Sterile but Sexy: Assessing the Mating Competitiveness of Irradiated Bagrada hilaris Males for the Development of a Sterile Insect Technique

 , , , ,
, , , ,  ,
, 
 and
and 
Simple Summary
Abstract
1. Introduction
- (a)
- Evaluation of the sexual performance between males irradiated at three different doses (60, 80, and 100 Gy) and non-irradiated males (control), in terms of the amount of time taken to initiate the first copulation, as well as the number and duration of matings over the 18 h of the bioassay.
- (b)
- Study of sexual behavior at the optimal dose: once the first experiment was completed, we evaluated differences in the mating success of irradiated and non-irradiated males under both no-choice and choice conditions. In addition, differences in feeding activity between irradiated males, non-irradiated males, and females were assessed. Unlike the first bioassay, observations were carried out for three days, allowing us to assess the variation in the performance of the irradiated males over time.
2. Materials and Methods
2.1. Insect Collection and Rearing
2.2. Irradiation Process
2.3. Evaluation of Sexual Performance at Three Different Doses
2.4. Study of Sexual Behavior at the Optimal Dose
2.5. Data Analysis
2.5.1. Evaluation of Sexual Performance at Three Different Doses
2.5.2. Study of Sexual Behavior at the Optimal Dose
3. Results
3.1. Evaluation of Sexual Performance at Three Different Doses
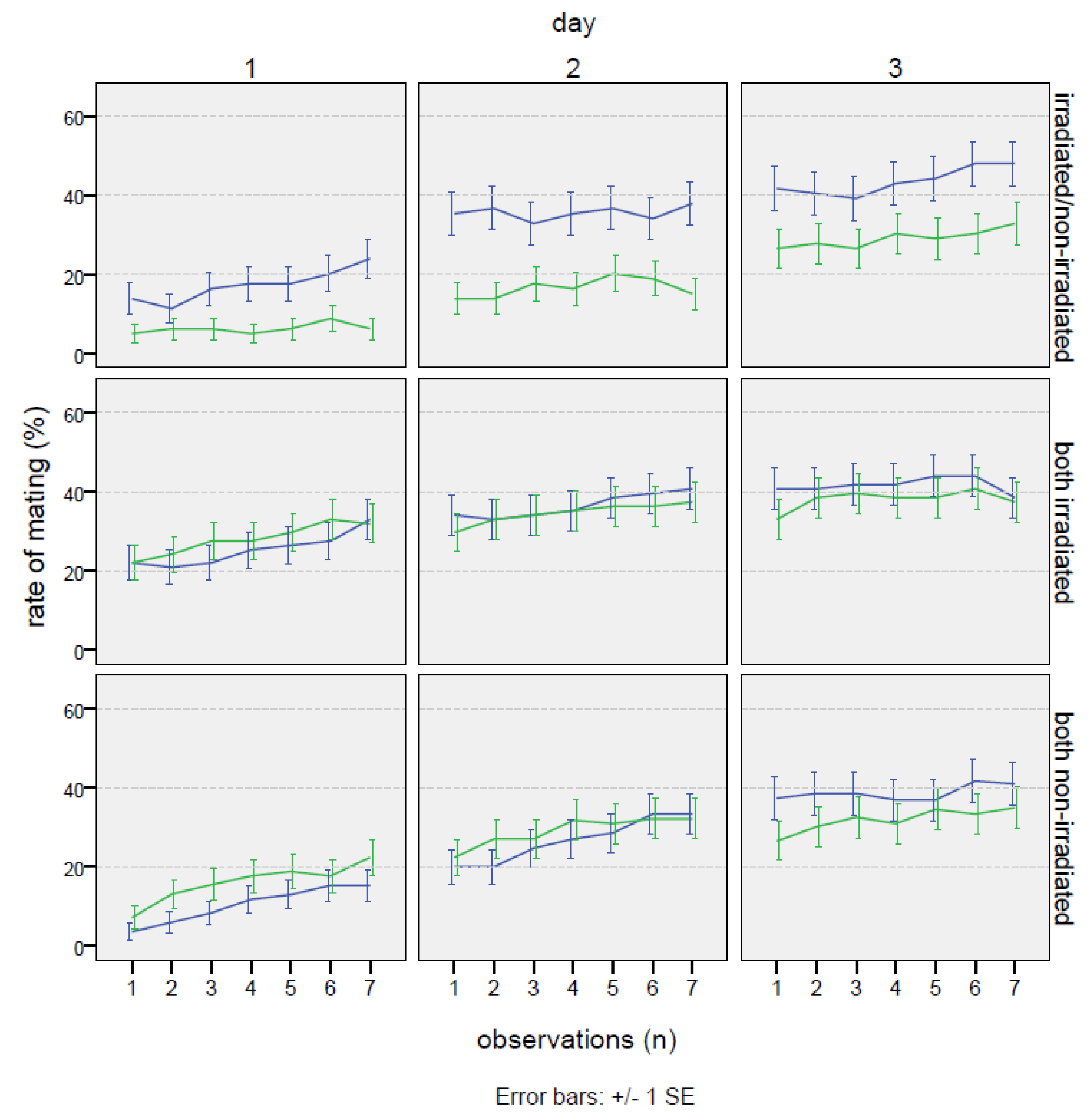
3.2. Study of Sexual Behavior at the Optimal Dose
3.2.1. No-Choice Test
3.2.2. Choice Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fixed Effects | Estimate | ±s.e. | Wald | p-Value |
|---|---|---|---|---|
| (intercept) | −0.800 | 0.110 | 52.5 | 4.4 × 10−13 *** |
| irradiated | 0.127 | 0.144 | 0.38 | 0.78 |
| poly(observations) 1 | 30.251 | 3.762 | 64.7 | 8.9 × 10−16 *** |
| poly(observations) 2 | −16.125 | 3.799 | 18.01 | 2.2 × 10−5 *** |
| poly(observations) 3 | 12.971 | 2.522 | 26.45 | 2.7 × 10−7 *** |
| Fixed Effects | Estimate | ±s.e. | Wald | p-Value |
|---|---|---|---|---|
| (intercept) | −0.252 | 0.103 | 6.05 | 0.014 * |
| irradiated | 0.202 | 0.141 | 2.05 | 0.152 |
| poly(observations) 1 | 20.021 | 3.942 | 25.80 | 3.8 × 10−7 *** |
| poly(observations) 2 | −22.088 | 3.235 | 46.63 | 8.6 × 10−12 *** |
| poly(observations) 3 | 10.401 | 2.203 | 22.30 | 2.3 × 10−6 *** |

| Fixed Effects | Estimate | ±s.e. | Wald | p-Value |
|---|---|---|---|---|
| (Intercept) | 0.1085 | 0.02301 | 22.24 | 2.4 × 10−6 *** |
| Irradiated male | −0.0062 | 0.02360 | 0.07 | 0.793 |
| Day 2 | 0.0059 | 0.02324 | 0.06 | 0.799 |
| Day 3 | −0.0466 | 0.02072 | 5.05 | 0.025 * |
| Analysis of “Wald statistic” Table | ||||
| df | X2 | p-value | ||
| Treatment | 1 | 0.05 | 0.8193 | |
| Days | 2 | 10.89 | 0.0043 ** | |
| Fixed Effects | Estimate | ±s.e. | Wald | p-Value |
|---|---|---|---|---|
| (Intercept) | −0.982 | 0.188 | 27.41 | 1.6 × 10−7 *** |
| Both non-irradiated | 0.077 | 0.216 | 0.13 | 0.72 |
| Both irradiated | 1.194 | 0.284 | 17.71 | 2.6 × 10−5 *** |
| Day 2 | 1.094 | 0.123 | 78.71 | <2 × 10−16 *** |
| Day 3 | 1.819 | 0.167 | 118.85 | <2 × 10−16 *** |
| Analysis of “Wald statistic” Table | ||||
| df | X2 | p-value | ||
| Treatment | 2 | 19.1 | 7.1 × 10−5 *** | |
| Days | 2 | 133.1 | <2 × 10−16 *** | |
| Comparison | Fixed Effects | Estimate | ±s.e. | Wald | p-Value |
|---|---|---|---|---|---|
| day 1 | 0.0324 | 0.0265 | 1.49 | 0.22 | |
| Irradiated/non-irradiated male | day 2 | 0.0010 | 0.0292 | 0.01 | 0.97 |
| day 3 | −0.0087 | 0.030478 | 0.08 | 0.78 | |
| Test ANOVA | df | X2 | p-value | ||
| days | 3 | 1.81 | 0.61 | ||
| Fixed effects | estimate | ±s.e. | Wald | p-value | |
| Female/non-irradiated male | day 1 | 0.1746 | 0.0298 | 34.3 | 4.8 × 10−9 *** |
| day 2 | 0.1181 | 0.0221 | 28.6 | 9.0 × 10−8 *** | |
| day 3 | 0.0836 | 0.0234 | 12.7 | 0.00036 *** | |
| Test ANOVA | df | X2 | p-value | ||
| days | 3 | 64.8 | 5.7 × 10−14 *** | ||
| Fixed effects | estimate | ±s.e. | Wald | p-value | |
| Female/irradiated male | day 1 | 0.1386 | 0.0255 | 29.6 | 5.2 × 10−8 *** |
| day 2 | 0.1121 | 0.0230 | 23.7 | 1.1 × 10−6 *** | |
| day 3 | 0.0898 | 0.0239 | 14.1 | 0.00018 *** | |
| Test ANOVA | df | X2 | p-value | ||
| days | 3 | 62.5 | 1.8 × 10−13 *** |
References
- Sage, R.F. Global Change Biology: A Primer. Glob. Change Biol. 2020, 26, 3–30. [Google Scholar] [CrossRef] [PubMed]
- Howard, C.W. The Entomological Section: The Bagrada Bug (Bagrada hilaris). Transvaal Agric. J. 1906, 5, 168–176. [Google Scholar]
- Ahuja, B.; Kalyan, R.K.; Ahuja, U.R.; Singh, S.K.; Sundria, M.M.; Dhandapani, A. Integrated Management Strategy for Painted Bug, Bagrada hilaris (Burm) Inflicting Injury at Seedling Stage of Mustard (Brassica juncea) in Arid Western Rajasthan. Pestic. Res. J. 2008, 20, 48–51. [Google Scholar]
- Bundy, C.S.; Grasswitz, T.R.; Sutherland, C. First Report of the Invasive Stink Bug Bagrada hilaris (Burmeister) (Heteroptera: Pentatomidae) from New Mexico, with Notes on Its Biology. Southwest. Entomol. 2012, 37, 411–414. [Google Scholar] [CrossRef]
- Palumbo, J.C.; Natwick, E.T. The Bagrada Bug (Hemiptera: Pentatomidae): A New Invasive Pest of Cole Crops in Arizona and California. Plant Health Prog. 2010, 11, 50. [Google Scholar] [CrossRef]
- Torres-Acosta, R.I.; Sánchez-Peña, S.R. Geographical Distribution of Bagrada hilaris (Hemiptera: Pentatomidae) in Mexico. J. Entomol. Sci. 2016, 51, 165–167. [Google Scholar] [CrossRef]
- Sánchez-Peña, S.R. First Record in Mexico of the Invasive Stink Bug Bagrada hilaris, on Cultivated Crucifers in Saltillo. Southwest. Entomol. 2014, 39, 375–377. [Google Scholar] [CrossRef]
- Palumbo, J.C.; Perring, T.M.; Millar, J.G.; Reed, D.A. Biology, Ecology, and Management of an Invasive Stink Bug, Bagrada hilaris, in North America. Annu. Rev. Entomol. 2016, 61, 453–473. [Google Scholar] [CrossRef]
- Faúndez, E.I. From Agricultural to Household Pest: The Case of the Painted Bug Bagrada hilaris (Burmeister) (Heteroptera: Pentatomidae) in Chile. J. Med. Entomol. 2018, 55, 1365–1368. [Google Scholar] [CrossRef]
- Carpintero, D.L.; Quiroga, V.N.; Celentano, E.; Holgado, M.G. Primer Registro de Bagrada hilaris (Burmeister, 1835)(Hemiptera: Pentatomidae) Para La República Argentina. Hist. Nat. 2021, 11, 179–184. [Google Scholar]
- Carapezza, A. Gli Eterotteri Dell’Isola Di Pantelleria (Insecta, Heteroptera). Il Nat. Sicil. S. 1981, 4, 73–91. [Google Scholar]
- Colazza, S.; Guarino, S. Bagrada hilaris (Burmeister)(Heteroptera: Pentatomidae) a Pest of Capper in the Island of Pantelleria [Capparis spinosa L.; Sicily]. Inf. Fitopatol. 2004, 12, 30–34. [Google Scholar]
- Huang, T.I.; Reed, D.A.; Perring, T.M.; Palumbo, J.C. Feeding Damage by Bagrada hilaris (Hemiptera: Pentatomidae) and Impact on Growth and Chlorophyll Content of Brassicaceous Plant Species. Arthropod-Plant Interact. 2014, 8, 89–100. [Google Scholar] [CrossRef]
- Guarino, S.; Peri, E.; Colazza, S.; Luchi, N.; Michelozzi, M.; Loreto, F. Impact of the Invasive Painted Bug Bagrada hilaris on Physiological Traits of Its Host Brassica oleracea Var Botrytis. Arthropod-Plant Interact. 2017, 11, 649–658. [Google Scholar] [CrossRef]
- Infantino, A.; Tomassoli, L.; Peri, E.; Colazza, S. Viruses, Fungi and Insect Pests Affecting Caper. Eur. J. Plant Sci. Biotechnol. 2007, 1, 170–179. [Google Scholar]
- Guarino, S.; Peri, E.; Lo Bue, P.; La Pillo, A.; Colazza, S. Impiego Di Insetticidi Di Origine Vegetale per Il Contenimento Delle Popolazioni Di Bagrada hilaris Nei Cappereti Panteschi. Inf. Fitopatol. 2007, 7, 53–58. [Google Scholar]
- Hendrichs, J.; Robinson, A. Sterile Insect Technique. In Encyclopedia of Insects; Elsevier: Amsterdam, The Netherlands, 2009; pp. 953–957. ISBN 978-0-12-374144-8. [Google Scholar]
- Ganjisaffar, F.; Talamas, E.J.; Bon, M.C.; Gonzalez, L.; Brown, B.V.; Perring, T.M. Trissolcus hyalinipennis Rajmohana & Narendran (Hymenoptera, Scelionidae), a Parasitoid of Bagrada hilaris (Burmeister) (Hemiptera, Pentatomidae), Emerges in North America. J. Hymenopt. Res. 2018, 65, 111–130. [Google Scholar] [CrossRef]
- Power, N.; Ganjisaffar, F.; Xu, K.; Perring, T.M. Effects of Parasitoid Age, Host Egg Age, and Host Egg Freezing on Reproductive Success of Ooencyrtus mirus (Hymenoptera: Encyrtidae) on Bagrada hilaris (Hemiptera: Pentatomidae) Eggs. Environ. Entomol. 2021, 50, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Martel, G.; Augé, M.; Talamas, E.; Roche, M.; Smith, L.; Sforza, R.F.H. First Laboratory Evaluation of Gryon gonikopalense (Hymenoptera: Scelionidae), as Potential Biological Control Agent of Bagrada hilaris (Hemiptera: Pentatomidae). Biol. Control 2019, 135, 48–56. [Google Scholar] [CrossRef]
- Reed, D.A.; Palumbo, J.C.; Perring, T.M.; May, C. Bagrada hilaris (Hemiptera: Pentatomidae), An Invasive Stink Bug Attacking Cole Crops in the Southwestern United States. J. Integ. Pest. Manag. 2013, 4, 1–7. [Google Scholar] [CrossRef]
- Taylor, M.E.; Bundy, C.S.; McPherson, J.E. Unusual Ovipositional Behavior of the Stink Bug Bagrada hilaris (Hemiptera: Heteroptera: Pentatomidae). Ann. Entomol. Soc. Am. 2014, 107, 872–877. [Google Scholar] [CrossRef]
- Martel, G.; Sforza, R.F.H. Catch Me If You Can: Novel Foraging Behavior of an Egg Parasitoid, Gryon gonikopalense, against the Stinkbug Pest, Bagrada hilaris. J. Pest. Sci. 2021, 94, 1161–1169. [Google Scholar] [CrossRef]
- Knipling, E.F. Possibilities of Insect Control or Eradication Through the Use of Sexually Sterile Males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Tillman, G.; Toews, M.; Blaauw, B.; Sial, A.; Cottrell, T.; Talamas, E.; Buntin, D.; Joseph, S.; Balusu, R.; Fadamiro, H.; et al. Parasitism and Predation of Sentinel Eggs of the Invasive Brown Marmorated Stink Bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae), in the Southeastern US. Biol. Control. 2020, 145, 104247. [Google Scholar] [CrossRef]
- Lance, D.R.; McInnis, D.O. Biological Basis of the Sterile Insect Technique. In Sterile Insect Technique; CRC Press: Boca Raton, FL, USA, 2021; pp. 113–142. [Google Scholar]
- Klassen, W.; Curtis, C.F.; Hendrichs, J. History of the Sterile Insect Technique. In Sterile Insect Technique; CRC Press: Boca Raton, FL, USA, 2021; pp. 1–44. [Google Scholar]
- Welsh, T.J.; Stringer, L.D.; Caldwell, R.; Carpenter, J.E.; Suckling, D.M. Irradiation Biology of Male Brown Marmorated Stink Bugs: Is There Scope for the Sterile Insect Technique? Int. J. Radiat. Biol. 2017, 93, 1357–1363. [Google Scholar] [CrossRef]
- Suckling, D.; Cristofaro, M.; Roselli, G.; Levy, M.; Cemmi, A.; Mazzoni, V.; Stringer, L.; Zeni, V.; Ioriatti, C.; Anfora, G. The Competitive Mating of Irradiated Brown Marmorated Stink Bugs, Halyomorpha halys, for the Sterile Insect Technique. Insects 2019, 10, 411. [Google Scholar] [CrossRef] [PubMed]
- Roselli, G.; Anfora, G.; Suckling, D.M.; Mazzoni, V.; Vanoni, V.; Menegotti, L.; Fellin, L.; Rossi Stacconi, M.V.; Ioriatti, C.; Cristofaro, M. Effects of Irradiation on Biology and Mating Behaviour of Wild Males of Brown Marmorated Stink Bug Using a 6 MV Medical Linear Accelerator. Insects 2023, 14, 460. [Google Scholar] [CrossRef]
- Mau, R.; Mitchell, W.C.; Anwar, M. Preliminary Studies on the Effects of Gamma Irradiation of Eggs and Adults of the Southern Green Stink Bug, Nezara viridula (L.). J. Econ. Entomol. 1967, 19, 415–417. [Google Scholar]
- Dyby, S.D.; Sailer, R.I. Impact of Low-Level Radiation on Fertility and Fecundity of Nezara viridula (Hemiptera: Pentatomidae). J. Econ. Entomol. 1999, 92, 945–953. [Google Scholar] [CrossRef]
- Žunič, A.; Čokl, A.; Serša, G. Effects of 5-Gy Irradiation on Fertility and Mating Behaviour of Nezara viridula (Heteroptera: Pentatomidae). Sterile Insect. Tech. 2002, 36, 231–237. [Google Scholar]
- Cristofaro, M.; Sforza, R.F.H.; Roselli, G.; Paolini, A.; Cemmi, A.; Musmeci, S.; Anfora, G.; Mazzoni, V.; Grodowitz, M. Effects of Gamma Irradiation on the Fecundity, Fertility, and Longevity of the Invasive Stink Bug Pest Bagrada hilaris (Burmeister) (Hemiptera: Pentatomidae). Insects 2022, 13, 787. [Google Scholar] [CrossRef]
- Vreysen, M.J.B.; Abd-Alla, A.M.M.; Bourtzis, K.; Bouyer, J.; Caceres, C.; De Beer, C.; Oliveira Carvalho, D.; Maiga, H.; Mamai, W.; Nikolouli, K.; et al. The Insect Pest Control Laboratory of the Joint FAO/IAEA Programme: Ten Years (2010–2020) of Research and Development, Achievements and Challenges in Support of the Sterile Insect Technique. Insects 2021, 12, 346. [Google Scholar] [CrossRef] [PubMed]
- Itô, Y.; Yamamura, K.; Manoukis, N.C. Role of Population and Behavioural Ecology in the Sterile Insect Technique. In Sterile Insect Technique; CRC Press: Boca Raton, FL, USA, 2021; pp. 245–282. ISBN 978-1-00-303557-2. [Google Scholar]
- Kawada, H.; Kitamura, C. The Reproductive Behavior of the Brown Marmorated Stink Bug, Halyomorpha mista UHLER (Heteroptera: Pentatomidae) I. Observation of Mating Behavior and Multiple Copulation. Appl. Entomol. Zool. 1983, 18, 234–242. [Google Scholar] [CrossRef]
- Zahn, D.K.; Girling, R.D.; McElfresh, J.S.; Cardé, R.T.; Millar, J.G. Biology and Reproductive Behavior of Murgantia histrionica (Heteroptera: Pentatomidae). Ann. Entomol. Soc. Am. 2008, 101, 215–228. [Google Scholar] [CrossRef]
- Silva, F.A.C.; Da Silva, J.J.; Depieri, R.A.; Panizzi, A.R. Feeding Activity, Salivary Amylase Activity, and Superficial Damage to Soybean Seed by Adult Edessa meditabunda (F.) and Euschistus heros (F.) (Hemiptera: Pentatomidae). Neotrop. Entomol. 2012, 41, 386–390. [Google Scholar] [CrossRef]
- Singh, H.; Malik, V.S. Biology of Painted Bug (Bagrada cruciferarum). Indian J. Agric. Sci. 1993, 63, 672. [Google Scholar]
- Baccaro, S.; Cemmi, A.; Di Sarcina, I.; Ferrara, G. Gamma Irradiation Calliope Facility at ENEA—Casaccia Research Centre (Rome, Italy); ENEA: Rome, Italy, 2019; ENEA Technical Report RT/2019/4/ENEA; ISSN 0393-3016. [Google Scholar]
- Guarino, S.; De Pasquale, C.; Peri, E.; Alonzo, G.; Colazza, S. Role of Volatile and Contact Pheromones in the Mating Behaviour of Bagrada hilaris (Heteroptera: Pentatomidae). Eur. J. Entomol. 2008, 105, 613–617. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Soft. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- SPSS Inc. PASW Statistics 17.0 for Windows, version 17.0; SPSS Inc.: Chicago, IL, USA, 2009. [Google Scholar]
- Hardin, J.W.; Hilbe, J.M. Generalized Estimating Equations; Chapman and Hall/CRC: Boca Raton, FL, USA, 2002. [Google Scholar]
- Halekoh, U.; Højsgaard, S.; Yan, J. The R Package Geepack for Generalized Estimating Equations. J. Stat. Soft. 2006, 15, 1–11. [Google Scholar] [CrossRef]
- Grilli, L.; Rampichini, C. Specification of Random Effects in Multilevel Models: A Review. Qual. Quant. 2015, 49, 967–976. [Google Scholar] [CrossRef]
- Pan, W. Akaike’s Information Criterion in Generalized Estimating Equations. Biometrics 2001, 57, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Hastie, T.J.; Pregibon, D. Generalized Linear Models. In Statistical Models In S; Chambers, J.M., Hastie, T.J., Eds.; Wadsworth & Brooks/Cole: Pacific Grove, CA, USA, 1992; Chapter 6. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Soft. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Peccerillo, C.; Mainardi, C.E.; Nieri, R.; Fouani, J.M.; Cemmi, A.; Cristofaro, M.; Anfora, G.; Mazzoni, V. The Effect of the Sterile Insect Technique on Vibrational Communication: The Case of Bagrada hilaris (Hemiptera: Pentatomidae). Insects 2023, 14, 353. [Google Scholar] [CrossRef] [PubMed]
- McLain, D.K. Female Choice and the Adaptive Significance of Prolonged Copulation in Nezara viridula (Hemiptera: Pentatomidae). Psyche A J. Entomol. 1980, 87, 325–336. [Google Scholar] [CrossRef]
- Alcock, J. Postinsemination Associations Between Males and Females in Insects: The Mate-Guarding Hypothesis. Annu. Rev. Entomol. 1994, 39, 1–21. [Google Scholar] [CrossRef]
- Van Gerwen, A.C.M.; Browne, L.B.; Hoo, C.F.S. Factors Affecting the Time of the Crepuscular Mating Flight and Duration of Mating of the Moth Oncopera alboguttata Tindale (Lepidoptera: Hepialidae). Aust. J. Entomol. 1972, 11, 340–343. [Google Scholar] [CrossRef]
- Mainardi, C.E.; Peccerillo, C.; Paolini, A.; Cemmi, A.; Sforza, R.F.H.; Musmeci, S.; Porretta, D.; Cristofaro, M. Using Gamma Irradiation to Predict Sperm Competition Mechanism in Bagrada hilaris (Burmeister) (Hemiptera: Pentatomidae): Insights for a Future Management Strategy. Insects 2023, 14, 681. [Google Scholar] [CrossRef]
- Danielsson, I. Mechanisms of Sperm Competition in Insects. Ann. Zool. Fenn. 1998, 35, 241–257. [Google Scholar]
- Parker, G.A. Sperm Competition Games: Raffles and Roles. Proc. R. Soc. Lond. B 1990, 242, 120–126. [Google Scholar] [CrossRef]
- Woodhead, A.P. Sperm Mixing in the Cockroach Diploptera punctata. Evolution 1985, 39, 159–164. [Google Scholar] [CrossRef]
- Huang, T.-I.; Reed, D.A.; Perring, T.M.; Palumbo, J.C. Diel Activity and Behavior of Bagrada hilaris (Hemiptera: Pentatomidae) on Desert Cole Crops. J. Econ. Entom. 2013, 106, 1726–1738. [Google Scholar] [CrossRef] [PubMed]
- Franz, G.; Bourtzis, K.; Cáceres, C. Practical and Operational Genetic Sexing Systems Based on Classical Genetic Approaches in Fruit Flies, an Example for Other Species Amenable to Large-Scale Rearing for the Sterile Insect Technique. In Sterile Insect Technique; CRC Press: Boca Raton, FL, USA, 2021; pp. 575–604. [Google Scholar]
- Kumano, N.; Haraguchi, D.; Kohama, T. Effect of Irradiation on Mating Ability in the Male Sweetpotato Weevil (Coleoptera: Curculionidae). J. Econ. Entomol. 2008, 101, 1198–1203. [Google Scholar]
- Hibino, Y.; Iwahashi, O. Appearance of Wild Females Unreceptive to Sterilized Males on Okinawa Is. in the Eradication Program of the Melon Fly, Dacus cucurbitae Coquillett (Diptera:Tephritidae). Appl. Entomol. Zool. 1991, 26, 265–270. [Google Scholar] [CrossRef]
- Lux, S.A.; Vilardi, J.C.; Liedo, P.; Gaggl, K.; Calcagno, G.E.; Munyiri, F.N.; Vera, M.T.; Manso, F. Effects of Irradiation on the Courtship Behavior of Medfly (Diptera, Tephritidae) Mass Reared for the Sterile Insect Technique. Fla. Entomol. 2002, 85, 102–112. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Baldwin, L.A. The Frequency of U-Shaped Dose Responses in the Toxicological Literature. Toxicol. Sci. 2001, 62, 330–338. [Google Scholar]
- Cutler, G.C.; Amichot, M.; Benelli, G.; Guedes, R.N.C.; Qu, Y.; Rix, R.R.; Ullah, F.; Desneux, N. Hormesis and Insects: Effects and Interactions in Agroecosystems. Sci. Total Environ. 2022, 825, 153899. [Google Scholar] [CrossRef] [PubMed]
- Allinghi, A.; Gramajo, C.; Willink, E.; Vilardi, J. Induction of Sterility in Anastrepha fraterculus (Diptera: Tephritidae) by Gamma Radiation. Fla. Entomol. 2007, 90, 96–102. [Google Scholar]
- Krüger, A.P.; Schlesener, D.C.H.; Martins, L.N.; Wollmann, J.; Deprá, M.; Garcia, F.R.M. Radiation Effects on Drosophila suzukii (Diptera: Drosophilidae) Reproductive Behaviour. J. Appl. Entomol. 2019, 143, 88–94. [Google Scholar] [CrossRef]
- Lanouette, G.; Brodeur, J.; Fournier, F.; Martel, V.; Firlej, A. Effect of Irradiation on the Mating Capacity and Competitiveness of Drosophila suzukii (Diptera: Drosophilidae) for the Development of the Sterile Insect Technique. Can. Entomol. 2020, 152, 563–574. [Google Scholar] [CrossRef]
- Opiyo, E. Survival and Reproductive Potential of Gamma Irradiated Male Glossina pallidipes Austen. Entomol. Exp. Appl. 2001, 99, 397–400. [Google Scholar] [CrossRef]
- Mutika, G.N.; Opiyo, E.; Robinson, A.S. Effect of Low Temperature Treatment on the Quality of Male Adult Glossina pallidipes (Diptera: Glossinidae) in Relation to the Sterile Insect Technique. Entomol. Sci. 2002, 5, 209–214. [Google Scholar]
- Stringer, L.D.; Sullivan, N.J.; Sullivan, T.E.S.; Mitchell, V.J.; Manning, L.-A.M.; Mas, F.; Hood-Nowotny, R.C.; Suckling, D.M. Attractiveness and Competitiveness of Irradiated Light Brown Apple Moths. Entomol. Exp. Appl. 2013, 148, 203–212. [Google Scholar] [CrossRef]
- McGovern, W.L.; McKibben, G.H.; Gueldner, R.C.; Cross, W.H. Irradiated Boll Weevils: Pheromone Production Determined by GLC Analysis12. J. Econ. Entomol. 1975, 68, 521–523. [Google Scholar] [CrossRef]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. (Eds.) Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Springer: Dordrecht, The Netherlands, 2005; ISBN 978-1-4020-4050-4. [Google Scholar]


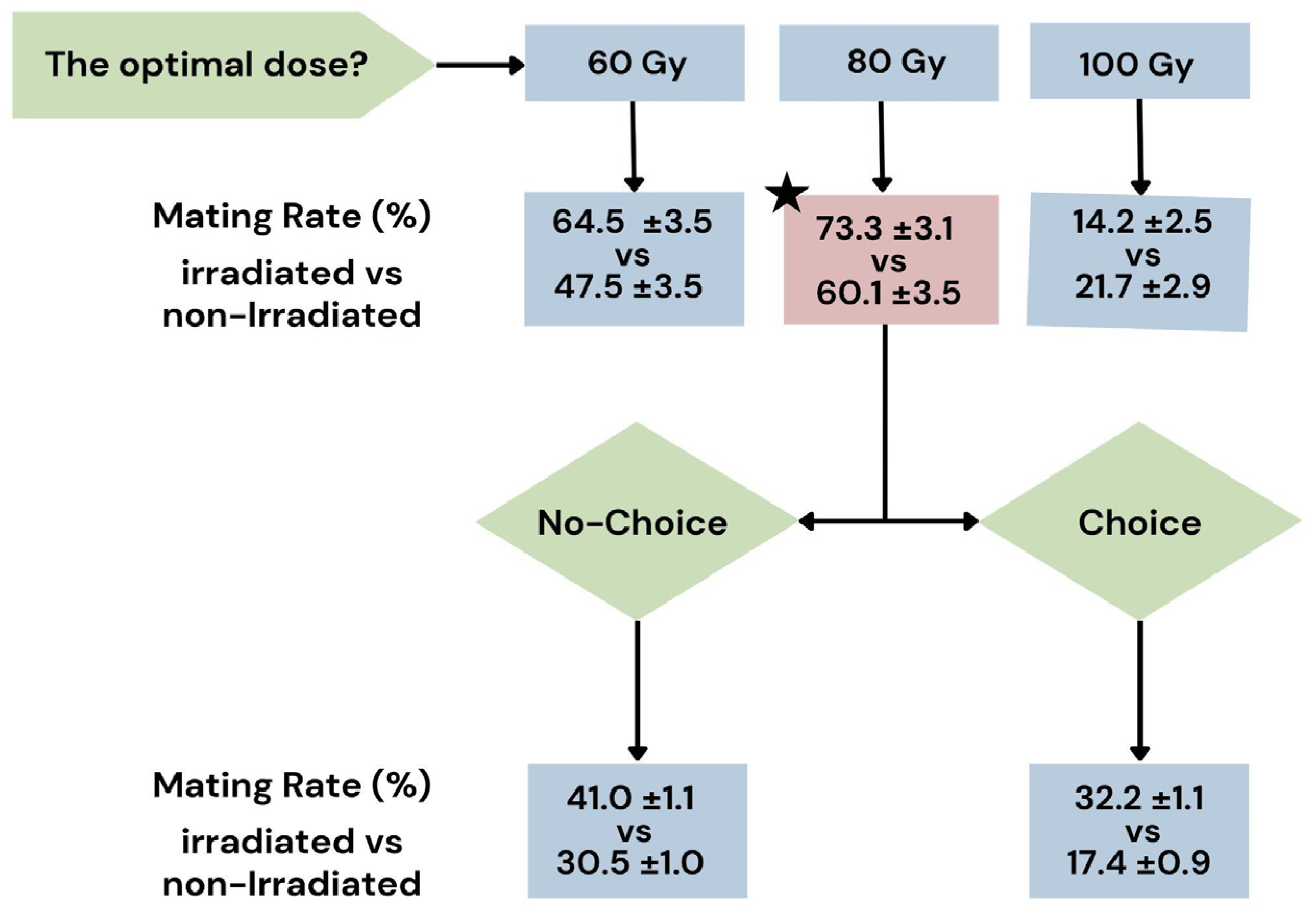
| Experimental Block | Dose (Gy) | Mating Rate (%) | Time Lapse Before First Mating (obs. n.) | First Mating Duration (obs. n.) | N. of Mating Events (n.) |
|---|---|---|---|---|---|
| 1 | 0 | 47.5 ± 3.5 | 9.33 ± 4.09 | 46.20 ± 6.24 | 1.25 ± 0.22 |
| 60 | 64.5 ± 3.5 | 7.06 ± 3.77 | 49.94 ± 4.78 | 1.40 ± 0.24 | |
| 2 | 0 | 60.1 ± 3.5 | 4.67 ± 2.21 | 43.67 ± 7.65 | 1.10 ± 0.19 |
| 80 | 73.3 ± 3.1 | 1.31 * ± 0.55 | 57.56 ± 5.64 | 1.16 ± 0.21 | |
| 3 | 0 | 21.7 ± 2.9 | 16.64 ± 4.40 | 28.55 ± 7.78 | 0.55 ± 0.11 |
| 100 | 14.2 * ± 2.5 | 26.00 ± 10.59 | 34.33 ± 8.80 | 0.30 ± 0.11 |
| Male Treatment | Mating | Female Feeding | Male Feeding | |||||
|---|---|---|---|---|---|---|---|---|
| Days | N Cages | % of Time | ±s.e | % of Time | ±s.e. | % of Time | ±s.e. | |
| 1 | irradiated | 95 | 38.5 | ±1.9 ** | 36.7 | ±1.9 *** | 22.3 | ±1.6 |
| non-irradiated | 99 | 21.5 | ±1.5 | 34.9 | ±1.8 *** | 21.1 | ±1.6 | |
| 2 | irradiated | 95 | 47.5 | ±1.9 ** | 51.1 | ±1.9 *** | 38.0 | ±1.9 |
| non-irradiated | 99 | 31.1 | ±1.8 | 58.2 | ±1.9 *** | 38.2 | ±1.8 | |
| 3 | irradiated | 95 | 37.0 | ±1.9 | 48.7 | ±1.9 ** | 39.1 | ±1.9 |
| non-irradiated | 99 | 39.2 | ±1.9 | 53.5 | ±1.9 ** | 45.3 | ±1.9 | |
| Total mean | irradiated | 95 | 41.0 | ±1.1 ** | 45.5 | ±1.1 *** | 33.1 | ±1.1 |
| non-irradiated | 99 | 30.5 | ±1.0 | 49.9 | ±1.1 *** | 34.9 | ±1.0 |
| Fixed Effects | Estimate | ±s.e. | Wald | p-Value |
|---|---|---|---|---|
| (Intercept) | −0.4416 | 0.1698 | 6.76 | 0.00930 ** |
| Irradiated | 0.8662 | 0.2722 | 10.12 | 0.00146 ** |
| Day 2 | 0.3526 | 0.1558 | 5.12 | 0.02361 * |
| Day 3 | −0.0674 | 0.1734 | 0.15 | 0.69775 |
| Irradiated × day 2 | −0.1559 | 0.2115 | 0.54 | 0.46095 |
| Irradiated × day 3 | −0.9271 | 0.2763 | 11.26 | 0.00079 *** |
| Analysis of “Wald statistic” Table | ||||
| df | X2 | p-value | ||
| Treatment | 1 | 4.88 | 0.02714 * | |
| Days | 2 | 15.95 | 0.00034 *** | |
| Treatment × days | 2 | 14.77 | 0.00062 *** | |
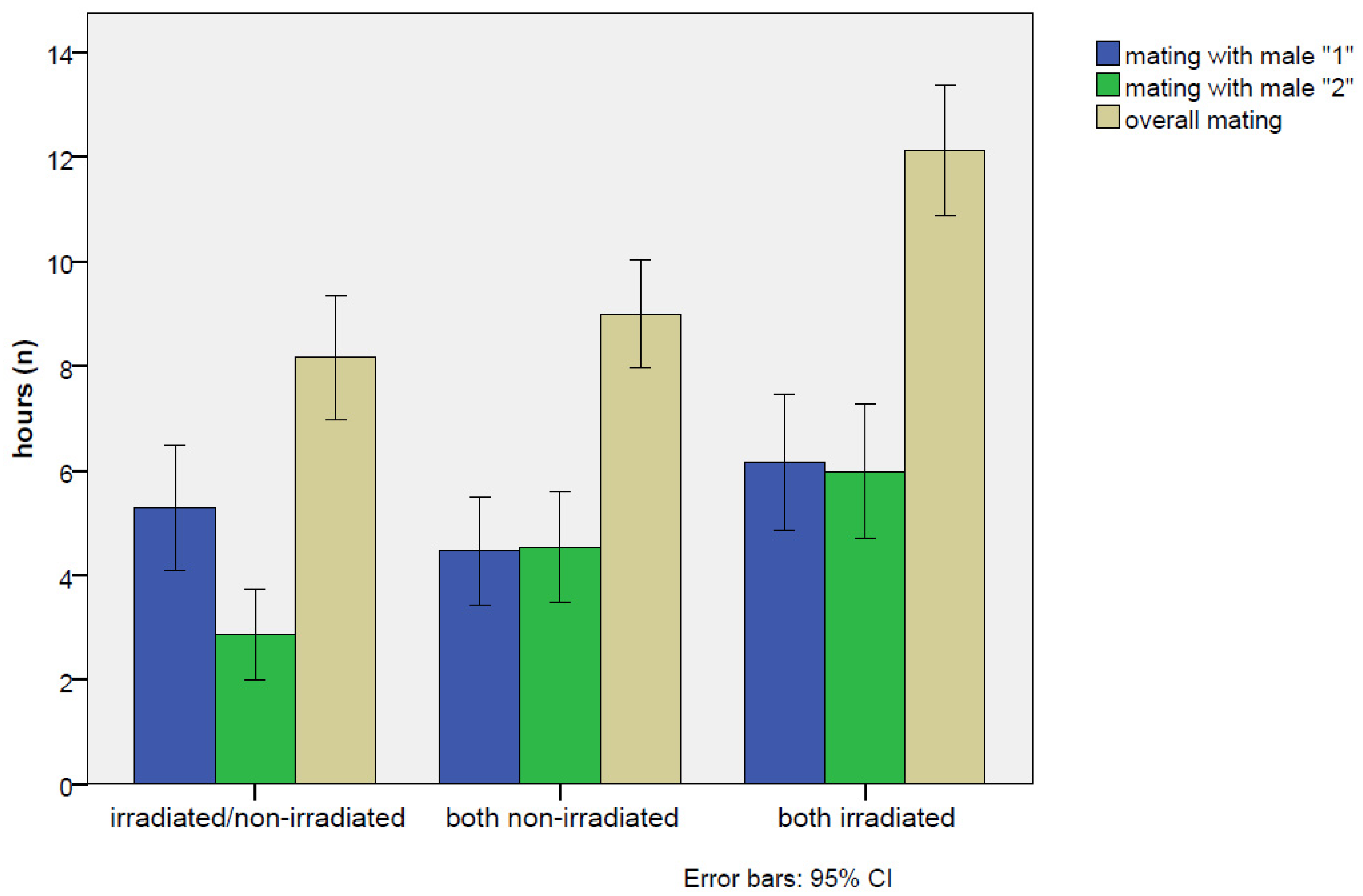
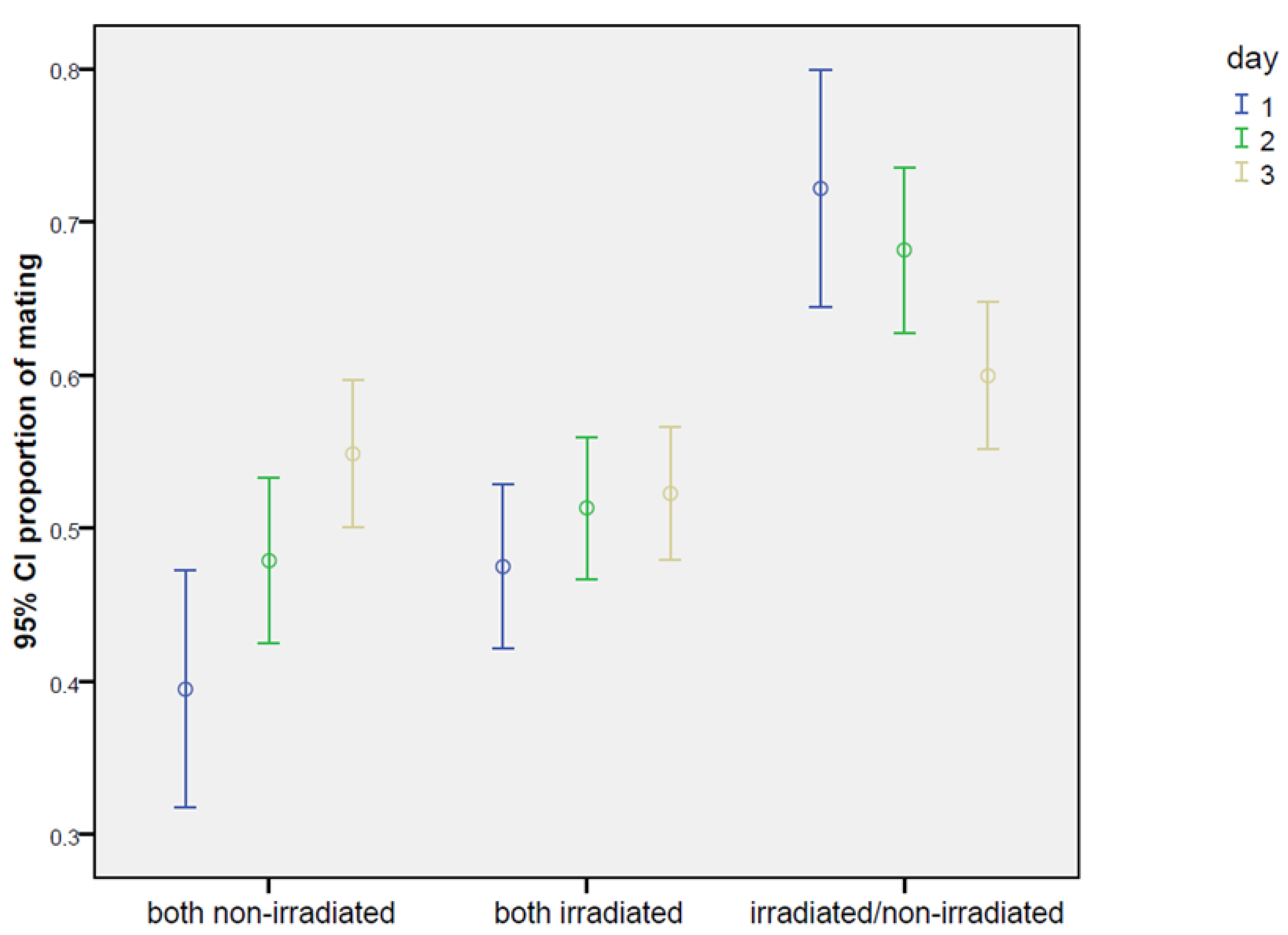
| Male Treatment | Day | Cages (n) | Total Rate of Mating (%) | ±s.e. | Rate of Mating (Male 1 *) (%) | ±s.e. | Rate of Mating (Male 2 *) (%) | ±s.e. |
|---|---|---|---|---|---|---|---|---|
| Irradiated/ non-irradiated * male 1 = irradiated * male 2 = non-irradiated | 1 | 23.7 a | 1.8 | 17.4 a | 1.6 ** | 6.3 a | 1.0 | |
| 2 | 79 | 52.3 b | 2.1 | 35.6 b | 2.0 ** | 16.6 b | 1.6 | |
| 3 | 72.7 c | 1.9 | 43.6 b | 2.1 | 29.1 c | 1.9 | ||
| Total mean | 49.6 A | 1.2 | 32.2 A | 1.1 * | 17.4 A | 0.9 | ||
| Both non-irradiated | 1 | 26.5 a | 1.8 | 10.4 a | 1.3 | 16.0 a | 1.5 | |
| 2 | 85 | 55.7 b | 2.0 | 26.7 b | 1.8 | 29.1 b | 1.9 | |
| 3 | 70.5 c | 1.9 | 38.7 c | 2.0 | 31.8 b | 1.9 | ||
| Total mean | 50.8 A | 1.2 | 25.2 A | 1.0 | 25.6 B | 1.0 | ||
| Both irradiated | 1 | 52.0 a | 2.0 | 25.3 a | 1.7 | 27.8 a | 1.8 | |
| 2 | 91 | 70.8 b | 1.8 | 36.4 b | 1.9 | 34.5 ab | 1.9 | |
| 3 | 80.0 b | 1.6 | 41.8 b | 2.0 | 38.1 b | 1.9 | ||
| Total mean | 67.9 B | 1.1 | 34.4 A | 1.1 | 33.5 B | 1.1 |
| Male Treatment | Fixed Effects | Estimate | ±s.e. | Wald | p-Value |
|---|---|---|---|---|---|
| day 1 | 0.2979 | 0.1096 | 7.40 | 0.0065 ** | |
| Irradiated/non-irradiated | day 2 | 0.2434 | 0.0906 | 7.23 | 0.0072 ** |
| day 3 | 0.1329 | 0.0826 | 2.59 | 0.1077 | |
| Test ANOVA | df | X2 | p-value | ||
| 3 | 9.17 | 0.027 | |||
| Fixed effects | estimate | ±s.e. | Wald | p-value | |
| Both non-irradiated | day 1 | −0.0283 | 0.0891 | 0.10 | 0.75 |
| day 2 | −0.1404 | 0.1207 | 1.35 | 0.24 | |
| day 3 | 0.0647 | 0.0825 | 0.62 | 0.43 | |
| Test ANOVA | df | X2 | p-value | ||
| 3 | 4.23 | 0.24 | |||
| Fixed effects | estimate | ±s.e. | Wald | p-value | |
| Both irradiated | day 1 | −0.0423 | 0.0911 | 0.22 | 0.64 |
| day 2 | 0.0178 | 0.0802 | 0.05 | 0.82 | |
| day 3 | 0.0339 | 0.0747 | 0.21 | 0.65 | |
| Test ANOVA | df | X2 | p-value | ||
| 3 | 1.28 | 0.73 |
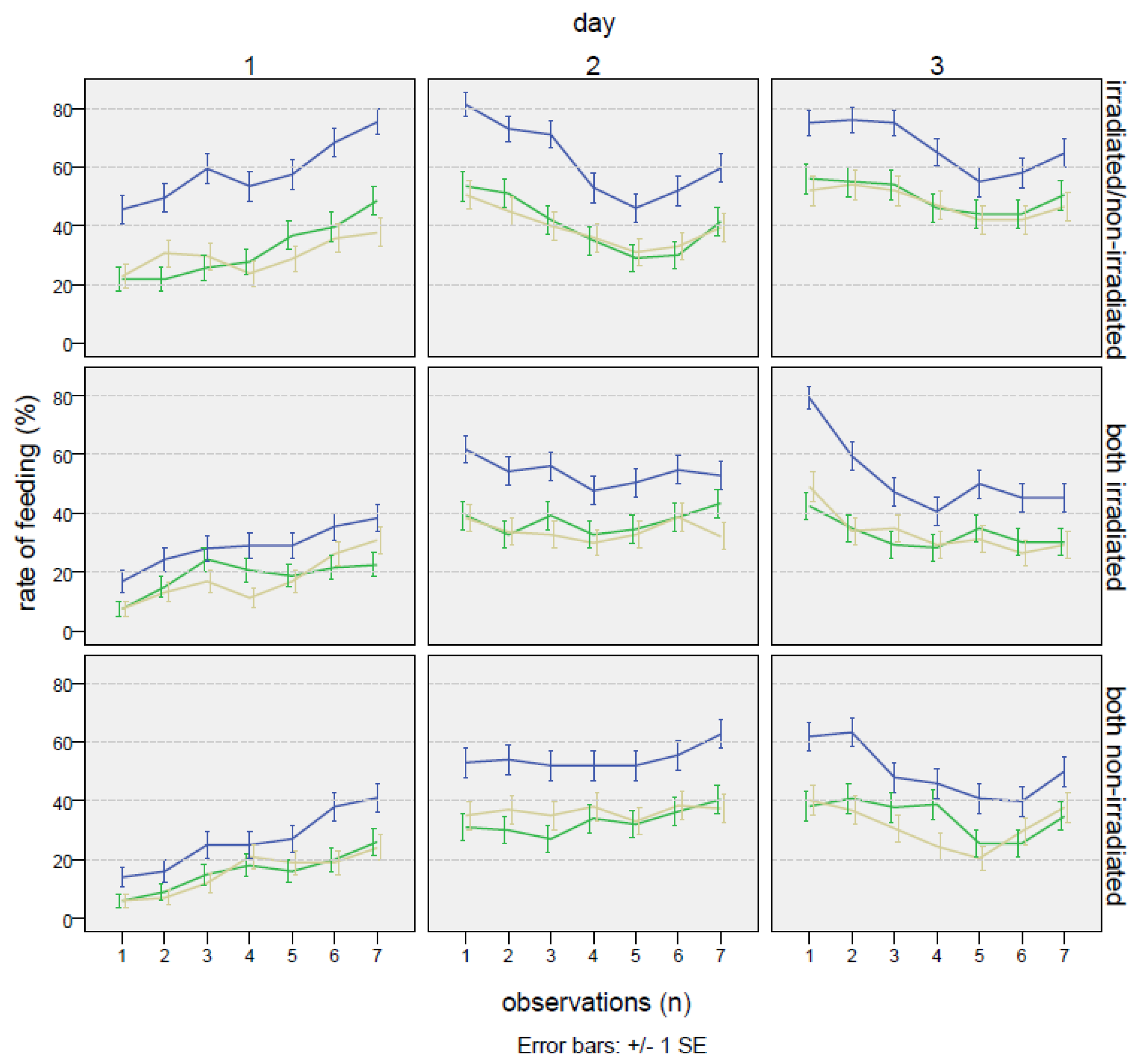
| Male Treatment | Day | Cages (n) | Feeding Rate of Female (%) | ±s.e. | Feeding Rate of Male 1 * (%) | ±s.e. | Feeding Rate of Male 2 * (%) | ±s.e. |
|---|---|---|---|---|---|---|---|---|
| Irradiated/ non-irradiated * male 1 = irradiated * male 2 = non-irradiated | 1 | 57.7 a | 2.1 | 34.4 a | 2.0 | 30.6 a | 2.0 | |
| 2 | 79 | 61.5 a | 2.1 | 43.6 ab | 2.1 | 41.2 ab | 2.1 | |
| 3 | 65.6 a | 2.0 | 51.4 b | 2.1 | 51.9 b | 2.1 | ||
| Total mean | 61.6 A | 1.2 | 43.1 A | 1.2 | 41.2 A | 1.2 | ||
| Both non-irradiated | 1 | 28.5 a | 1.9 | 15.9 a | 1.5 | 16.6 a | 1.5 | |
| 2 | 85 | 58.8 b | 2.0 | 34.8 b | 2.0 | 40.3 b | 2.0 | |
| 3 | 53.8 b | 2.1 | 38.0 b | 2.0 | 34.1 b | 2.0 | ||
| Total mean | 47.1 B | 1.2 | 29.5 B | 1.1 | 30.3 B | 1.1 | ||
| Both irradiated | 1 | 30.1 a | 1.8 | 15.9 a | 1.4 | 19.5 a | 1.6 | |
| 2 | 91 | 54.5 b | 2.0 | 38.8 b | 1.9 | 37.4 b | 1.9 | |
| 3 | 51.6 b | 2.0 | 33.3 b | 1.9 | 34.7 b | 1.9 | ||
| Total mean | 45.4 B | 1.1 | 29.3 B | 1.0 | 30.5 B | 1.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mainardi, C.E.; Peccerillo, C.; Musmeci, S.; Paolini, A.; Sforza, R.F.H.; Cemmi, A.; Di Sarcina, I.; Anfora, G.; Porretta, D.; Marini, F.; et al. Sterile but Sexy: Assessing the Mating Competitiveness of Irradiated Bagrada hilaris Males for the Development of a Sterile Insect Technique. Insects 2025, 16, 391. https://doi.org/10.3390/insects16040391
Mainardi CE, Peccerillo C, Musmeci S, Paolini A, Sforza RFH, Cemmi A, Di Sarcina I, Anfora G, Porretta D, Marini F, et al. Sterile but Sexy: Assessing the Mating Competitiveness of Irradiated Bagrada hilaris Males for the Development of a Sterile Insect Technique. Insects. 2025; 16(4):391. https://doi.org/10.3390/insects16040391
Chicago/Turabian StyleMainardi, Chiara Elvira, Chiara Peccerillo, Sergio Musmeci, Alessandra Paolini, René F. H. Sforza, Alessia Cemmi, Ilaria Di Sarcina, Gianfranco Anfora, Daniele Porretta, Francesca Marini, and et al. 2025. "Sterile but Sexy: Assessing the Mating Competitiveness of Irradiated Bagrada hilaris Males for the Development of a Sterile Insect Technique" Insects 16, no. 4: 391. https://doi.org/10.3390/insects16040391
APA StyleMainardi, C. E., Peccerillo, C., Musmeci, S., Paolini, A., Sforza, R. F. H., Cemmi, A., Di Sarcina, I., Anfora, G., Porretta, D., Marini, F., & Cristofaro, M. (2025). Sterile but Sexy: Assessing the Mating Competitiveness of Irradiated Bagrada hilaris Males for the Development of a Sterile Insect Technique. Insects, 16(4), 391. https://doi.org/10.3390/insects16040391