
Prevalence of Virus Infections and GLRaV-3 Genetic Diversity in Selected Clones of Croatian Indigenous Grapevine Cultivar Plavac Mali

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
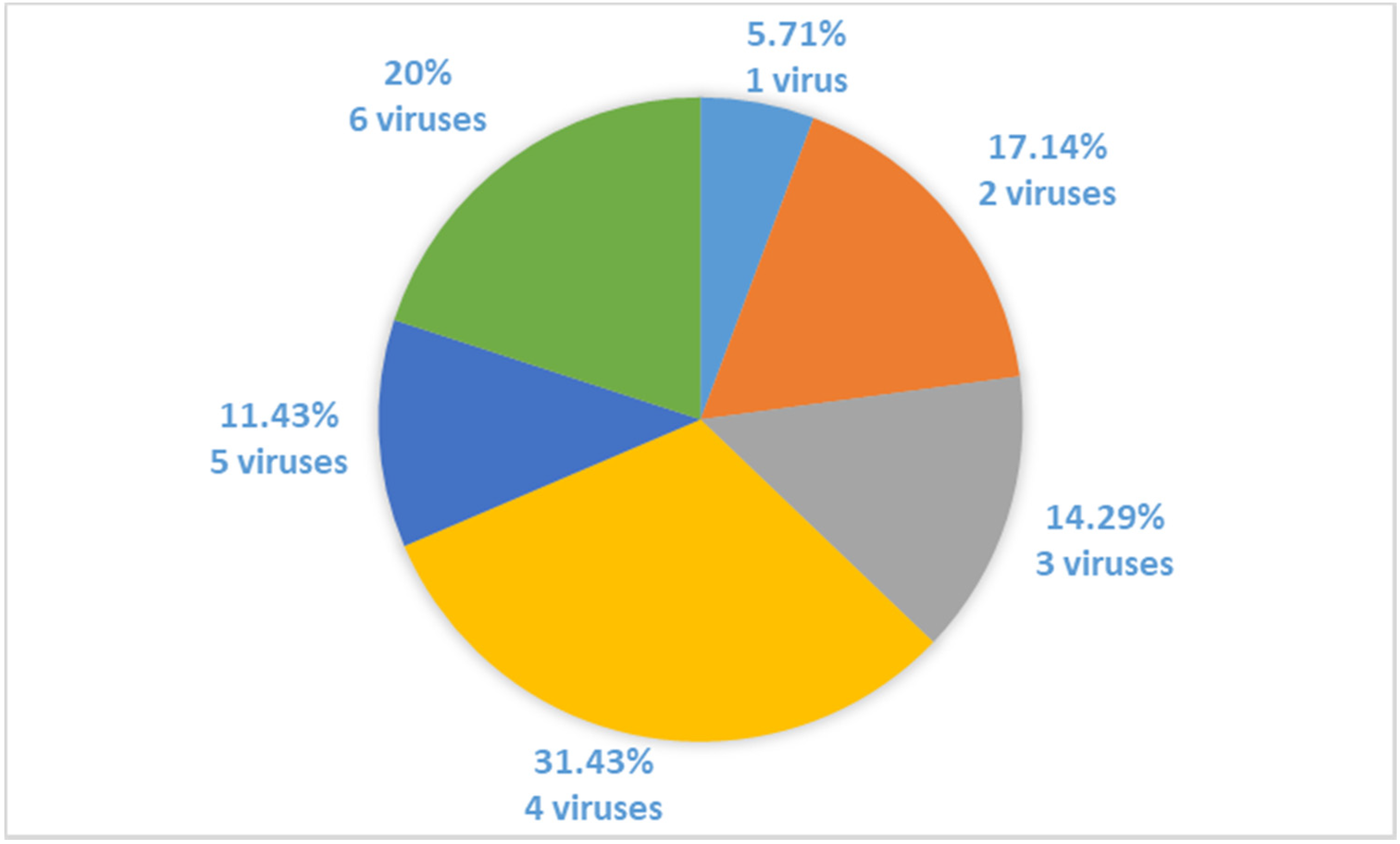
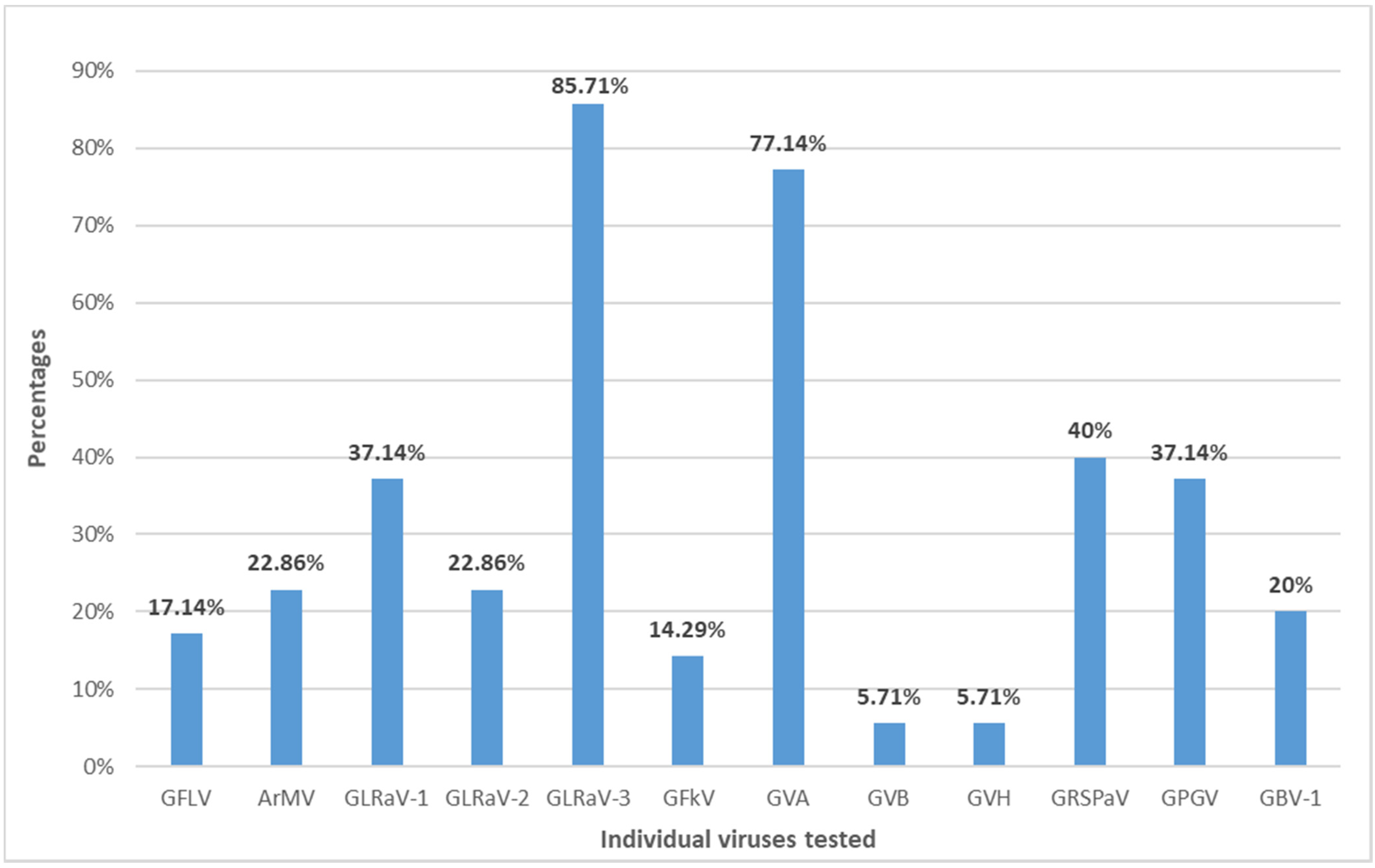
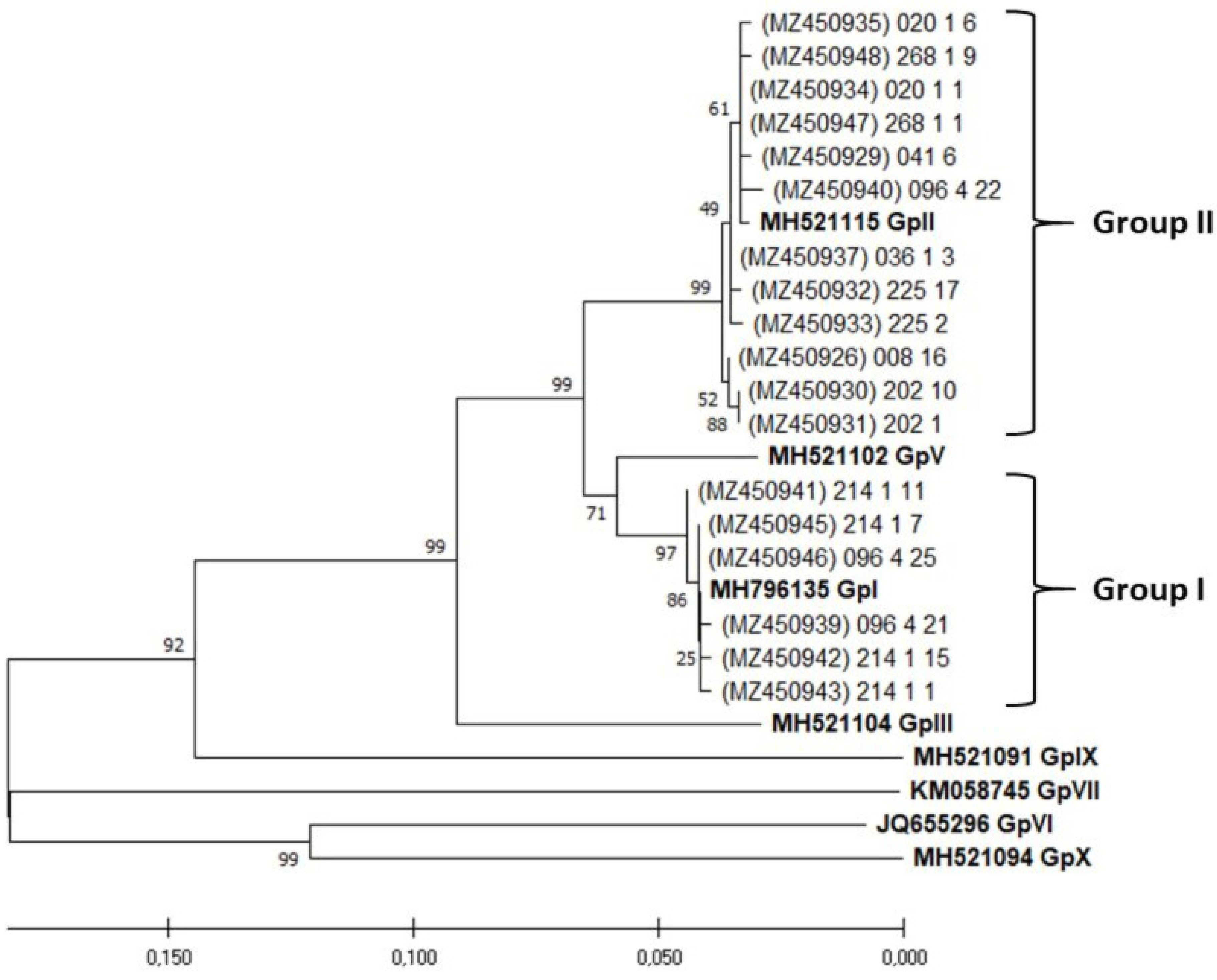
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material and Symptoms Assessment
4.2. RNA Extraction, cDNA Synthesis and DNA Extraction
4.3. Virus Detection
4.4. Cloning and Single-Stranded Conformational Polymorphism (SSCP) Analysis
4.5. Sequencing and Sequence Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Croatian Bureau of Statistics Basic Survey on Vineyard Structure. 2015. Available online: https://www.dzs.hr/Hrv_Eng/publication/2016/01-01-33_01_2016.htm (accessed on 11 November 2020).
- Maletić, E.; Pejić, I.; Kontić, J.K.; Piljac, J.; Dangl, G.S.; Vokurka, A.; Lacombe, T.; Mirošević, N.; Meredith, C.P. Zinfandel, Dobričić, and Plavac Mali: The genetic relationship among three cultivars of the Dalmatian coast of Croatia. Am. J. Enol. Vitic. 2004, 55, 174–180. [Google Scholar]
- Žulj Mihaljević, M.; Maletić, E.; Preiner, D.; Zdunić, G.; Bubola, M.; Zyprian, E.; Pejić, I. Genetic diversity, population structure, and parentage analysis of Croatian grapevine germplasm. Genes 2020, 11, 737. [Google Scholar] [CrossRef]
- Preiner, D.; Poljanić, M.; Kontić, J.K.; Marković, Z.; Maletić, E. Amphelographic characteristics of cv. Plavac mali (V. vinifera L.) clone candidates in year 2008. In Proceedings of the 45. Hrvatski i 5. Međunarodni Simpozij Agronoma, Opatija, Croatia, 15–19 February 2010; p. 1193. [Google Scholar]
- Zdunić, G.; Maletić, E.; Vokurka, A.; Karoglan Kontić, J.; Pezo, I.; Pejić, I. Phenotypical, sanitary and ampelometric variability within the population of cv. Plavac Mali (Vitis vinifera L.). Agric. Conspec. Sci. 2007, 72, 117–128. [Google Scholar]
- Pejic, I.; Maletic, E.; Karoglan Kontic, J.; Kozina, B.; Mirosevic, N. Diversity of autochthonous grapevine genotypes in Croatia. In Proceedings of the VII International Symposium on Grapevine Genetics and Breeding 528, Montpellier, France, 6–8 July 1998; pp. 67–74. [Google Scholar]
- Mucalo, A.; Maletić, E.; Zdunić, G. Extended harvest date alter flavonoid composition and chromatic characteristics of Plavac Mali (Vitis vinifera L.) grape berries. Foods 2020, 9, 1155. [Google Scholar] [CrossRef]
- Žurga, P.; Vahčić, N.; Pasković, I.; Banović, M.; Staver, M.M. Croatian Wines from Native Grape Varieties Have Higher Distinct Phenolic (Nutraceutic) Profiles than Wines from Non-Native Varieties with the Same Geographic Origin. Chem. Biodivers. 2019, 16, e1900218. [Google Scholar] [CrossRef] [PubMed]
- Budić-Leto, I.; Zdunić, G.; Gajdoš-Kljusurić, J.; Mucalo, A.; Vrhovšek, U. Differentiation between Croatian dessert wine Prošek and dry wines based on phenolic composition. J. Food Compos. Anal. 2017, 62, 211–216. [Google Scholar] [CrossRef]
- Jelaska, M. Ampelografija Dalmatinskih Sorata; Institut for Adriatic Crops and Carst Reclamation: Split, Croatia, 1960. [Google Scholar]
- Martelli, G.P. An Overview on Grapevine Viruses, Viroids and the Diseases They Cause. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 31–46. ISBN 9783319577067. [Google Scholar]
- Digiaro, M.; Elbeaino, T.; Martelli, G.P. Grapevine fanleaf virus and other Old World nepoviruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Cham, Switzerland, 2017; pp. 47–82. [Google Scholar]
- Atallah, S.S.; Gómez, M.I.; Fuchs, M.F.; Martinson, T.E. Economic impact of grapevine leafroll disease on Vitis vinifera cv. Cabernet franc in Finger Lakes vineyards of New York. Am. J. Enol. Vitic. 2012, 63, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Habili, N.; Nutter, F.W. Temporal and spatial analysis of grapevine leafroll-associated virus 3 in pinot noir grapevines in Australia. Plant Dis. 1997, 81, 625–628. [Google Scholar] [CrossRef] [Green Version]
- Naidu, R.A.; O’Neil, S.; Walsh, D. Grapevine Leafroll Disease. In WSU Extension Bulletin EB2027E; 2008; 20p, Available online: http://whatcom.wsu.edu/ag/documents/smallfruit/eb2027e.pdf (accessed on 13 May 2021).
- Maree, H.J.; Almeida, R.P.P.; Bester, R.; Chooi, K.M.; Cohen, D.; Dolja, V.V.; Fuchs, M.F.; Golino, D.A.; Jooste, A.E.C.; Martelli, G.P.; et al. Grapevine leafroll-associated virus 3. Front. Microbiol. 2013, 4, 82. [Google Scholar] [CrossRef] [Green Version]
- Burger, J.T.; Maree, H.J.; Gouveia, P.; Naidu, R.A. Grapevine leafroll-associated virus 3. In Grapevine Viruses: Molecular Biology, Diagnostics and Management, 1st ed.; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 167–194. ISBN 9783319577067. [Google Scholar]
- Sabanadzovic, S.; Aboughanem-Sabanadzovic, N.; Martelli, G.P. Grapevine fleck and similar viruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management, 1st ed.; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 331–349. [Google Scholar]
- Martelli, G.P. Virus Diseases of Grapevine. ELS 2001, 1, 1–8. [Google Scholar] [CrossRef]
- Diaz-Lara, A.; Brisbane, R.S.; Aram, K.; Golino, D.; Al Rwahnih, M. Detection of new vitiviruses infecting grapevine in California. Arch. Virol. 2019, 164, 2573–2580. [Google Scholar] [CrossRef] [PubMed]
- Rosa, C.; Jimenez, J.F.; Margaria, P.; Rowhani, A. Symptomatology and Effects of Viruses Associated with Rugose Wood Complex on the Growth of Four Different Rootstocks. Am. J. Enol. Vitic. 2011, 2, 207–213. [Google Scholar] [CrossRef]
- Saldarelli, P.; Giampetruzzi, A.; Morelli, M.; Malossini, U.; Pirolo, C.; Bianchedi, P.; Gualandri, V. Genetic variability of Grapevine Pinot gris virus and its association with Grapevine leaf mottling and deformation. Phytopathology 2015, 105, 555–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolja, V.V.; Meng, B.; Martelli, G.P. Evolutionary aspects of grapevine virology. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Cham, Switzerland, 2017; pp. 659–688. [Google Scholar]
- Vončina, D.; Almeida, R.P.P. Screening of some Croatian autochthonous grapevine varieties reveals a multitude of viruses, including novel ones. Arch. Virol. 2018, 163, 2239–2243. [Google Scholar] [CrossRef] [Green Version]
- Turturo, C.; Saldarelli, P.; Yafeng, D.; Digiaro, M.; Minafra, A.; Savino, V.; Martelli, G.P. Genetic variability and population structure of Grapevine leafroll-associated virus 3 isolates. J. Gen. Virol. 2005, 86, 217–224. [Google Scholar] [CrossRef]
- Bester, R.; Maree, H.J.; Burger, J.T. Complete nucleotide sequence of a new strain of grapevine leafroll-associated virus 3 in South Africa. Arch. Virol. 2012, 157, 1815–1819. [Google Scholar] [CrossRef]
- Chooi, K.M.; Cohen, D.; Pearson, M.N. Generic and sequence-variant specific molecular assays for the detection of the highly variable Grapevine leafroll-associated virus 3. J. Virol. Methods 2013, 189, 20–29. [Google Scholar] [CrossRef]
- Diaz-Lara, A.; Klaassen, V.; Stevens, K.; Sudarshana, M.R.; Rowhani, A.; Maree, H.J.; Chooi, K.M.; Blouin, A.G.; Habili, N.; Song, Y.; et al. Characterization of grapevine leafroll-associated virus 3 genetic variants and application towards RT-qPCR assay design. PLoS ONE 2018, 13, e0208862. [Google Scholar] [CrossRef] [Green Version]
- Naidu, R.A.; Maree, H.J.; Burger, J.T. Grapevine Leafroll Disease and Associated Viruses: A Unique Pathosystem. Annu. Rev. Phytopathol. 2015, 53, 613–634. [Google Scholar] [CrossRef]
- Karoglan Kontić, J.; Preiner, D.; Šimon, S.; Zdunić, G.; Poljuha, D.; Maletić, E. Sanitary status of Croatian native grapevine varieties. Agric. Conspec. Sci. 2009, 74, 99–103. [Google Scholar]
- Vončina, D.; Al Rwahnih, M.; Rowhani, A.; Gouran, M.; Almeida, R.P.P. Viral diversity in autochthonous croatian grapevine cultivars. Plant Dis. 2017, 101, 1230–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šarić, A.; Corte, A. Sintomatologia della degenerazione infetiva della vite in Istria. Atti Ist. Bot. Lab. Crit. Univ. Pavia 1959, 17, 3–8. [Google Scholar]
- Poljuha, D.; Sladonja, B.; Bubola, M. Incidence of viruses infecting grapevine varieties in Istria (Croatia). J. Food Agric. Environ. 2010, 8, 166–169. [Google Scholar]
- Vončina, D.; Preiner, D.; Šimon, S.; Cvjetković, B.; Maletić, E.; Pejić, I.; Kontić, J. Distribution of nine viruses in croatian autochthonous grapevine (Vitis vinifera L.) cultivars from dalmatian region included in clonal selection. J. Cent. Eur. Agric. 2019, 20, 262–273. [Google Scholar] [CrossRef]
- Hančević, K.; Saldarelli, P.; Čarija, M.; Černi, S.; Zdunić, G.; Mucalo, A.; Radić, T. Predominance and Diversity of GLRaV-3 in Native Vines of Mediterranean Croatia. Plants 2021, 10, 17. [Google Scholar] [CrossRef]
- Vega, A.; Gutiérrez, R.A.; Peña-Neira, A.; Cramer, G.R.; Arce-Johnson, P. Compatible GLRaV-3 viral infections affect berry ripening decreasing sugar accumulation and anthocyanin biosynthesis in Vitis vinifera. Plant Mol. Biol. 2011, 77, 261–274. [Google Scholar] [CrossRef]
- Mannini, F.; Mollo, A.; Credi, R. Field performance and wine quality modification in a clone of Vitis vinifera cv. dolcetto after GLRaV-3 elimination. Am. J. Enol. Vitic. 2012, 63, 144–147. [Google Scholar] [CrossRef] [Green Version]
- Czotter, N.; Molnar, J.; Szabó, E.; Demian, E.; Kontra, L.; Baksa, I.; Szittya, G.; Kocsis, L.; Deak, T.; Bisztray, G.; et al. NGS of virus-derived small RNAs as a diagnostic method used to determine viromes of Hungarian Vineyards. Front. Microbiol. 2018, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- Hančević, K.; Zdunić, G.; Vončina, D.; Radić, T. Virus composition influences virus elimination success and in vitro growth characteristics of the Grapevine cv. Plavac mali. J. Plant Pathol. 2015, 97, 199–202. [Google Scholar]
- Pesqueira, A.M.; Cabaleiro, C.; Velasco, L. Genetic analysis of Grapevine leafroll-associated virus 3 population from Galicia, Spain. Plant Pathol. 2016, 65, 310–321. [Google Scholar] [CrossRef]
- Gouveia, P.; Santos, M.T.; Eiras-Dias, J.E.; Nolasco, G. Five phylogenetic groups identified in the coat protein gene of grapevine leafroll-associated virus 3 obtained from Portuguese grapevine varieties. Arch. Virol. 2011, 156, 413–420. [Google Scholar] [CrossRef]
- Farooq, A.B.U.; Ma, Y.; Wang, Z.; Zhuo, N.; Wenxing, X.; Wang, G.; Hong, N. Genetic diversity analyses reveal novel recombination events in Grapevine leafroll-associated virus 3 in China. Virus Res. 2013, 171, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Gambino, G.; Gribaudo, I. Simultaneous detection of nine grapevine viruses by multiplex reverse transcription-polymerase chain reaction with coamplification of a plant RNA as internal control. Phytopathology 2006, 96, 1223–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afechtal, M.; Mounir, M.; Minafra, A.; Saldarelli, P.; Kubaa, R.A. First report on the occurence of grapevine rupestris stem pitting-associated virus in Moroccan grapevines. J. Plant Pathol. 2019, 101, 405. [Google Scholar] [CrossRef] [Green Version]
- Candresse, T.; Theil, S.; Faure, C.; Marais, A. Determination of the complete genomic sequence of grapevine virus H, a novel vitivirus infecting grapevine. Arch. Virol. 2018, 163, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Panailidou, P.; Lotos, L.; Sassalou, C.-L.; Gagiano, E.; Pietersen, G.; Katis, N.; Maliogka, V.I. First report of grapevine virus H (GVH) in grapevine in Greece. Plant Dis. 2021, 105, 2738. [Google Scholar] [CrossRef]
- Jagunić, M.; Lazarević, B.; Nikolić, K.; Stupić, D.; Preiner, D.; Vončina, D. Detection, transmission, and characterization of grapevine virus H in Croatia. Pathogens 2021, 10, 1578. [Google Scholar] [CrossRef]
- Malagnini, V.; de Lillo, E.; Saldarelli, P.; Beber, R.; Duso, C.; Raiola, A.; Zanotelli, L.; Valenzano, D.; Giampetruzzi, A.; Morelli, M.; et al. Transmission of grapevine Pinot gris virus by Colomerus vitis (Acari: Eriophyidae) to grapevine. Arch. Virol. 2016, 161, 2595–2599. [Google Scholar] [CrossRef]
- Hily, J.M.; Poulicard, N.; Candresse, T.; Vigne, E.; Beuve, M.; Renault, L.; Velt, A.; Spilmont, A.S.; Lemaire, O. Datamining, Genetic Diversity Analyses, and Phylogeographic Reconstructions Redefine the Worldwide Evolutionary History of Grapevine Pinot gris virus and Grapevine berry inner necrosis virus. Phytobiomes J. 2020, 4, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Bertazzon, N.; Forte, V.; Filippin, L.; Causin, R.; Maixner, M.; Angelini, E. Association between genetic variability and titre of Grapevine Pinot gris virus with disease symptoms. Plant Pathol. 2017, 66, 949–959. [Google Scholar] [CrossRef]
- MacKenzie, D.J.; McLean, M.A.; Mukerji, S.; Green, M. Improved RNA extraction from woody plants for the detection of viral pathogens by reverse transcription-polymerase chain reaction. Plant Dis. 1997, 81, 222–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morelli, M.; de Catarino, A.M.; Susca, L.; Saldarelli, P.; Gualandri, V.; Martelli, G.P. First report of Grapevine pinot gris virus from table grapes in southern Italy. J. Plant Pathol. 2014, 96, 439. [Google Scholar]
- Al Rwahnih, M.; Dave, A.; Anderson, M.M.; Rowhani, A.; Uyemoto, J.K.; Sudarshana, M.R. Association of a DNA virus with grapevines affected by red blotch disease in California. Phytopathology 2013, 103, 1069–1076. [Google Scholar] [CrossRef] [Green Version]
- Černi, S.; Ruščić, J.; Nolasco, G.; Gatin, Ž.; Krajačić, M.; Škorić, D. Stem pitting and seedling yellows symptoms of Citrus tristeza virus infection may be determined by minor sequence variants. Virus Genes 2008, 36, 241–249. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vine-Growing Area | Clone | * GFLV | ArMV | GLRaV-1 | GLRaV-2 | GLRaV-3 | GFkV | GVA | GVB | GVH | GRSPaV | GPGV | GBV-1 | GLD Symptoms 2020. | GLD Symptoms 2021. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Šolta | 004/3 | + | + | + | + | AS | MI | ||||||||
| Čiovo | 008/1 | + | + | + | + | AS | MO | ||||||||
| Hvar | 010/1 | + | + | + | + | MI | MI | ||||||||
| 020/1 | + | + | + | + | + | + | S | - | |||||||
| 020/2 | + | + | + | + | + | + | S | - | |||||||
| 026/1 | + | + | + | + | + | AS | AS | ||||||||
| 026/3 | + | + | + | + | AS | AS | |||||||||
| 027/3 | + | + | AS | AS | |||||||||||
| 027/4 | + | + | + | AS | MI | ||||||||||
| Zemunik | 036/1 | + | + | + | + | + | + | MO | MI | ||||||
| 036/3 | + | + | + | + | MO | MI | |||||||||
| Svinišće | 041/1 | + | + | + | + | MI | MI | ||||||||
| Labin | 043/1 | + | + | + | + | + | + | MO | AS | ||||||
| Pelješac | 056/1 | + | + | MI | AS | ||||||||||
| 059/3 | + | + | + | + | + | MO | - | ||||||||
| Korčula | 091/1 | + | + | + | MO | MI | |||||||||
| 095/1 | + | + | AS | AS | |||||||||||
| 096/4 | + | + | + | + | MO | MI | |||||||||
| 096/5 | + | + | MO | MI | |||||||||||
| Vrgorac | 202/3 | + | + | + | AS | MI | |||||||||
| Vis | 210/1 | + | + | + | + | MO | MI | ||||||||
| 210/5 | + | + | MO | MI | |||||||||||
| 214/1 | + | + | + | + | + | + | MO | MI | |||||||
| 214/2 | + | + | + | + | + | + | MO | MI | |||||||
| 215/1 | + | + | + | + | MI | MI | |||||||||
| 215/3 | + | + | + | + | + | MI | MI | ||||||||
| 217/1 | + | AS | MI | ||||||||||||
| 217/2 | + | AS | MI | ||||||||||||
| 217/3 | + | + | AS | MI | |||||||||||
| 217/4 | + | + | + | MI | MI | ||||||||||
| 217/5 | + | + | + | + | MI | MI | |||||||||
| Kaštela | 225/3 | + | + | + | + | + | MI | MI | |||||||
| Brač | 268/1 | + | + | + | + | + | + | S | AS | ||||||
| 268/2 | + | + | + | S | AS | ||||||||||
| Trogir | TG | + | + | + | + | MO | MO |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čarija, M.; Radić, T.; Černi, S.; Mucalo, A.; Zdunić, G.; Vončina, D.; Jagunić, M.; Hančević, K. Prevalence of Virus Infections and GLRaV-3 Genetic Diversity in Selected Clones of Croatian Indigenous Grapevine Cultivar Plavac Mali. Pathogens 2022, 11, 176. https://doi.org/10.3390/pathogens11020176
Čarija M, Radić T, Černi S, Mucalo A, Zdunić G, Vončina D, Jagunić M, Hančević K. Prevalence of Virus Infections and GLRaV-3 Genetic Diversity in Selected Clones of Croatian Indigenous Grapevine Cultivar Plavac Mali. Pathogens. 2022; 11(2):176. https://doi.org/10.3390/pathogens11020176
Chicago/Turabian StyleČarija, Mate, Tomislav Radić, Silvija Černi, Ana Mucalo, Goran Zdunić, Darko Vončina, Martin Jagunić, and Katarina Hančević. 2022. "Prevalence of Virus Infections and GLRaV-3 Genetic Diversity in Selected Clones of Croatian Indigenous Grapevine Cultivar Plavac Mali" Pathogens 11, no. 2: 176. https://doi.org/10.3390/pathogens11020176
APA StyleČarija, M., Radić, T., Černi, S., Mucalo, A., Zdunić, G., Vončina, D., Jagunić, M., & Hančević, K. (2022). Prevalence of Virus Infections and GLRaV-3 Genetic Diversity in Selected Clones of Croatian Indigenous Grapevine Cultivar Plavac Mali. Pathogens, 11(2), 176. https://doi.org/10.3390/pathogens11020176