
Analysis of the Effects of Food Additives on Porphyromonas gingivalis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
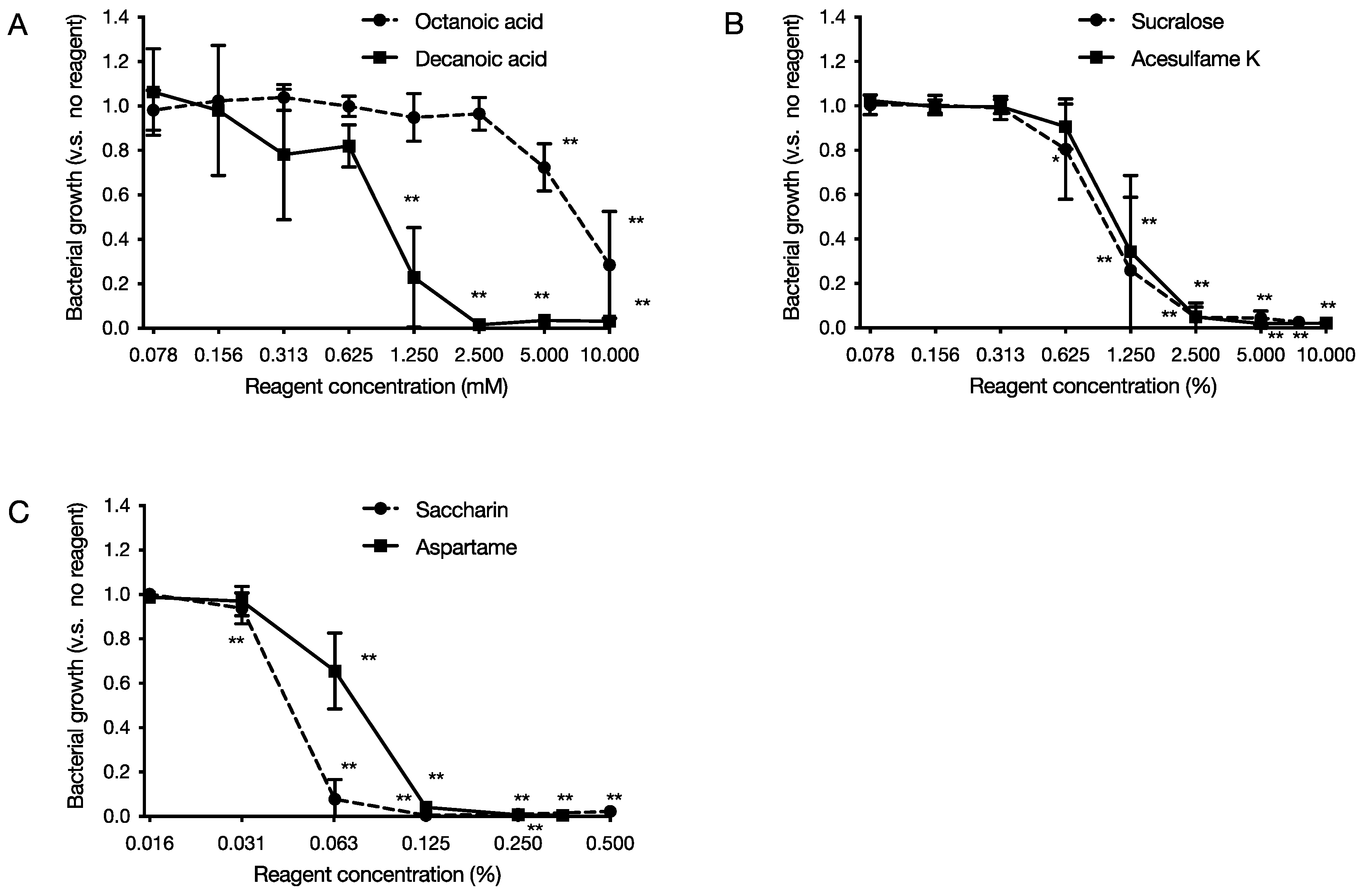
2.1. Growth Inhibition Effect of Food Additives
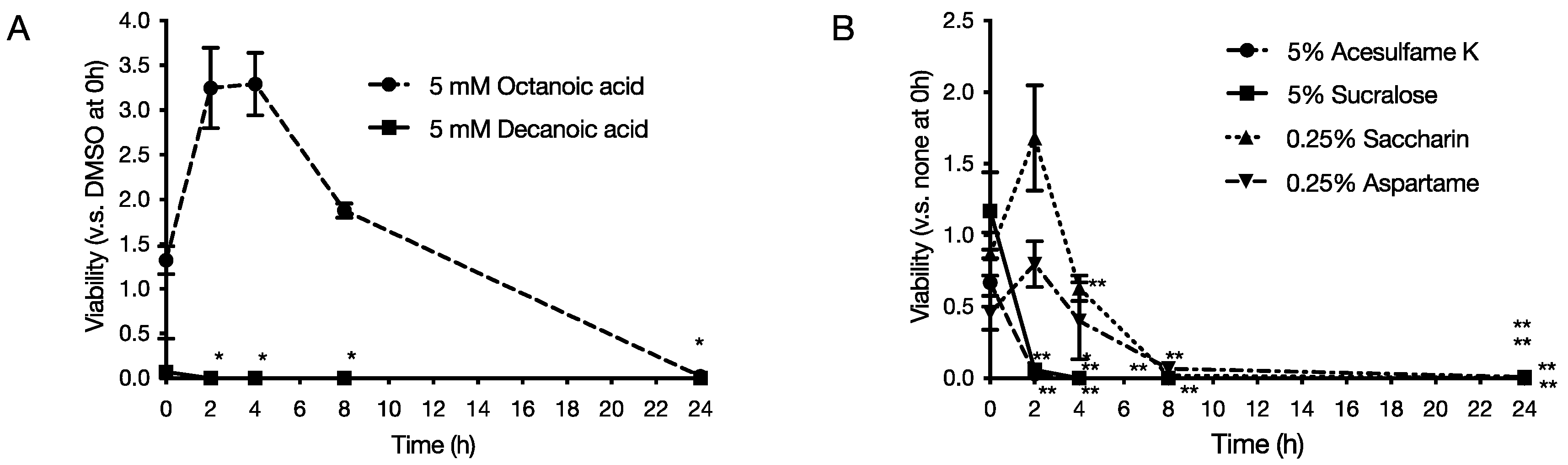
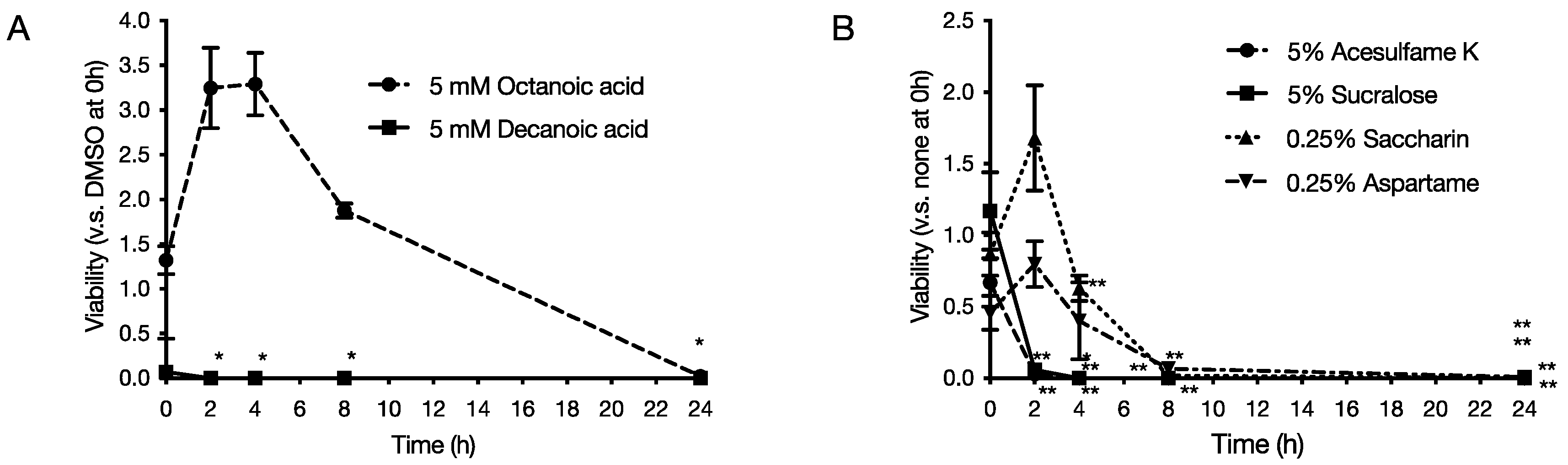
2.2. Bactericidal Activity of Food Additives
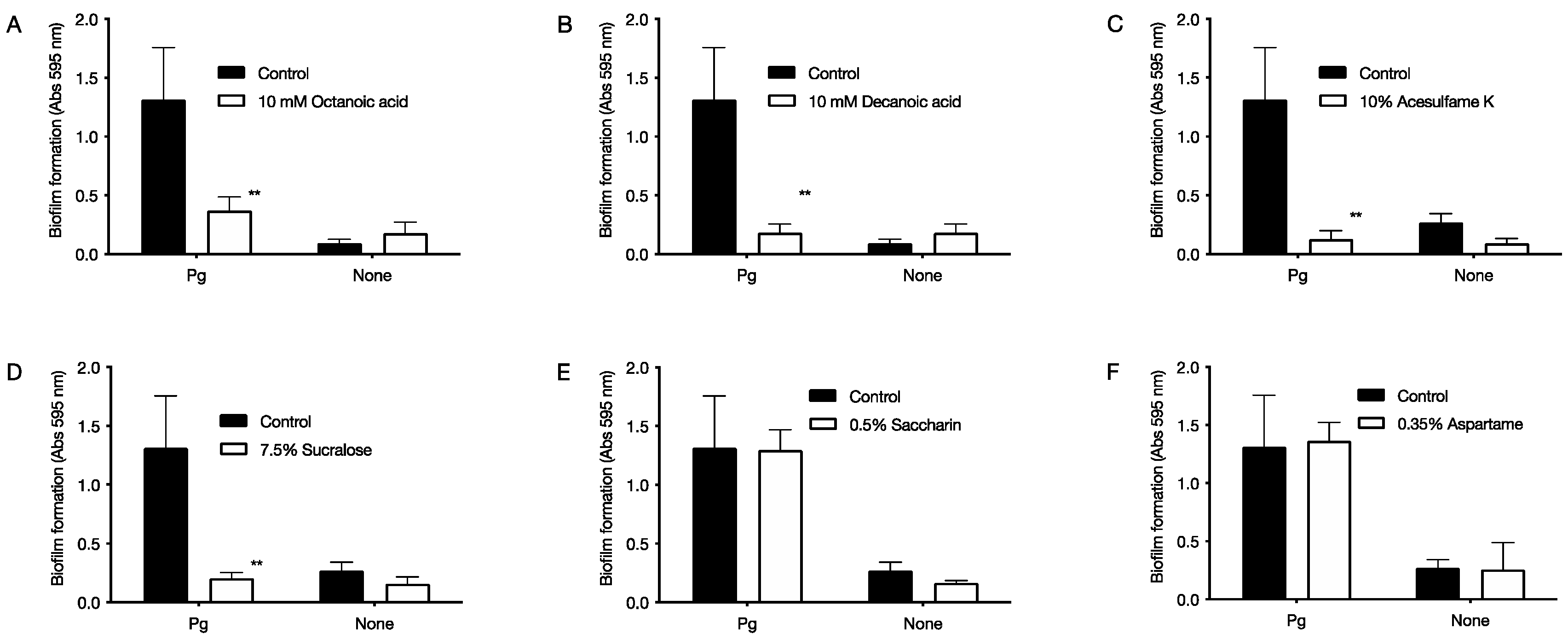
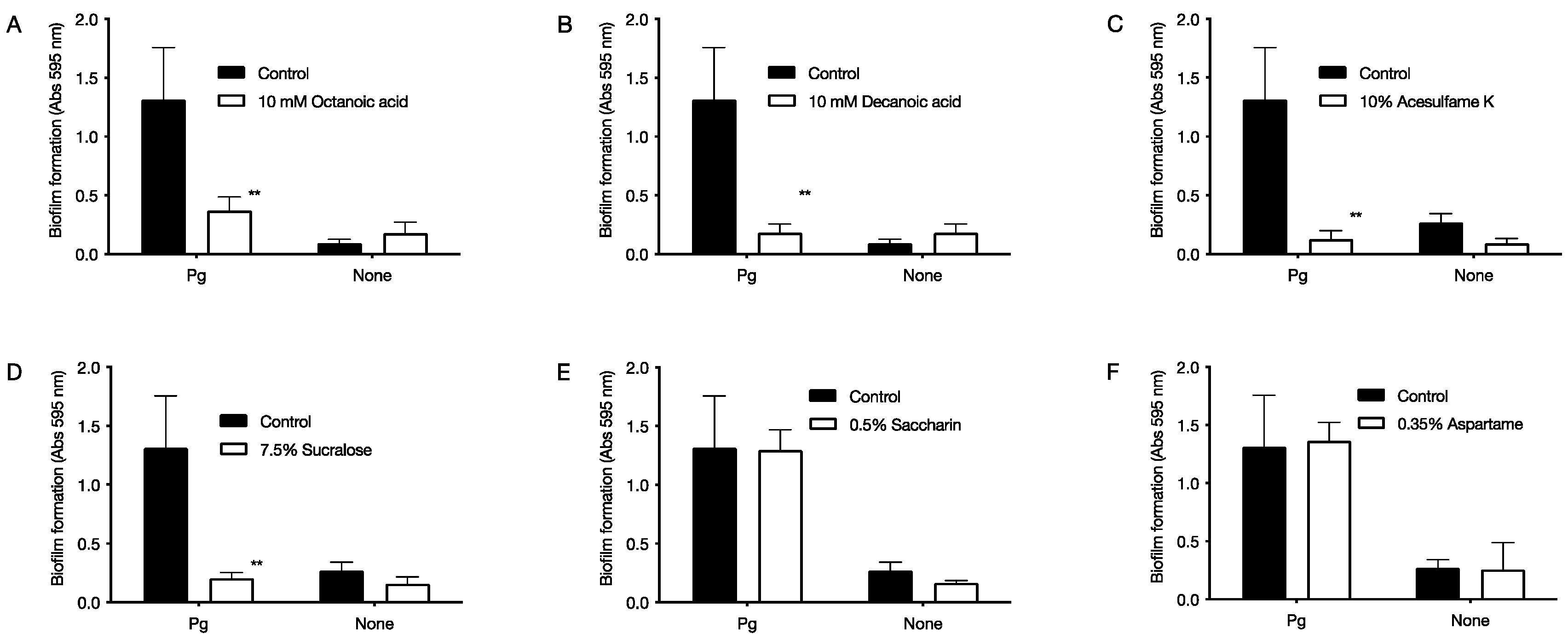
2.3. Inhibitory Effect of Food Additives on Biofilm Formation
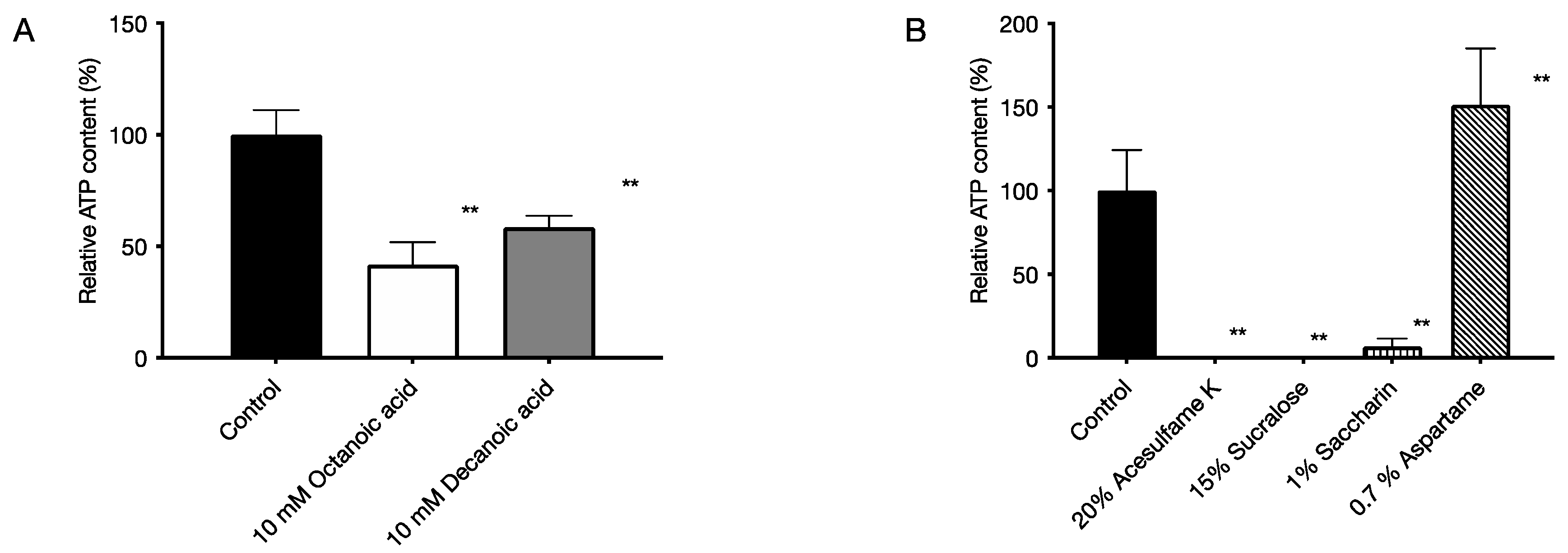
2.4. Bactericidal Activity of Food Additives against Bacteria in Biofilms
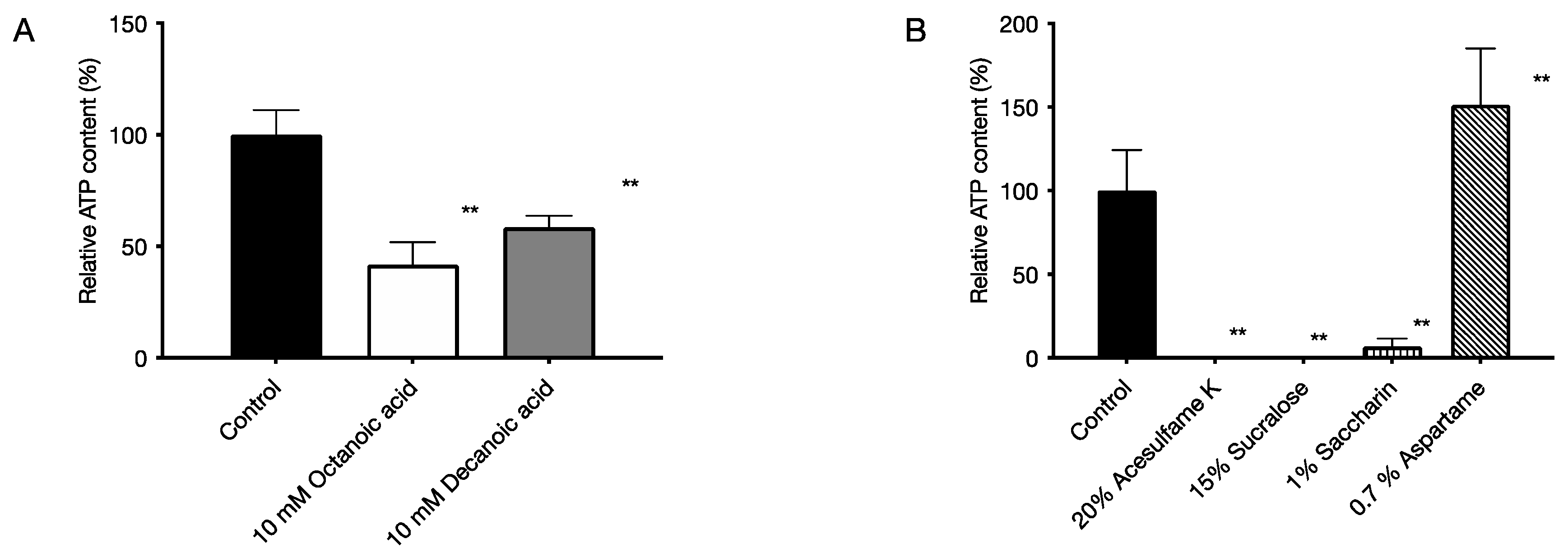
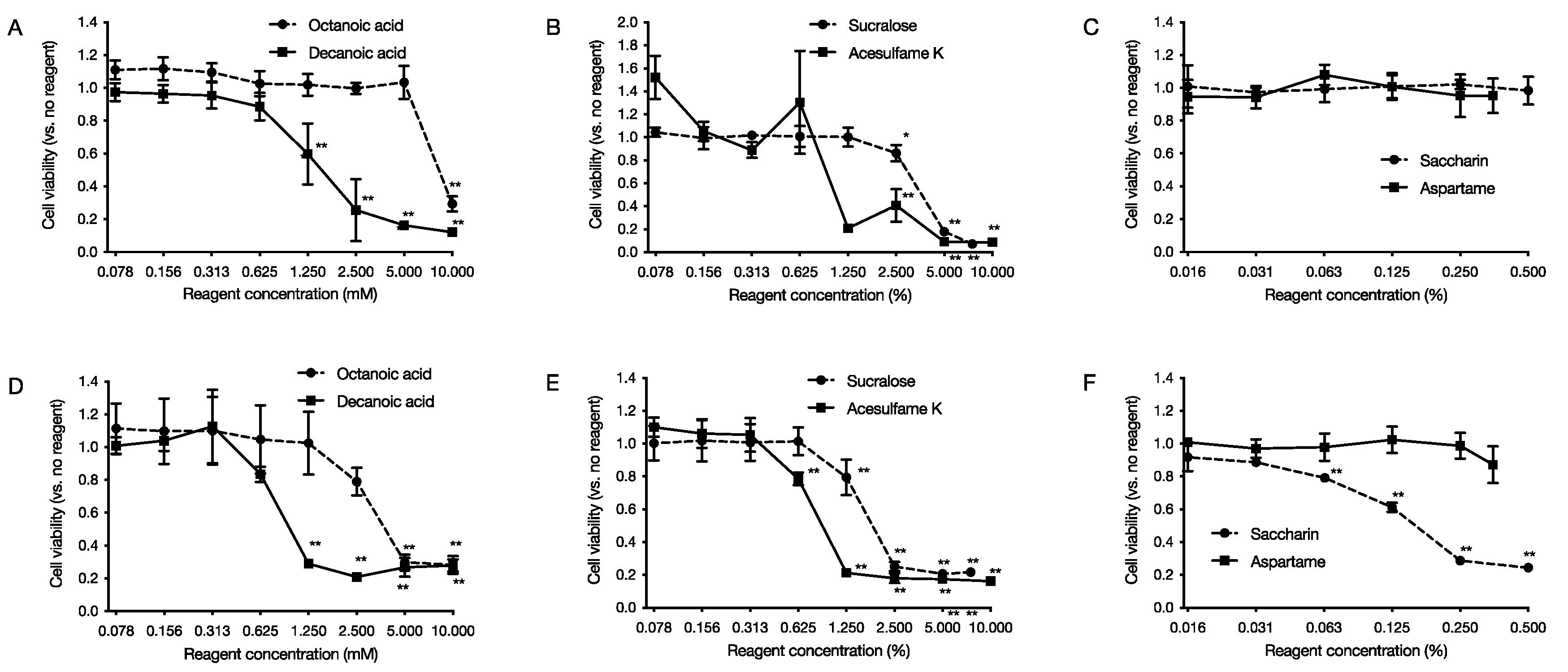
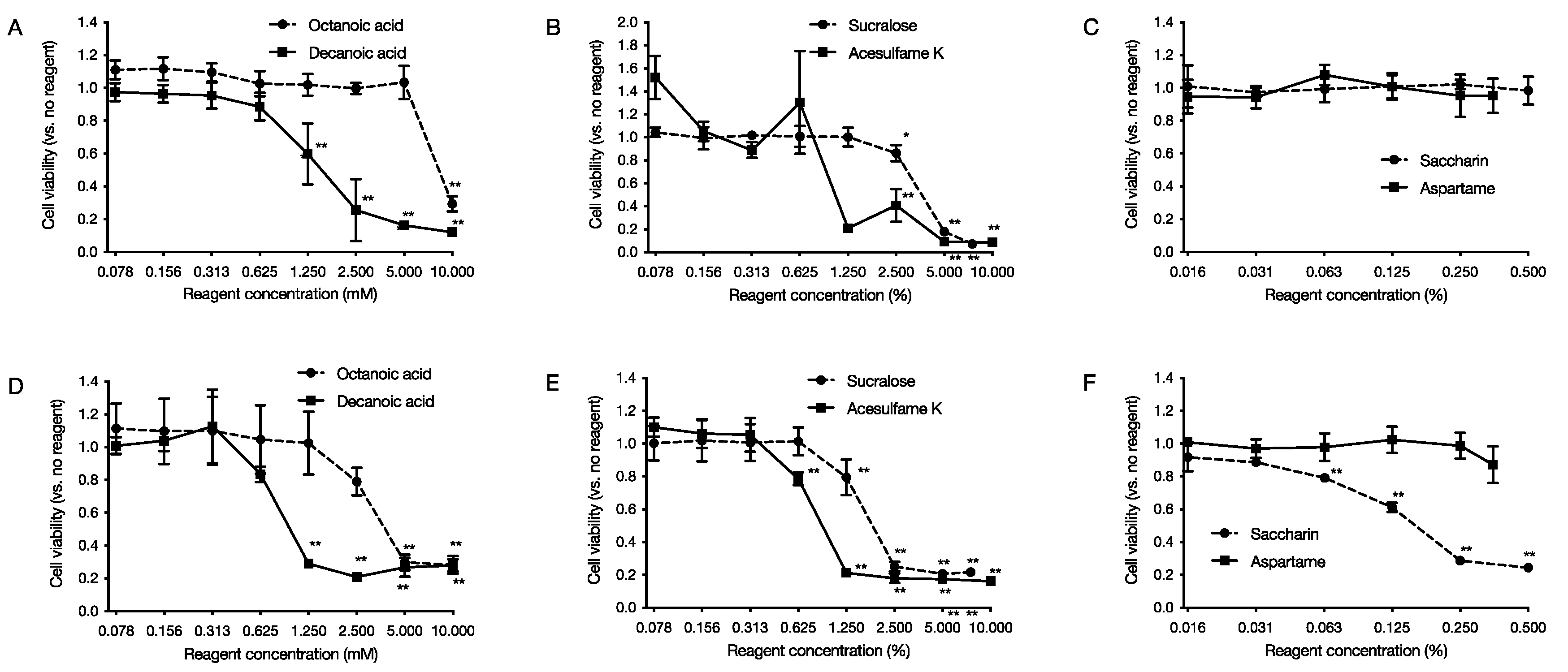
2.5. Cytotoxic Effects on Human-Derived Cells
3. Discussion
4. Materials and Methods
4.1. Bacterial Culture
4.2. Food Additives
4.3. Determination of Minimum Inhibitory Concentration (MIC)
4.4. Time-Kill Assay
4.5. Inhibition of Biofilm Formation
4.6. Bactericidal Test against Bacteria in Biofilm
4.7. Cytotoxicity Test
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Genco, R.J.; Sanz, M. Clinical and Public Health Implications of Periodontal and Systemic Diseases: An Overview. Periodontol. 2000 2020, 83, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Chavakis, T. Local and Systemic Mechanisms Linking Periodontal Disease and Inflammatory Comorbidities. Nat. Rev. Immunol. 2021, 21, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G. Periodontitis: From Microbial Immune Subversion to Systemic Inflammation. Nat. Rev. Immunol. 2014, 15, 30–44. [Google Scholar] [CrossRef]
- Vos, T.; Barber, R.M.; Bell, B.; Bertozzi-Villa, A.; Biryukov, S.; Bolliger, I.; Charlson, F.; Davis, A.; Degenhardt, L.; Dicker, D.; et al. Global, Regional, and National Incidence, Prevalence, and Years Lived with Disability for 301 Acute and Chronic Diseases and Injuries in 188 Countries, 1990–2013: A Systematic Analysis for the Global Burden of Disease Study 2013. Lancet 2015, 386, 743–800. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, A.B.; Crean, S.; Olsen, I.; Singhrao, S.K. Periodontitis, Microbiomes and Their Role in Alzheimer’s Disease. Front. Aging Neurosci. 2017, 9, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elwishahy, A.; Antia, K.; Bhusari, S.; Ilechukwu, N.C.; Horstick, O.; Winkler, V. Porphyromonas Gingivalis as a Risk Factor to Alzheimer’s Disease: A Systematic Review. J. Alzheimer’s Dis. Rep. 2021, 5, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Leech, M.T.; Bartold, P.M. The Association between Rheumatoid Arthritis and Periodontitis. Best Pract. Res. Clin. Rheumatol. 2015, 29, 189–201. [Google Scholar] [CrossRef]
- Geurs, N.C.; Lewis, C.E.; Jeffcoat, M.K. Osteoporosis and Periodontal Disease Progression. Periodontol. 2000 2003, 32, 105–110. [Google Scholar] [CrossRef]
- Terpenning, M.S.; Taylor, G.W.; Lopatin, D.E.; Kerr, C.K.; Dominguez, B.L.; Loesche, W.J. Aspiration Pneumonia: Dental and Oral Risk Factors in an Older Veteran Population. J. Am. Geriatr. Soc. 2001, 49, 557–563. [Google Scholar] [CrossRef]
- Pace, C.C.; McCullough, G.H. The Association between Oral Microorgansims and Aspiration Pneumonia in the Institutionalized Elderly: Review and Recommendations. Dysphagia 2010, 25, 307–322. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Zhang, W.; Liu, X.; Zhang, W.; Li, Y. Interrelationship between Diabetes and Periodontitis: Role of Hyperlipidemia. Arch. Oral Biol. 2015, 60, 667–674. [Google Scholar] [CrossRef]
- Zardawi, F.; Gul, S.; Abdulkareem, A.; Sha, A.; Yates, J. Association Between Periodontal Disease and Atherosclerotic Cardiovascular Diseases: Revisited. Front. Cardiovasc. Med. 2021, 7, 625579. [Google Scholar] [CrossRef]
- Verma, D.; Garg, P.K.; Dubey, A.K. Insights into the Human Oral Microbiome. Arch. Microbiol. 2018, 200, 525–540. [Google Scholar] [CrossRef] [PubMed]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L. Microbial Complexes in Subgingival Plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef]
- Zenobia, C.; Hajishengallis, G. Porphyromonas Gingivalis Virulence Factors Involved in Subversion of Leukocytes and Microbial Dysbiosis. Virulence 2015, 6, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Olsen, I.; Lambris, J.D.; Hajishengallis, G. Porphyromonas Gingivalis Disturbs Host–Commensal Homeostasis by Changing Complement Function. J. Oral Microbiol. 2017, 9, 1340085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popa, C.; Filioreanu, A.M.; Stelea, C.; Maftei, G.A.; Popescu, E. Prevalence of oral lesions modulated by patient’s age: The young versus the elderly. Rom. J. Oral Rehabil. 2018, 10, 50–56. [Google Scholar]
- Liu, B.; Dion, M.R.; Jurasic, M.M.; Gibson, G.; Jones, J.A. Xerostomia and Salivary Hypofunction in Vulnerable Elders: Prevalence and Etiology. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2012, 114, 52–60. [Google Scholar] [CrossRef]
- Burcham, Z.M.; Garneau, N.L.; Comstock, S.S.; Tucker, R.M.; Knight, R.; Metcalf, J.L.; Miranda, A.; Reinhart, B.; Meyers, D.; Woltkamp, D.; et al. Patterns of Oral Microbiota Diversity in Adults and Children: A Crowdsourced Population Study. Sci. Rep. 2020, 10, 2133. [Google Scholar] [CrossRef] [Green Version]
- Mann, J.; Bernstein, Y.; Findler, M. Periodontal Disease and Its Prevention, by Traditional and New Avenues. Exp. Ther. Med. 2020, 19, 1504–1506. [Google Scholar] [CrossRef] [PubMed]
- Martinon, P.; Fraticelli, L.; Giboreau, A.; Dussart, C.; Bourgeois, D.; Carrouel, F. Nutrition as a Key Modifiable Factor for Periodontitis and Main Chronic Diseases. J. Clin. Med. 2021, 10, 197. [Google Scholar] [CrossRef] [PubMed]
- Najeeb, S.; Zafar, M.S.; Khurshid, Z.; Zohaib, S.; Almas, K. The Role of Nutrition in Periodontal Health: An Update. Nutrients 2016, 8, 530. [Google Scholar] [CrossRef] [PubMed]
- Hujoel, P. Dietary Carbohydrates and Dental-Systemic Diseases. J. Dent. Res. 2009, 88, 490–502. [Google Scholar] [CrossRef]
- Prashant, G.; Patil, R.B.; Nagaraj, T.; Patel, V.B. The Antimicrobial Activity of the Three Commercially Available Intense Sweeteners against Common Periodontal Pathogens: An in Vitro Study. J. Contemp. Dent. Pract. 2012, 13, 749–752. [Google Scholar] [CrossRef]
- Berg, T.; Licht, D. International Legislation on Trace Elements as Contaminants in Food: A Review. Food Addit. Contam. 2002, 19, 916–927. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How Antibiotics Kill Bacteria: From Targets to Networks. Nat. Rev. Microbiol. 2010, 8, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Shah, H.N.; Williams, R.A.D. Utilization of Glucose and Amino Acids ByBacteroides Intermedius AndBacteroides Gingivalis. Curr. Microbiol. 1987, 15, 241–246. [Google Scholar] [CrossRef]
- Martu, M.-A.; Maftei, G.-A.; Luchian, I.; Popa, C.; Filioreanu, A.-M.; Tatarciuc, D.; Nichitean, G.; Hurjui, L.-L.; Foia, L.-G. Wound healing of periodontal and oral tissues: Part ii—Patho-phisiological conditions and metabolic diseases. review. Rom. J. Oral Rehabil. 2020, 12, 30–40. [Google Scholar]
- Suárez, L.J.; Arboleda, S.; Angelov, N.; Arce, R.M. Oral Versus Gastrointestinal Mucosal Immune Niches in Homeostasis and Allostasis. Front. Immunol. 2021, 12, 705206. [Google Scholar] [CrossRef]
- Altun, E.; Walther, C.; Borof, K.; Petersen, E.; Lieske, B.; Kasapoudis, D.; Jalilvand, N.; Beikler, T.; Jagemann, B.; Zyriax, B.-C.; et al. Association between Dietary Pattern and Periodontitis—A Cross-Sectional Study. Nutrients 2021, 13, 4167. [Google Scholar] [CrossRef]
- Huang, W.-C.; Tsai, T.-H.; Chuang, L.-T.; Li, Y.-Y.; Zouboulis, C.C.; Tsai, P.-J. Anti-Bacterial and Anti-Inflammatory Properties of Capric Acid against Propionibacterium Acnes: A Comparative Study with Lauric Acid. J. Dermatol. Sci. 2014, 73, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Petschow, B.W.; Batema, R.P.; Ford, L.L. Susceptibility of Helicobacter Pylori to Bactericidal Properties of Medium-Chain Monoglycerides and Free Fatty Acids. Antimicrob. Agents Chemother. 1996, 40, 302–306. [Google Scholar] [CrossRef] [Green Version]
- Ramanarayanan, S.; Mittal, S.; Hiregoudar, M.; Basha, S.; Manjunath, P.G.; Natraj, C.G. Antifungal Activity of Four Commercially Available Intense Sweeteners against Candida Albicans—An In Vitro Study. Univ. Res. J. Dent. 2013, 3, 60–63. [Google Scholar] [CrossRef]
- Subramaniam, R.; Mittal, S.; Hiregoudar, M.; Ratan, G.N.; Sakeenabi, B.; Prashant, G.M.; Chandu, G.N. Antimicrobial Activity of Stevioside on Periodontal Pathogens and Candida Albicans—An Invitro Study. J. Indian Assoc. Public Health Dent. 2011, 2011, 325–329. [Google Scholar]
- Hashino, E.; Kuboniwa, M.; Alghamdi, S.A.; Yamaguchi, M.; Yamamoto, R.; Cho, H.; Amano, A. Erythritol Alters Microstructure and Metabolomic Profiles of Biofilm Composed of Streptococcus Gordonii and Porphyromonas Gingivalis. Mol. Oral Microbiol. 2013, 28, 435–451. [Google Scholar] [CrossRef]
- Asahi, Y.; Noiri, Y.; Miura, J.; Maezono, H.; Yamaguchi, M.; Yamamoto, R.; Azakami, H.; Hayashi, M.; Ebisu, S. Effects of the Tea Catechin Epigallocatechin Gallate on Porphyromonas Gingivalis Biofilms. J. Appl. Microbiol. 2014, 116, 1164–1171. [Google Scholar] [CrossRef]
- Karched, M.; Bhardwaj, R.G.; Inbamani, A.; Asikainen, S. Quantitation of Biofilm and Planktonic Life Forms of Coexisting Periodontal Species. Anaerobe 2015, 35, 13–20. [Google Scholar] [CrossRef]
- Scherer, W.F.; Syverton, J.T.; Gey, G.O. Studies on the propagation in vitro of poliomyelitis viruses. J. Exp. Med. 1953, 97, 695–710. [Google Scholar] [CrossRef] [Green Version]
- Drexler, H.G.; Gaedicke, G.; Minowada, J. T-Leukemia Cell Lines CCRF-CEM, HPB-ALL, JM and MOLT-4: Changes in Isoenzyme Profiles during Induction of Differentiation. Ann. Hematol. 1987, 54, 79–87. [Google Scholar] [CrossRef]
- Malaekeh-Nikouei, B.; Mousavi, S.H.; Shahsavand, S.; Mehri, S.; Nassirli, H.; Moallem, S.A. Assessment of Cytotoxic Properties of Safranal and Nanoliposomal Safranal in Various Cancer Cell Lines. Phytother. Res. 2013, 27, 1868–1873. [Google Scholar] [CrossRef]
- Ghoran, S.H.; Rahimi, H.; Kazemi, A.; Scognamiglio, M.; Naderian, M.; Iraji, A.; Bordbar, F. Allium Hooshidaryae (Alliaceae); Chemical Compositions, Biological and Ethnomedicine Uses. J. Ethnopharmacol. 2021, 274, 113918. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, H.; Ishiyama, M.; Ohseto, F.; Sasamoto, K.; Hamamoto, T.; Suzuki, K.; Watanabe, M. A Water-Soluble Tetrazolium Salt Useful for Colorimetric Cell Viability Assay. Anal. Commun. 1999, 36, 47–50. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shinohara, M.; Maetani, M.; Kitada, C.; Nishigami, Y.; Yazawa, A.; Kamitani, S. Analysis of the Effects of Food Additives on Porphyromonas gingivalis. Pathogens 2022, 11, 65. https://doi.org/10.3390/pathogens11010065
Shinohara M, Maetani M, Kitada C, Nishigami Y, Yazawa A, Kamitani S. Analysis of the Effects of Food Additives on Porphyromonas gingivalis. Pathogens. 2022; 11(1):65. https://doi.org/10.3390/pathogens11010065
Chicago/Turabian StyleShinohara, Mai, Miki Maetani, Chiharu Kitada, Yasuko Nishigami, Ayaka Yazawa, and Shigeki Kamitani. 2022. "Analysis of the Effects of Food Additives on Porphyromonas gingivalis" Pathogens 11, no. 1: 65. https://doi.org/10.3390/pathogens11010065
APA StyleShinohara, M., Maetani, M., Kitada, C., Nishigami, Y., Yazawa, A., & Kamitani, S. (2022). Analysis of the Effects of Food Additives on Porphyromonas gingivalis. Pathogens, 11(1), 65. https://doi.org/10.3390/pathogens11010065