
Detection of Lotmaria passim and Crithidia mellificae in Selected Bumblebee Species


Abstract
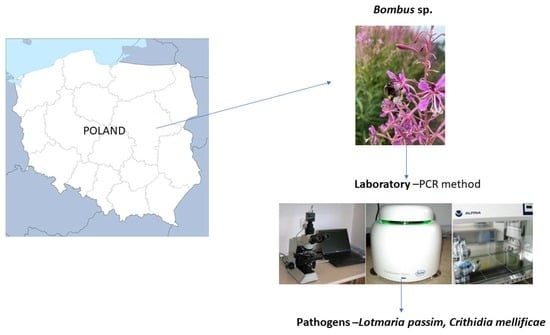
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Winter, K.; Adams, L.; Thorp, R.; Inouye, D.; Day, L.; Ascher Buchmann, S. Importation of Non-Native Bumble Bees into North America: Potential Consequences of Using Bombus Terrestris and Other Non-Native Bumble Bees for the Greenhouse Crop Pollination in Canada, Mexico, and the United States; North American Pollinator Protection Campaign: San Francisco, CA, USA, 2006. [Google Scholar]
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–882. [Google Scholar] [CrossRef]
- Mobley, M.W.; Gegear, R.J. Immune-cognitive system connectivity reduces bumblebee foraging success in complex multisensory floral environments. Sci. Rep. 2018, 8, 5953. [Google Scholar] [CrossRef] [PubMed]
- Pawlikowski, T.; Pawlikowski, K. Trzmielowate Polski (Hymenoptera: Apidae: Bombini); Wydawnictwo Naukowe Uniwersytetu Mikołaja Kopernika: Toruń, Poland, 2012. [Google Scholar]
- Mobley, M.W.; Gegear, R.J. One size does not fit all: Caste and sex differences in the response of bumblebees (Bombus impatiens) to chronic oral neonicotinoid exposure. PLoS ONE 2018, 13, e0200041. [Google Scholar] [CrossRef] [PubMed]
- Colla, S.R.; Otterstatter, M.C.; Gegear, R.J.; Thomson, J.D. Plight of the bumble bee: Pathogen spillover from commercial to wild populations. Biol. Conserv. 2006, 129, 461–467. [Google Scholar] [CrossRef]
- Goulson, D.; Hughes, W.O.H. Mitigating the anthropogenic spread of bee parasites to protect wild pollinators. Biol. Conserv. 2015, 191, 10–19. [Google Scholar] [CrossRef]
- Sokol, R.; Michalczyk, M.; Micholap, P. Preliminary studies on the occurrence of honeybee pathogens in the national bumblebee population. Ann. Parasitol. 2018, 64, 385–390. [Google Scholar] [PubMed]
- Kostygov, A.Y.; Karnkowska, A.; Votýpka, J.; Tashyreva, D.; Maciszewski, K.; Yurchenko, Y.; Lukeš, J. Euglenozoa: Taxonomy, diversity and ecology, symbioses and viruses. Open Biol. 2021, 11, 200407. [Google Scholar] [CrossRef] [PubMed]
- Maslov, D.A.; Votýpka, J.; Yurchenko, V.; Lukeš, J. Diversity and phylogeny of insect trypanosomatids: All that is hidden shall be revealed. Trends Parasitol. 2013, 29, 43–52. [Google Scholar] [CrossRef]
- Fantham, H.B.; Porter, A. Note on certain protozoa found in bees. Suppl. J. Board. Agricul. 1912, 19, 138. [Google Scholar]
- Giavarini, I. Sui flagellati dell’ intestino tenue dell’ape domestica. Bolletino Zool. 1950, 17, 603–608. [Google Scholar] [CrossRef]
- Arismendi, N.; Caro, S.; Castro, M.P.; Vargas, M.; Riveros, G.; Venegas, T. Impact of Mixed Infections of Gut Parasites Lotmaria passim and Nosema ceranae on the Lifespan and Immune-related Biomarkers in Apis mellifera. Insects 2020, 11, 420. [Google Scholar] [CrossRef] [PubMed]
- Buendía-Abad, M.; García-Palencia, P.; de Pablos, L.M.; Alunda, J.M.; Osuna, A.; Martín-Hernández, R.; Higes, M. First description of Lotmaria passim and Crithidia mellificae haptomonad stages in the honeybee hindgut. Int. J. Parasitol. 2022, 52, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Sacks, D.; Kamhawi, S. Molecular Aspects of Parasite-Vector and Vector-Host Interactions in Leishmaniasis. Annu. Rev. Microbiol. 2001, 55, 453–483. [Google Scholar] [CrossRef]
- Schmid-Hempel, R.; Tognazzo, M. Molecular divergence defines two distinct lineages of Crithidia bombi (Trypanosomatidae), parasites of bumblebees. J. Eukaryot. Microbiol. 2010, 57, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Graystock, P.; Goulson, D.; Hughes, W.O.H. The relationship between managed bees and the prevalence of parasites in bumblebees. Peerj 2014, 2, e522. [Google Scholar] [CrossRef]
- Strobl, V.; Yañez, O.; Straub, L.; Albrecht, M.; Neumann, P. Trypanosomatid parasites infecting managed honeybees and wild solitary bees. Int. J. Parasitol. 2019, 49, 605–613. [Google Scholar] [CrossRef]
- Schwarz, R.S.; Bauchan, G.R.; Murphy, C.A.; Ravoet, J.; De Graaf, D.C.; Evans, J.D. Characterization of two species of trypanosomatidae from the Honey Bee Apis mellifera: Crithidia mellificae Langridge and McGhee, and Lotmaria passim n. gen., n. sp. J. Eukaryot. Microbiol. 2015, 62, 567–583. [Google Scholar] [CrossRef]
- Runckel, C.; Flenniken, M.L.; Engel, J.C.; Ruby, J.G.; Ganem, D.; Andino, R.; DeRisi, J.L. Temporal analysis of the honey bee microbiome reveals four novel viruses and seasonal prevalence of known viruses, Nosema, and Crithidia. PLoS ONE 2011, 6, e20656. [Google Scholar] [CrossRef]
- Schwarz, R.S.; Evans, J.D. Single and mixed-species trypanosome andmicrosporidia infections elicit distinct, ephemeral cellular and humoral immune responses in honey bees. Dev. Comp. Immunol. 2013, 40, 300–310. [Google Scholar] [CrossRef]
- Lukeš, J.; Wheeler, R.; Jirsová, D.; David, V.; Archibald, J.M. Massive mitochondrial DNA content in diplonemid and kinetoplastid protists. IUBMB Life 2018, 70, 1267–1274. [Google Scholar] [CrossRef]
- Arismendi, N.; Bruna, A.; Zapata, N.; Vargas, M. PCR-specific detection of recently described Lotmaria passim (Trypanosomatidae) in Chilean apiaries. J. Invertebr. Pathol. 2016, 134, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Castelli, L.; Branchiccela, B.; Invernizzi, C.; Tomasco, I.; Basualdo, M.; Rodriguez, M.; Zunino, P.; Antúnez, K. Detection of Lotmaria passim in Africanized and European honey bees from Uruguay, Argentina and Chile. J. Invertebr. Pathol. 2019, 160, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Vejnovic, B.; Stevanovic, J.; Schwarz, R.S.; Aleksic, N.; Mirilovic, M.; Jovanovic, N.M.; Stanimirovic, Z. Quantitative PCR assessment of Lotmaria passim in Apis mellifera colonies co-infected naturally with Nosema ceranae. J. Invertebr. Pathol. 2018, 151, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Michalczyk, M.; Sokół, R.; Bancerz-Kisiel, A. Presence of Lotmaria passim, Crithidia mellificae and Nosema spp. in differently aged Apis mellifera brood. J. Apic. Res. 2022. [Google Scholar] [CrossRef]
- Ravoet, J.; Maharramov, J.; Meeus, I.; de Smet, L.; Wenseleers, T.; Smagghe, G.; de Graaf, D.C. Comprehensive bee pathogen screening in Belgium reveals Crithidia mellificae as a new contributory factor to winter mortality. PLoS ONE 2013, 8, e72443. [Google Scholar] [CrossRef] [PubMed]
- Cepero, A.; Ravoet, J.; Gómez-Moracho, T.; Bernal, J.; Del Nozal, M.J.; Bartolomé, C.; Maside, X.; Meana, A.; González-Porto, A.V.; de Graaf, D.C.; et al. Holistic screening of collapsing honey bee colonies in Spain: A case study, BMC Res. Notes 2014, 7, 649. [Google Scholar] [CrossRef]
- Bartolomé, C.; Buendía-Abad, M.; Benito, M.; Sobrino, B.; Amigo, J.; Carracedo, A.; Martín-Hernández, R.; Higes, M.; Maside, X. Longitudinal analysis on parasite diversity in honeybee colonies: New taxa, high frequency of mixed infections and seasonal patterns of variation. Sci. Rep. 2020, 10, 10454. [Google Scholar] [CrossRef]
- D’Alvise, P.; Seeburger, V.; Gihring, K.; Kieboom, M.; Hasselmann, M. Seasonaldynamics and co-occurence patterns of honey bee pathogens revealed by high-throughput RT- qPCR. Ecol. Evol. 2019, 9, 10241–10252. [Google Scholar] [CrossRef]
- Nanetti, A.; Ellis, J.D.; Cardaio, I.; Cilia, G. Detection of Lotmaria passim, Crithidia mellificae and Replicative Forms of Deformed Wing Virus and Kashmir Bee Virus in the Small Hive Beetle (Aethina tumida). Pathogens 2021, 10, 372. [Google Scholar] [CrossRef]
- Michalczyk, M.; Bancerz-Kisiel, A.; Sokół, R. Lotmaria passim as Third Parasite Gastrointestinal Tract of Honey Bees Living in Tree Trunk. J. Apic. Sci. 2020, 64, 143–151. [Google Scholar] [CrossRef]
- Arismendi, N.; Riveros, G.; Zapata, N.; Smagghe, G.; González, C.; Vargas, M. Occurrence of bee viruses and pathogens associated with emerging infectious diseases in native and non-native bumble bees in southern Chile. Biol. Invasions 2021, 23, 1175–1189. [Google Scholar] [CrossRef]
- Gamboa, V.; Ravoet, J.; Brunain, M.; Smagghe, G.; Meeus, I.; Figueroa, J.; de Graaf, D. Bee pathogens found in Bombus atratus from Colombia: A case study. J. Invertebr. Pathol. 2015, 129, 36–39. [Google Scholar] [CrossRef]
- Plischuk, S.; de Landa, G.F.; Revainera, P.; Quintana, S.; Pocco, M.E.; Cigliano, M.M.; Lange, C.E. Parasites and pathogens associated with native bumble bees (Hymenoptera: Apidae: Bombus spp.) from highlands in Bolivia and Peru. Stud. Neotrop. Fauna Environ. 2021, 56, 93–98. [Google Scholar] [CrossRef]
- Pislak Ocepek, M.; Toplak, I.; Zajc, U.; Bevk, D. The Pathogens Spillover and Incidence Correlation in Bumblebees and Honeybees in Slovenia. Pathogens 2021, 10, 884. [Google Scholar] [CrossRef] [PubMed]
- Bartolomé, C.; Buendía-Abad, M.; Ornosa, C.; De la Rúa, P.; Martín-Hernández, R.; Higes, M.; Maside, X. Bee Trypanosomatids: First Steps in the Analysis of the Genetic Variation and Population Structure of Lotmaria passim, Crithidia bombi and Crithidia mellificae. Microb. Ecol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Averill, A.L.; Couto, A.V.; Andersen, J.C.; Elkinton, J.S. Parasite Prevalence May Drive the Biotic Impoverishment of New England (USA) Bumble Bee Communities. Insects 2021, 12, 941. [Google Scholar] [CrossRef] [PubMed]
- Schlüns, H.; Sadd, B.M.; Schmid-Hempel, P.; Crozier, R.H. Infection with the trypanosome Crithidia bombi and expression of immune-related genes in the bumblebee Bombus terrestris. Dev. Comp. Immunol. 2010, 34, 705–709. [Google Scholar] [CrossRef]
- Tlak Gajger, I.; Šimenc, L.; Toplak, I. The First Detection and Genetic Characterization of Four Different Honeybee Viruses in Wild Bumblebees from Croatia. Pathogens 2021, 10, 808. [Google Scholar] [CrossRef]
- Toplak, I.; Šimenc, L.; Pislak Ocepek, M.; Bevk, D. Determination of Genetically Identical Strains of Four Honeybee Viruses in Bumblebee Positive Samples. Viruses 2020, 12, 1131. [Google Scholar] [CrossRef]
- Alger, S.A.; Burnham, P.A.; Boncristiani, H.F.; Brody, A.K. RNA virus spillover from managed honeybees (Apis mellifera) to wild bumblebees (Bombus spp.). PLoS ONE 2019, 14, e0217822. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Number of Samples | Bumblebee Species | L. passim | C. mellificae |
|---|---|---|---|
| 1. | B. lapidarius | - | - |
| 2. | B. lapidarius | - | - |
| 3. | B. lapidarius | - | - |
| 4. | B. lapidarius | - | - |
| 5. | B. lucorum | - | - |
| 6. | B. pascuorum | - | - |
| 7. | B. pascuorum | + | - |
| 8. | B. pascuorum | - | - |
| 9. | B. pascuorum | - | - |
| 10. | B. pascuorum | - | - |
| 11. | B. pascuorum | - | - |
| 12. | B. terrestris | - | - |
| 13. | B. terrestris | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michalczyk, M.; Sokół, R. Detection of Lotmaria passim and Crithidia mellificae in Selected Bumblebee Species. Pathogens 2022, 11, 1053. https://doi.org/10.3390/pathogens11091053
Michalczyk M, Sokół R. Detection of Lotmaria passim and Crithidia mellificae in Selected Bumblebee Species. Pathogens. 2022; 11(9):1053. https://doi.org/10.3390/pathogens11091053
Chicago/Turabian StyleMichalczyk, Maria, and Rajmund Sokół. 2022. "Detection of Lotmaria passim and Crithidia mellificae in Selected Bumblebee Species" Pathogens 11, no. 9: 1053. https://doi.org/10.3390/pathogens11091053
APA StyleMichalczyk, M., & Sokół, R. (2022). Detection of Lotmaria passim and Crithidia mellificae in Selected Bumblebee Species. Pathogens, 11(9), 1053. https://doi.org/10.3390/pathogens11091053