Proteins in Sf-9 Cells
2.4. mRNA Expression of Immune-Related Genes in Different Tissues
2.5. Challenge Test
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Materials
4.3. Gene Cloning and Analysis
4.4. Construction of Donor Vectors pFastBac-VP35, pFastBac-VP4, and pFastBac-VP35-VP4
4.5. Generation of Recombinant Baculoviruses
4.6. Analysis of VP4 and VP35 Expression in Sf-9 Cells with Western Blotting
4.7. Detection of VP4 and VP35 Expression in Sf-9 Cells with Immunofluorescence Analysis
4.8. Immune Protection and Sample Collection
4.9. Detection of mRNA Expression of Immune-Related Genes by Using qRT-PCR
4.10. GCRV Challenge Test
4.11. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Huang, R.; Sun, J.; Luo, Q.; He, L.; Liao, L.; Li, Y.; Guo, F.; Zhu, Z.; Wang, Y. Genetic variations of body weight and GCRV resistance in a random mating population of grass carp. Oncotarget 2015, 6, 35433–35442. [Google Scholar] [CrossRef]
- Rao, Y.; Su, J. Insights into the antiviral immunity against grass carp (Ctenopharyngodon idella) reovirus (GCRV) in grass carp. Allergy Asthma Immunol. Res. 2015, 2015, 670437. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, Y. Morphological and physico-chemical characterization of the hemorrhagic virus of grass carp. Sci. Bull. 1984, 6, 832–835. [Google Scholar]
- Yan, Y.; Guo, H.; Sun, X.; Shao, L.; Fang, Q. Characterization of grass carp reovirus minor core protein VP4. Virol. J. 2012, 9, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Fang, Q.; Jaafar, F.M.; Cantaloube, J.-F.; Biagini, P.; De Micco, P.; De Lamballerie, X. Common evolutionary origin of aquareoviruses and orthoreoviruses revealed by genome characterization of Golden shiner reovirus, Grass carp reovirus, Striped bass reovirus and golden ide reovirus (genus Aquareovirus, family Reoviridae). Nat. Rev. Microbiol. 2002, 83, 1941–1951. [Google Scholar]
- Liu, H.; Gong, H.; Chou, H. Cloning of the Major Capsid Protein (MCP) of Grouper Iridovirus of Taiwan (TGIV) and Preliminary Evaluation of a Recombinant MCP Vaccine against TGIV. Int. J. Mol. Sci. 2015, 16, 28647–28656. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Yao, Y.; Cui, Z.; Zhang, X.; Peng, K.; Guo, X.; Wang, B.; Zhou, Y.; Li, S.; Wu, W.; et al. Comparative study of the immunoprotective effect of two DNA vaccines against grass carp reovirus. Fish Shellfish Immunol. 2018, 75, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Chi, X.; Zhang, L.; Qu, H.; Liu, Y.; Wang, X.; Zhou, J. Biotechnology, enhanced expression of GCRV VP6 in CIK cells by relative sequence optimization. Appl. Biochem. Biotechnol. 2014, 173, 2129–2139. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pei, C.; Gao, X.C.; Chen, Z.Y.; Zhang, Q.Y. Fish reovirus GCReV-109 VP33 protein elicits protective immunity in rare minnows. Arch. Virol. 2016, 161, 573–582. [Google Scholar] [CrossRef]
- Liang, H.; Fu, X.; Liu, L.; Lin, Q.; Guo, H.; Li, Y.; Liu, J.; Huang, Z.; Li, N. Inhibition of Grass Carp Reovirus Infection by DNA Aptamers against S10 Protein. J. Aquat. Anim. Health 2017, 29, 89–94. [Google Scholar] [CrossRef]
- Yan, X.Y.; Wang, Y.; Xiong, L.F.; Jian, J.C.; Wu, Z.H. Phylogenetic analysis of newly isolated grass carp reovirus. Springerplus 2014, 3, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Hang, S.; Ling, Y.; Guo, S. A plasmid containing CpG ODN as vaccine adjuvant against grass carp reovirus in grass carp Ctenopharyngodon idella. Dev. Comp. Immunol. 2016, 60, 218–227. [Google Scholar]
- Tian, Y.; Ye, X.; Zhang, L.; Deng, G.; Bai, Y. Development of a novel candidate subunit vaccine against Grass carp reovirus Guangdong strain (GCRV-GD108). Fish Shellfish Immunol. 2013, 35, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Pei, C.; Sun, X.; Zhang, C.; Li, L.; Kong, X. Novel subunit vaccine based on grass carp reovirus VP35 protein provides protective immunity against grass carp hemorrhagic disease. Fish Shellfish Immunol. 2018, 75, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Bian, Q.; Zeng, W.; Ren, P.; Sun, H.; Lin, Z.; Tang, Z.; Zhou, X.; Wang, Q.; Wang, Y.; et al. Oral delivery of Bacillus subtilis spores expressing grass carp reovirus VP4 protein produces protection against grass carp reovirus infection. Fish Shellfish Immunol. 2019, 84, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Wang, B.; Wang, N.; Yang, G.; Cao, H. Transcriptome analysis of rare minnow (Gobiocypris rarus) infected by the grass carp reovirus. Fish Shellfish Immunol. 2019, 89, 337–344. [Google Scholar] [CrossRef]
- Hui, W.; Xi, C.; Lin, L.; Qin, L.; Wen, G.; Lan, Y. Observations on the ultra-thin sections of the main organs and tissues of hemorrhagic gobiocypris rarus artificially infected bu grass carp hemorrhagic virus (GCHV). Acta Hydroch. Hydrob. 1993, 4, 342–346. [Google Scholar]
- Al-Sulaiman, A.M.; Vallely, P.J.; Klapper, P.E.; Al Baradie, R.; Almatrrouk, S.A.; Alharbi, K.K. Expression of variable viruses as herpes simplex glycoprotein D and varicella zoster gE glycoprotein using a novel plasmid based expression system in insect cell. Saudi J. Biol. Sci. 2017, 24, 1497–1504. [Google Scholar] [CrossRef]
- Chen, C.Y.; Lin, C.Y.; Chen, G.Y. Baculovirus as a gene delivery vector: Recent understandings of molecular alterations in transduced cells and latest applications. Biotechnol. Adv. 2011, 29, 618–631. [Google Scholar] [CrossRef]
- Yang, D.K.; Kim, H.H.; Nah, J.J.; Song, J.Y. Rabbit hemorrhagic disease virus variant recombinant VP60 protein induces protective immunogenicity. J. Microbiol. Biotech. 2015, 25, 1960–1965. [Google Scholar] [CrossRef]
- Li, Z.; Yin, X.; Yi, Y.; Li, X.; Li, B.; Lan, X.; Zhang, Z.; Liu, J. FMD subunit vaccine produced using a silkworm–baculovirus expression system: Protective efficacy against two type Asia1 isolates in cattle. Vet. Microbiol. 2011, 149, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Musthaq, S.S.; Madhan, S.; Hameed, A.S.S.; Kwang, J. Localization of VP28 on the baculovirus envelope and its immunogenicity against white spot syndrome virus in Penaeus monodon. Virology 2009, 391, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Yuan, R.; Zhang, M.; Zhang, T.; Gu, Y.; Zhou, Y.; Dai, Y.; Fang, P.; Feng, Y.; Hu, X.; et al. Recombinant baculovirus BacCarassius-D4ORFs has potential as a live vector vaccine against CyHV-2. Fish Shellfish Immunol. 2019, 92, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Xue, R.; Liu, L.; Cao, G.; Xu, S.; Li, J.; Zou, Y.; Chen, H.; Gong, C. Oral vaccination of BacFish-vp6 against grass carp reovirus evoking antibody response in grass carp. Fish Shellfish Immunol. 2013, 34, 348–355. [Google Scholar] [CrossRef]
- Liu, B.; Gong, Y.; Li, Z.; Hu, X.; Cao, G.; Xue, R.; Gong, C. Baculovirus-mediated GCRV vp7 and vp6 genes expression in silkworm and grass carp. Mol. Biol. Rep. 2016, 43, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Fang, J.; Pan, Q.; Wang, Y.; Xue, T.; Li, L.; Chen, T. Efficient and stable delivery of multiple genes to fish cells by a modified recombinant baculovirus system. Int. J. Mol. Sci. 2018, 19, 3767. [Google Scholar] [CrossRef]
- Ana, I.; Saint-Jean, S.R.; Pérez-Prieto, S.I.J.F. Immunology, Immunogenic and protective effects of an oral DNA vaccine against infectious pancreatic necrosis virus in fish. Fish Shellfish Immunol. 2010, 28, 562–570. [Google Scholar]
- Pei, C.; Ke, F.; Chen, Z.Y.; Zhang, Q.Y. Complete genome sequence and comparative analysis of grass carp reovirus strain 109 (GCReV-109) with other grass carp reovirus strains reveals no significant correlation with regional distribution. Arch. Virol. 2014, 159, 2435–2440. [Google Scholar] [CrossRef]
- Gao, Y.; Pei, C.; Sun, X.; Zhang, C.; Li, L.; Kong, X. Plasmid pcDNA3.1-S11 constructed based on the S11 segment of grass carp reovirus as DNA vaccine provides immune protection. NPJ Vaccines 2018, 36, 3613–3621. [Google Scholar] [CrossRef]
- Doronina, V.A.; Wu, C.; Felipe, P.D.; Sachs, M.S.; Ryan, M.D.; Brown, J.D. Site-specific release of nascent chains from ribosomes at a sense codon. Mol. Cell. Biol. 2008, 28, 4227–4239. [Google Scholar] [CrossRef]
- Szymczak-Workman, A.L.; Vignali, K.M.; Vignali, D.A.A. Design and Construction of 2A Peptide-Linked Multicistronic Vectors. Cold Spring Harb. Protoc. 2012, 2012, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Mir, F.A.; Kaufmann, S.H.E.; Eddine, A.N. A multicistronic DNA vaccine induces significant protection against tuberculosis in mice and offers flexibility in the expressed antigen repertoire. Clin. Vaccine Immunol. 2009, 16, 1467–1475. [Google Scholar] [CrossRef] [PubMed]
- Langevin, C.; Aleksejeva, E.; Passoni, G.; Palha, N.; Levraud, J.P.; Boudinot, P. The Antiviral Innate Immune Response in Fish: Evolution and Conservation of the IFN System. J. Mol. Biol. 2013, 425, 4904–4920. [Google Scholar] [CrossRef] [PubMed]
- Hao, K.; Chen, X.H.; Qi, X.Z.; Zhu, B.; Wang, G.X.; Ling, F. Display of GCRV vp7 protein on the surface of Escherichia coli and its immunoprotective effects in grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2018, 72, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Gong, Y.X.; Liu, G.L.; Zhu, B.; Wang, G.X. Protective immunity of grass carp immunized with DNA vaccine against Aeromonas hydrophila by using carbon nanotubes as a carrier molecule. Fish shellfish Immunol. 2016, 55, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Uribe, C.; Folch, H.; Enríquez, R.; Moran, G.J.V.M. Innate and adaptive immunity in teleost fish: A review. Vet. Med. (Praha) 2011, 56, 486–503. [Google Scholar] [CrossRef]
- Baud, V.; Karin, M. Signal transduction by tumor necrosis factor and its relatives. Trends Cell Biol. 2011, 11, 372–377. [Google Scholar] [CrossRef]
- Dong, X.; Zhang, Z.; Meng, S.; Pan, C.; Yang, M.; Wu, X.; Yang, L.; Xu, H. Parental exposure to bisphenol A and its analogs influences zebrafish offspring immunity. Total Environ. 2018, 610, 291–297. [Google Scholar] [CrossRef]
- Zhang, J.; Kong, X.; Zhou, C.; Li, L.; Nie, G.; Li, X. Toll-like receptor recognition of bacteria in fish: Ligand specificity and signal pathways. Fish Shellfish Immunol. 2017, 41, 380–388. [Google Scholar] [CrossRef]
- Nishiya, T.; Kajita, E.; Miwa, S.; Defranco, A.L. TLR3 and TLR7 are targeted to the same intracellular compartments by distinct regulatory elements. J. Biol. Chem. 2005, 44, 37107–37117. [Google Scholar] [CrossRef]
- Schroeder, M.; Bowie, A.G. TLR3 in antiviral immunity: Key player or bystander. Trends Immunol. 2005, 26, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, A.; Oshiumi, H.; Tsujita, T.; Mitani, H.; Kasai, H.; Yoshimizu, M.; Matsumoto, M.; Seya, T. Teleost TLR22 recognizes RNA duplex to induce IFN and protect cells from birnaviruses. J. Immunol. 2008, 181, 3474–3485. [Google Scholar] [CrossRef] [PubMed]
- Lelania, B.A.; Geoffrey, C.W. Activation of TLR3 and TLR5 in channel catfish exposed to virulent Edwardsiella ictaluri. Dev. Comp. Immunol. 2005, 29, 713–721. [Google Scholar]
- Su, J.; Zhu, Z.; Wang, Y.; Zou, J.; Hu, W. Toll-like receptor 3 regulates Mx expression in rare minnow Gobiocypris rarus after viral infection. Immunogenetics 2008, 60, 195–205. [Google Scholar] [CrossRef]
- Veronica, B.; Borre, R. Characterization of Atlantic halibut (Hippoglossus hippoglossus) Mx protein expression. Dev. Comp. Immunol. 2004, 28, 1037–1047. [Google Scholar]
- Haller, O.; Kochs, G. Interferon-induced Mx proteins: Dynamin-like GTPases with antiviral activity. Traffic 2003, 3, 710–717. [Google Scholar] [CrossRef]
- Larsen, R.; Rkenes, T.P.; Robertsen, B. Inhibition of infectious pancreatic necrosis virus replication by atlantic salmon Mx1 protein. J. Virol. 2004, 78, 7938–7944. [Google Scholar] [CrossRef]
- DeWitte-Orr, S.J.; Leong, J.A.C.; Bols, N.C. Induction of antiviral genes, Mx and vig-1, by dsRNA and Chum salmon reovirus in rainbow trout monocyte/macrophage and fibroblast cell lines. Fish Shellfish Immunol. 2007, 22, 670–682. [Google Scholar] [CrossRef]
- Tang, D.; Gao, Y.; Wang, R. Characterization, genomic organization, and expression profiles of MyD88, a key adaptor molecule in the TLR signaling pathways in miiuy croaker (Miichthys miiuy). Fish Physiol. Biochem. 2012, 38, 1667–1677. [Google Scholar] [CrossRef]
- Salinas, I.; Zhang, Y.A.; Sunyer, O.J. Mucosal immunoglobulins and B cells of teleost fish. Dev. Comp. Immunol. 2011, 35, 1346–1365. [Google Scholar] [CrossRef]
- Dai, Z.; Li, J.; Hu, C.; Wang, F.; Wang, B.; Shi, X.; Hou, Q.; Huang, W.; Lin, G. Transcriptome data analysis of grass carp (Ctenopharyngodon idella) infected by reovirus provides insights into two immune-related genes. Fish Shellfish Immunol. 2017, 64, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Ou, M.; Huang, R.; Luo, Q.; Xiong, L.; Chen, K.; Wang, Y. Characterisation of scavenger receptor class B type 1 in rare minnow (Gobiocypris rarus). Fish Shellfish Immunol. 2019, 89, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, C.; Ma, D.; Liu, M.; Huang, S. Lipid accumulation, oxidative stress and immune-related molecules affected by tributyltin exposure in muscle tissues of rare minnow ( Gobiocypris rarus ). Fish Shellfish Immunol. 2017, 71, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Hong, X.; Chen, R.; Yuan, L.; Zha, J.; Qin, J. New cytokines and TLR pathway signaling molecules in Chinese rare minnow (Gobiocypris rarus): Molecular characterization, basal expression, and their response to chlorpyrifos. Chemosphere 2018, 199, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, X.; Yan, S.; Zha, J.; Ma, X. The immune responses of the Chinese rare minnow (Gobiocypris rarus) exposed to environmentally relevant concentrations of cypermethrin and subsequently infected by the bacteria Pseudomonas fluorescens. Int. J. Environ. Pollut. 2019, 250, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Zhao, X.; Tian, X.; Li, J.; Zha, J. Changes of hematological and biochemical parameters revealed genotoxicity and immunotoxicity of neonicotinoids on Chinese rare minnows (Gobiocypris rarus). Int. J. Environ. Pollut. 2018, 233, 862–871. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, C.; Vakharia, V.N.; Zhou, Y.; Jiang, N.; Liu, W.; Meng, Y.; Li, Y.; Xue, M.; Zhang, J.; Zeng, L.; et al. A Novel Subunit Vaccine Based on Outer Capsid Proteins of Grass Carp Reovirus (GCRV) Provides Protective Immunity against GCRV Infection in Rare Minnow (Gobiocypris rarus). Pathogens 2020, 9, 945. https://doi.org/10.3390/pathogens9110945
Mu C, Vakharia VN, Zhou Y, Jiang N, Liu W, Meng Y, Li Y, Xue M, Zhang J, Zeng L, et al. A Novel Subunit Vaccine Based on Outer Capsid Proteins of Grass Carp Reovirus (GCRV) Provides Protective Immunity against GCRV Infection in Rare Minnow (Gobiocypris rarus). Pathogens. 2020; 9(11):945. https://doi.org/10.3390/pathogens9110945
Chicago/Turabian StyleMu, Changyong, Vikram N. Vakharia, Yong Zhou, Nan Jiang, Wenzhi Liu, Yan Meng, Yiqun Li, Mingyang Xue, Jieming Zhang, Lingbing Zeng, and et al. 2020. "A Novel Subunit Vaccine Based on Outer Capsid Proteins of Grass Carp Reovirus (GCRV) Provides Protective Immunity against GCRV Infection in Rare Minnow (Gobiocypris rarus)" Pathogens 9, no. 11: 945. https://doi.org/10.3390/pathogens9110945
APA StyleMu, C., Vakharia, V. N., Zhou, Y., Jiang, N., Liu, W., Meng, Y., Li, Y., Xue, M., Zhang, J., Zeng, L., Zhong, Q., & Fan, Y. (2020). A Novel Subunit Vaccine Based on Outer Capsid Proteins of Grass Carp Reovirus (GCRV) Provides Protective Immunity against GCRV Infection in Rare Minnow (Gobiocypris rarus). Pathogens, 9(11), 945. https://doi.org/10.3390/pathogens9110945